脊椎動物の神経堤細胞と頭部プラコードは進化的に共通の起源をもつ
堀江良子・堀江健生
(米国Princeton大学Lewis-Sigler Institute for Integrative Genomics)
email:堀江健生
DOI: 10.7875/first.author.2018.073
Shared evolutionary origin of vertebrate neural crest and cranial placodes.
Ryoko Horie, Alex Hazbun, Kai Chen, Chen Cao, Michael Levine, Takeo Horie
Nature, 560, 228-232 (2018)
プラコードおよび神経堤細胞は脊椎動物を特徴づける組織であるが,脊椎動物の進化の過程においてこれらがどのように獲得されたのかは不明であった.この研究において,筆者らは,原始的な脊索動物であるホヤの神経板の境界領域について,細胞系譜の追跡,遺伝子の機能の阻害,1細胞トランスクリプトーム解析などの手法を組み合わせ包括的に解析した.その結果,ホヤの神経板の境界領域はFoxc遺伝子,Six1/2遺伝子,Msxb遺伝子により区画化されており,この区画化のパターンはホヤと脊椎動物とのあいだで保存されていることが明らかにされた.また,おのおのの遺伝子により区画化された領域から,頭部プラコードや神経堤細胞の起源的な性質を備えた感覚ニューロンがそれぞれ生じることが明らかにされた.神経板の境界領域においてこれらの遺伝子制御ネットワークを乱すと,おのおのの感覚ニューロンは互いに運命を変換した.とくに,神経板の境界領域の後方から発生し神経堤細胞との類似性が示されているBTNに着目したところ,BTNは神経板の境界領域のもっとも前方に位置するPSCに運命が変換されることが明らかにされた.また,BTNのPSCへの運命の変換は1細胞トランスクリプトーム解析により確かめられた.これらの結果から,神経板の境界領域の区画化は脊椎動物よりまえに起こっていること,頭蓋プラコードと神経堤細胞は共通の進化的な起源をもつことが示唆された.
脊椎動物の大きな特徴のひとつは,発達した脳,眼,鼻,耳などの感覚器を備えた頭部の構造である.脊椎動物の頭部を構成する脳や脊髄などの中枢神経系は神経板から形成され,頭部の感覚器や感覚器を構成する感覚ニューロンは頭部プラコードおよび神経堤細胞から形成される1-4).発生の過程において,神経板と表皮の前方の境界領域には頭部プラコードが,神経板と表皮の側方の境界領域には神経堤細胞が形成される.頭部プラコードおよび神経堤細胞は脊椎動物に特有の構造であると考えられていたが,最近,脊椎動物にもっとも近縁な無脊椎動物であるホヤにおいて,頭部プラコードおよび神経提細胞の起源的な性質を備えた細胞の存在が報告された5-7).しかしながら,脊椎動物の進化の過程において,頭部プラコードや神経堤細胞がどのように獲得されたのか,たとえば,頭部プラコードと神経堤細胞は進化的に共通の起源をもつのか,それとも,別々の起源をもつのかは不明であった3).この研究において,筆者らは,ホヤを用いて頭部プラコードおよび神経提細胞の発生プログラムについて詳細に解析することにより,脊椎動物の頭部プラコードおよび神経堤細胞の進化的な起源について考察した.
ホヤには神経板の境界領域から生じる4種類の感覚ニューロンが存在し,前方から,PSC(palp sensory cell),aATEN(anterior apical trunk epidermal neuron),pATEN(posterior apical trunk epidermal neuron),BTN(bipolar tail neuron)と名づけられている5-8).このうち,PSCおよびaATENは頭部プラコードの原始的な性質をもつことが報告されている5,6).とくに,aATENは鼻プラコードに由来する細胞にみられるような化学受容性の細胞としての性質と,下垂体プラコードに由来する細胞にみられるような性腺刺激ホルモン放出ホルモンを放出する神経内分泌性の細胞としての性質の両方を備えることが報告されている5).もっとも後方に位置するBTNは神経堤細胞に由来する後根神経節と性質が似ていることが報告されており,神経堤細胞との相似性が指摘されている7).
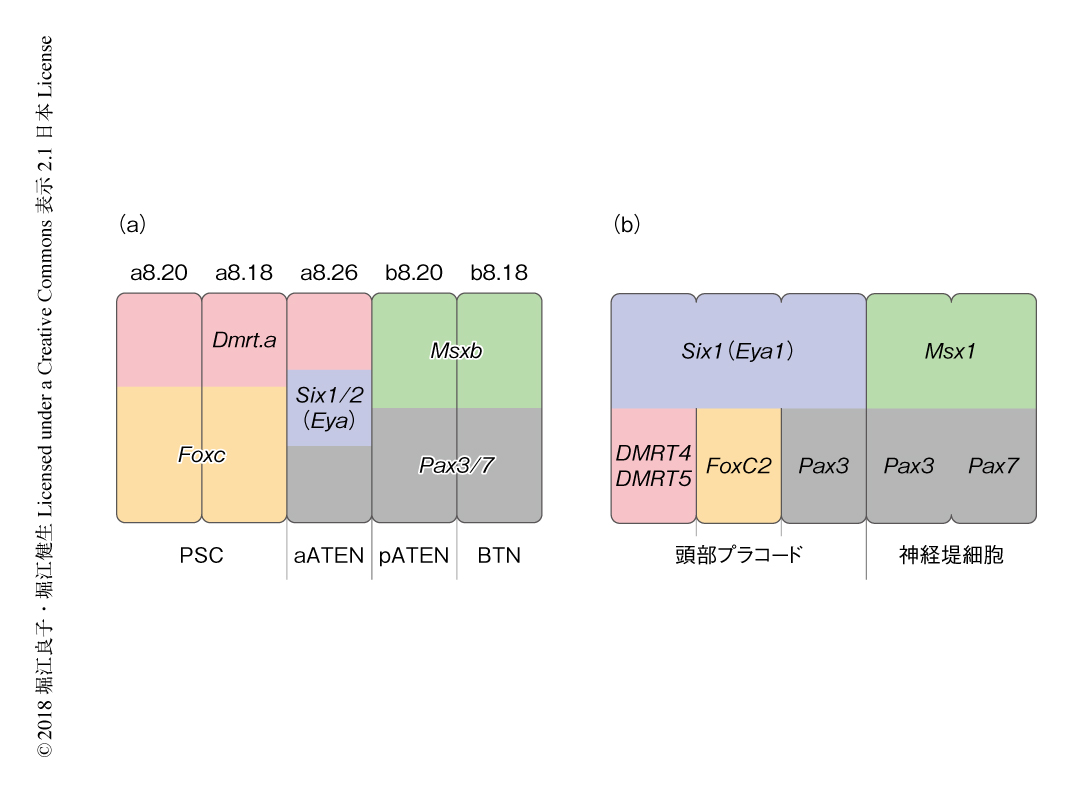
DiIを用いた細胞系譜の追跡およびレポーター遺伝子の発現を詳細に観察することにより,神経板の境界領域から生じるおのおのの感覚ニューロンの細胞系譜が明らかにされた.ホヤの110細胞期において,PSCはa8.20割球およびa8.18割球から,aATENはa8.26割球から,pATENはb8.20割球から,BTNはb8.18割球から,発生した.発生の過程において,おのおのの感覚ニューロンはそれぞれ異なる割球に由来することが明らかにされた.以前のホヤの初期胚の遺伝子制御ネットワークの詳細な解析から,Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子,Pax3/7遺伝子,Msxb遺伝子などの転写因子をコードする遺伝子は神経板の境界領域において発現することが明らかにされている9).Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子は神経板の境界領域の前方のプラコード領域において発現する一方,Msxb遺伝子は神経板の境界領域の後方において発現し,Pax3/7遺伝子は神経板の境界領域の前方と後方にまたがり発現する.Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子,Pax3/7遺伝子,Msxb遺伝子による神経板の境界領域の区画化のパターンをホヤと脊椎動物とで比較したところ,非常によく保存されていた(図1).また,これらの転写因子をコードする遺伝子の前後軸にそった発現のパターンは,頭部プラコードに由来する感覚ニューロン,および,神経堤細胞に由来する感覚ニューロンが生じる位置と対応した.
神経板の境界領域におけるおのおのの転写因子をコードする遺伝子の発現パターンが,互いの発現パターンに影響するのかどうか調べた.Dmrt.a遺伝子の機能を阻害した胚においてFoxc遺伝子,Six1/2遺伝子,Eya遺伝子の発現が消失したことから,Dmrt.a遺伝子はFoxc遺伝子およびSix1/2遺伝子の発現を活性化することが明らかにされた.Msxb遺伝子の機能を阻害した胚においてはSix1/2遺伝子やEya遺伝子といったプラコード遺伝子の発現が後方に広がった.つまり,Msxb遺伝子はプラコード遺伝子の発現を抑制することにより,プラコードが神経板の境界領域の後方に形成されないようにすることが示唆された.また,この結果は,Six1/2遺伝子のプラコード領域の最小エンハンサーに多数のMsxb結合配列が存在することと一致した.また,Dmrt.a遺伝子の機能を阻害した胚においてMsxb遺伝子の発現が前方にシフトしたことから,Dmrt.a遺伝子とMsxb遺伝子は互いに発現を抑制しあうことにより神経板の境界領域の前方と後方の境界を確立する可能性が示唆された.
ホヤと脊椎動物の神経板の境界領域の運命図においてもっとも異なるのは,ホヤにおいては神経板の境界領域の前方は異なる2つの領域に分けられることである.つまり,PSCが生じる領域にはFoxc遺伝子が,aATENが生じる領域にはSix1/2遺伝子が発現する.では,プラコードに由来する感覚ニューロンであるPSCおよびaATENは共通の発生プログラムを利用するのだろうか? Foxc遺伝子の機能を阻害したところ,PSCは消失し,本来はPSCが生じる領域においてSix1/2遺伝子の異所的な発現がみられ,そこにaATENが異所的に生じた.この結果から,Foxc遺伝子はPSCにおいてはたらく遺伝子を活性化するとともに,aATENにおいてはたらく遺伝子の発現を抑制することにより,PSCとしてのアイデンティティを確立することが示唆された.さらに,PSCとaATENは互いに運命の変換が可能であったことから,神経板の境界領域の前方は互いに似た発生プログラムを利用する可能性が示唆された.
BTNは脊椎動物において神経堤細胞に由来する感覚ニューロンである後根神経節との関連性が指摘されている7).このことから,ホヤの神経板の境界領域の後方は神経堤細胞の起源的な性質を備えることが示唆され,さらに,脊椎動物よりまえの神経板の境界領域は,脊椎動物のプラコードと共通の起源である可能性が示唆された.このことを検証するため,BTNが神経板の境界領域から生じるほかの細胞に運命が変換することが可能かどうか調べた.Pax3/7遺伝子あるいはMsxb遺伝子の発現制御領域を用い,Foxc遺伝子を神経板の境界領域の後方において異所的に発現させたところ,BTNはPSCに運命を変換したが,その際,いくつかの表現型が観察された.すなわち,PSCのマーカー遺伝子のみを発現する個体もあれば,PSCとBTNの両方のマーカー遺伝子を発現する個体もみられた.BTNがPSCに運命を変換したことは,神経板の境界領域の区画化については異なるが,似ている感覚ニューロンの起源になるという主張を裏づけるキーポイントとなる現象であった.
ホヤの中期尾芽胚は細胞数が約1500個と少なく,細胞系譜も詳細に解析されているため,胚をまるごと用いた1細胞トランスクリプトーム解析により細胞の由来を解析するのによいモデルとなる.Foxc遺伝子を神経板の境界領域の後方において異所的に発現させた胚をバラバラにし,マイクロ流体系を用いた1細胞トランスクリプトーム解析により,BTNのPSCへの運命の変換について詳細に解析した.その結果,tSNE(t-distributed stochastic neighbor embedding,t分布型確率的近傍埋め込み)プロットにより,ホヤの中期尾芽胚には脊索,内胚葉,尾部筋肉,内胚葉,表皮,中枢神経系など,約20種類の細胞クラスターが存在することが示された.合計で45個のBTNが同定され,そのうち約半数の21個は運命を変換せずに本来のBTNの発現を維持していた.10個のBTNは完全にPSCへ運命を変換しており,14個はBTNとPSCの両方のマーカー遺伝子を発現するハイブリッドな表現型を示した.この結果は,レポーター遺伝子を用いた実験の結果と一致した.1細胞トランスクリプトーム解析によりBTNがPSCに運命を変換することが強く示され,このことから,神経板の境界領域から生じるおのおのの感覚ニューロンは似た発生プログラムを使用することが示唆された.
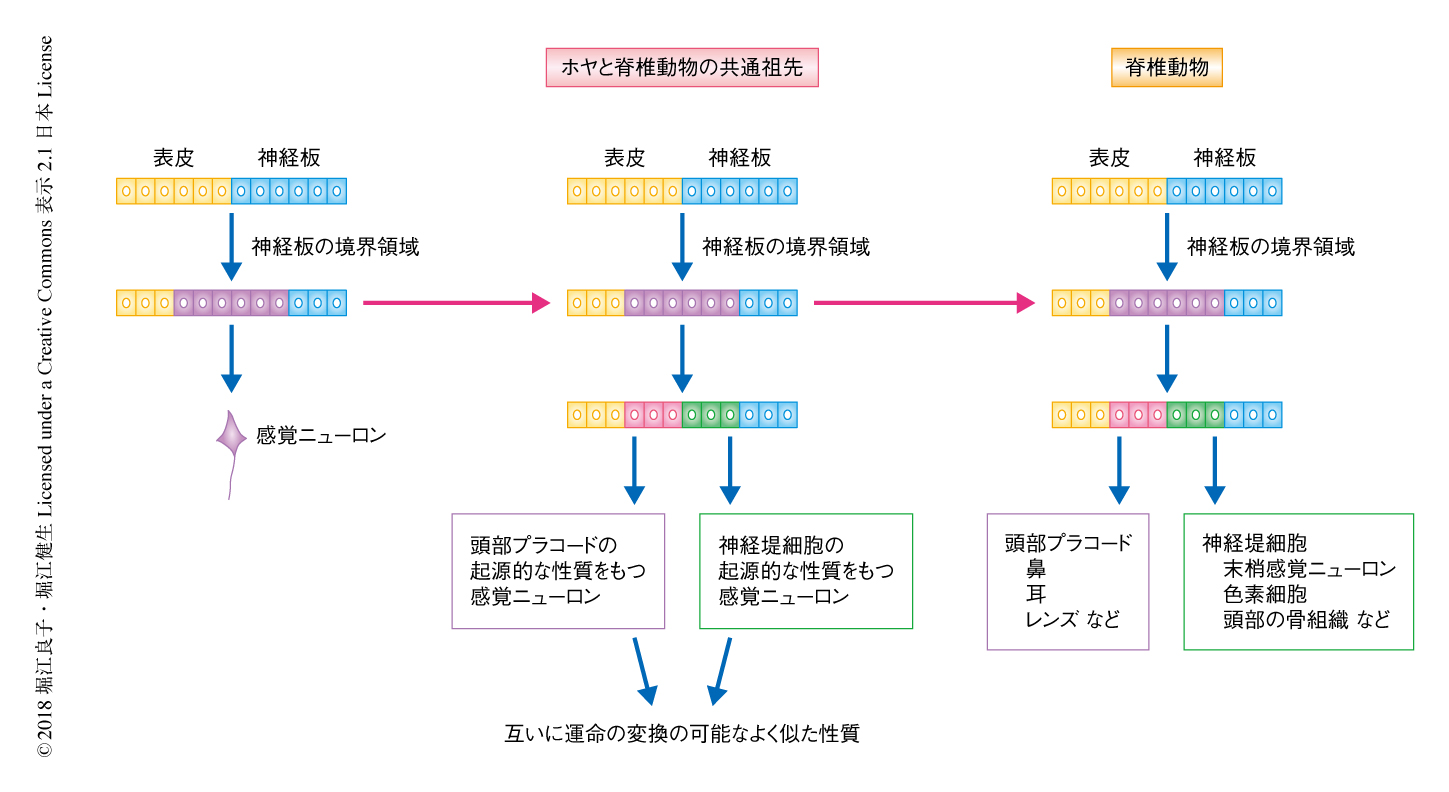
尾索動物のホヤにおいて,神経板の境界領域の前後軸にそった区画化により,互いに似ているが異なる感覚ニューロンが生じることが示された.脊椎動物のプラコードあるいは神経堤細胞から生じる体性感覚ニューロンは互いに顕著な類似性がみられる10).この研究において,筆者らは,神経板の境界領域の区画化は脊椎動物よりまえに存在し,ホヤと脊椎動物の共通祖先の神経板の境界領域は,頭部プラコードおよび神経堤細胞の共通の進化的な起源であるというモデルを提唱する(図2).
略歴:2015年より米国Princeton大学Research Specialist.
研究テーマ:脊椎動物への進化の背景にある遺伝子プログラム.
関心事:家事と仕事の時短.
堀江 健生(Takeo Horie)
筑波大学生命環境系 助教.
研究室URL:http://accafe.jp/Horie_Takeo/index.php
© 2018 堀江良子・堀江健生 Licensed under CC 表示 2.1 日本
(米国Princeton大学Lewis-Sigler Institute for Integrative Genomics)
email:堀江健生
DOI: 10.7875/first.author.2018.073
Shared evolutionary origin of vertebrate neural crest and cranial placodes.
Ryoko Horie, Alex Hazbun, Kai Chen, Chen Cao, Michael Levine, Takeo Horie
Nature, 560, 228-232 (2018)
要 約
プラコードおよび神経堤細胞は脊椎動物を特徴づける組織であるが,脊椎動物の進化の過程においてこれらがどのように獲得されたのかは不明であった.この研究において,筆者らは,原始的な脊索動物であるホヤの神経板の境界領域について,細胞系譜の追跡,遺伝子の機能の阻害,1細胞トランスクリプトーム解析などの手法を組み合わせ包括的に解析した.その結果,ホヤの神経板の境界領域はFoxc遺伝子,Six1/2遺伝子,Msxb遺伝子により区画化されており,この区画化のパターンはホヤと脊椎動物とのあいだで保存されていることが明らかにされた.また,おのおのの遺伝子により区画化された領域から,頭部プラコードや神経堤細胞の起源的な性質を備えた感覚ニューロンがそれぞれ生じることが明らかにされた.神経板の境界領域においてこれらの遺伝子制御ネットワークを乱すと,おのおのの感覚ニューロンは互いに運命を変換した.とくに,神経板の境界領域の後方から発生し神経堤細胞との類似性が示されているBTNに着目したところ,BTNは神経板の境界領域のもっとも前方に位置するPSCに運命が変換されることが明らかにされた.また,BTNのPSCへの運命の変換は1細胞トランスクリプトーム解析により確かめられた.これらの結果から,神経板の境界領域の区画化は脊椎動物よりまえに起こっていること,頭蓋プラコードと神経堤細胞は共通の進化的な起源をもつことが示唆された.
はじめに
脊椎動物の大きな特徴のひとつは,発達した脳,眼,鼻,耳などの感覚器を備えた頭部の構造である.脊椎動物の頭部を構成する脳や脊髄などの中枢神経系は神経板から形成され,頭部の感覚器や感覚器を構成する感覚ニューロンは頭部プラコードおよび神経堤細胞から形成される1-4).発生の過程において,神経板と表皮の前方の境界領域には頭部プラコードが,神経板と表皮の側方の境界領域には神経堤細胞が形成される.頭部プラコードおよび神経堤細胞は脊椎動物に特有の構造であると考えられていたが,最近,脊椎動物にもっとも近縁な無脊椎動物であるホヤにおいて,頭部プラコードおよび神経提細胞の起源的な性質を備えた細胞の存在が報告された5-7).しかしながら,脊椎動物の進化の過程において,頭部プラコードや神経堤細胞がどのように獲得されたのか,たとえば,頭部プラコードと神経堤細胞は進化的に共通の起源をもつのか,それとも,別々の起源をもつのかは不明であった3).この研究において,筆者らは,ホヤを用いて頭部プラコードおよび神経提細胞の発生プログラムについて詳細に解析することにより,脊椎動物の頭部プラコードおよび神経堤細胞の進化的な起源について考察した.
1.神経板の境界領域における遺伝子の区画化のパターンはホヤと脊椎動物とのあいだで保存されている
ホヤには神経板の境界領域から生じる4種類の感覚ニューロンが存在し,前方から,PSC(palp sensory cell),aATEN(anterior apical trunk epidermal neuron),pATEN(posterior apical trunk epidermal neuron),BTN(bipolar tail neuron)と名づけられている5-8).このうち,PSCおよびaATENは頭部プラコードの原始的な性質をもつことが報告されている5,6).とくに,aATENは鼻プラコードに由来する細胞にみられるような化学受容性の細胞としての性質と,下垂体プラコードに由来する細胞にみられるような性腺刺激ホルモン放出ホルモンを放出する神経内分泌性の細胞としての性質の両方を備えることが報告されている5).もっとも後方に位置するBTNは神経堤細胞に由来する後根神経節と性質が似ていることが報告されており,神経堤細胞との相似性が指摘されている7).
DiIを用いた細胞系譜の追跡およびレポーター遺伝子の発現を詳細に観察することにより,神経板の境界領域から生じるおのおのの感覚ニューロンの細胞系譜が明らかにされた.ホヤの110細胞期において,PSCはa8.20割球およびa8.18割球から,aATENはa8.26割球から,pATENはb8.20割球から,BTNはb8.18割球から,発生した.発生の過程において,おのおのの感覚ニューロンはそれぞれ異なる割球に由来することが明らかにされた.以前のホヤの初期胚の遺伝子制御ネットワークの詳細な解析から,Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子,Pax3/7遺伝子,Msxb遺伝子などの転写因子をコードする遺伝子は神経板の境界領域において発現することが明らかにされている9).Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子は神経板の境界領域の前方のプラコード領域において発現する一方,Msxb遺伝子は神経板の境界領域の後方において発現し,Pax3/7遺伝子は神経板の境界領域の前方と後方にまたがり発現する.Dmrt.a遺伝子,Foxc遺伝子,Six1/2遺伝子,Pax3/7遺伝子,Msxb遺伝子による神経板の境界領域の区画化のパターンをホヤと脊椎動物とで比較したところ,非常によく保存されていた(図1).また,これらの転写因子をコードする遺伝子の前後軸にそった発現のパターンは,頭部プラコードに由来する感覚ニューロン,および,神経堤細胞に由来する感覚ニューロンが生じる位置と対応した.
神経板の境界領域におけるおのおのの転写因子をコードする遺伝子の発現パターンが,互いの発現パターンに影響するのかどうか調べた.Dmrt.a遺伝子の機能を阻害した胚においてFoxc遺伝子,Six1/2遺伝子,Eya遺伝子の発現が消失したことから,Dmrt.a遺伝子はFoxc遺伝子およびSix1/2遺伝子の発現を活性化することが明らかにされた.Msxb遺伝子の機能を阻害した胚においてはSix1/2遺伝子やEya遺伝子といったプラコード遺伝子の発現が後方に広がった.つまり,Msxb遺伝子はプラコード遺伝子の発現を抑制することにより,プラコードが神経板の境界領域の後方に形成されないようにすることが示唆された.また,この結果は,Six1/2遺伝子のプラコード領域の最小エンハンサーに多数のMsxb結合配列が存在することと一致した.また,Dmrt.a遺伝子の機能を阻害した胚においてMsxb遺伝子の発現が前方にシフトしたことから,Dmrt.a遺伝子とMsxb遺伝子は互いに発現を抑制しあうことにより神経板の境界領域の前方と後方の境界を確立する可能性が示唆された.
2.神経板の境界領域の前方の領域は互いに似た発生プログラムを利用する
ホヤと脊椎動物の神経板の境界領域の運命図においてもっとも異なるのは,ホヤにおいては神経板の境界領域の前方は異なる2つの領域に分けられることである.つまり,PSCが生じる領域にはFoxc遺伝子が,aATENが生じる領域にはSix1/2遺伝子が発現する.では,プラコードに由来する感覚ニューロンであるPSCおよびaATENは共通の発生プログラムを利用するのだろうか? Foxc遺伝子の機能を阻害したところ,PSCは消失し,本来はPSCが生じる領域においてSix1/2遺伝子の異所的な発現がみられ,そこにaATENが異所的に生じた.この結果から,Foxc遺伝子はPSCにおいてはたらく遺伝子を活性化するとともに,aATENにおいてはたらく遺伝子の発現を抑制することにより,PSCとしてのアイデンティティを確立することが示唆された.さらに,PSCとaATENは互いに運命の変換が可能であったことから,神経板の境界領域の前方は互いに似た発生プログラムを利用する可能性が示唆された.
3.神経板の境界領域から生じる感覚ニューロンは互いに運命の変換の可能なよく似た性質を備える
BTNは脊椎動物において神経堤細胞に由来する感覚ニューロンである後根神経節との関連性が指摘されている7).このことから,ホヤの神経板の境界領域の後方は神経堤細胞の起源的な性質を備えることが示唆され,さらに,脊椎動物よりまえの神経板の境界領域は,脊椎動物のプラコードと共通の起源である可能性が示唆された.このことを検証するため,BTNが神経板の境界領域から生じるほかの細胞に運命が変換することが可能かどうか調べた.Pax3/7遺伝子あるいはMsxb遺伝子の発現制御領域を用い,Foxc遺伝子を神経板の境界領域の後方において異所的に発現させたところ,BTNはPSCに運命を変換したが,その際,いくつかの表現型が観察された.すなわち,PSCのマーカー遺伝子のみを発現する個体もあれば,PSCとBTNの両方のマーカー遺伝子を発現する個体もみられた.BTNがPSCに運命を変換したことは,神経板の境界領域の区画化については異なるが,似ている感覚ニューロンの起源になるという主張を裏づけるキーポイントとなる現象であった.
ホヤの中期尾芽胚は細胞数が約1500個と少なく,細胞系譜も詳細に解析されているため,胚をまるごと用いた1細胞トランスクリプトーム解析により細胞の由来を解析するのによいモデルとなる.Foxc遺伝子を神経板の境界領域の後方において異所的に発現させた胚をバラバラにし,マイクロ流体系を用いた1細胞トランスクリプトーム解析により,BTNのPSCへの運命の変換について詳細に解析した.その結果,tSNE(t-distributed stochastic neighbor embedding,t分布型確率的近傍埋め込み)プロットにより,ホヤの中期尾芽胚には脊索,内胚葉,尾部筋肉,内胚葉,表皮,中枢神経系など,約20種類の細胞クラスターが存在することが示された.合計で45個のBTNが同定され,そのうち約半数の21個は運命を変換せずに本来のBTNの発現を維持していた.10個のBTNは完全にPSCへ運命を変換しており,14個はBTNとPSCの両方のマーカー遺伝子を発現するハイブリッドな表現型を示した.この結果は,レポーター遺伝子を用いた実験の結果と一致した.1細胞トランスクリプトーム解析によりBTNがPSCに運命を変換することが強く示され,このことから,神経板の境界領域から生じるおのおのの感覚ニューロンは似た発生プログラムを使用することが示唆された.
おわりに
尾索動物のホヤにおいて,神経板の境界領域の前後軸にそった区画化により,互いに似ているが異なる感覚ニューロンが生じることが示された.脊椎動物のプラコードあるいは神経堤細胞から生じる体性感覚ニューロンは互いに顕著な類似性がみられる10).この研究において,筆者らは,神経板の境界領域の区画化は脊椎動物よりまえに存在し,ホヤと脊椎動物の共通祖先の神経板の境界領域は,頭部プラコードおよび神経堤細胞の共通の進化的な起源であるというモデルを提唱する(図2).
文 献
- Northcutt, R. G. & Gans, C.: The genesis of neural crest and epidermal placodes: a reinterpretation of vertebrate origins. Q. Rev. Biol., 58, 1-28 (1983)[PubMed]
- Baker, C. V. & Bronner-Fraser, M.: The origins of the neural crest. Part II: an evolutionary perspective. Mech. Dev., 69, 13-29 (1997)[PubMed]
- Schlosser, G.: Do vertebrate neural crest and cranial placodes have a common evolutionary origin? Bioessays, 30, 659-672 (2008)[PubMed]
- Schlosser, G., Patthey, C. & Shimeld, S. M.: The evolutionary history of vertebrate cranial placodes II. Evolution of ectodermal patterning. Dev. Biol., 389, 98-119 (2014)[PubMed]
- Manni, L., Lane, N. J., Joly, J. S. et al.: Neurogenic and non-neurogenic placodes in ascidians. J. Exp. Zool. B Mol. Dev. Evol., 302, 483-504 (2004)[PubMed]
- Abitua, P. B., Gainous, T. B., Kaczmarczyk, A. N. et al.: The pre-vertebrate origins of neurogenic placodes. Nature, 524, 462-465 (2015)[PubMed]
- Stolfi, A., Ryan, K., Meinertzhagen, I. A. et al.: Migratory neuronal progenitors arise from the neural plate borders in tunicates. Nature, 527, 371-374 (2015)[PubMed]
- Imai, J. H. & Meinerzhagen, I. A.: Neurons of the ascidian larval nervous system in Ciona intestinalis: II. peripheral nervous system. J. Comp. Neurol., 501, 335-352 (2007)[PubMed]
- Imai, K. S., Levine, M., Satoh, N. et al.: Regulatory blueprint for a chordate embryo. Science, 312, 1183-1187 (2006)[PubMed]
- Schlosser, G.: Induction and specification of cranial placode. Dev. Biol., 294, 303-351 (2006)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2015年より米国Princeton大学Research Specialist.
研究テーマ:脊椎動物への進化の背景にある遺伝子プログラム.
関心事:家事と仕事の時短.
堀江 健生(Takeo Horie)
筑波大学生命環境系 助教.
研究室URL:http://accafe.jp/Horie_Takeo/index.php
© 2018 堀江良子・堀江健生 Licensed under CC 表示 2.1 日本
