イネにおいてエチレン-ジベレリンリレーは定期的な洪水への適応に貢献する
黒羽 剛1・芦苅基行2
(1東北大学大学院生命科学研究科 植物細胞壁分野,2名古屋大学生物機能開発利用研究センター 高次生体分子機能研究分野)
email:黒羽 剛,芦苅基行
DOI: 10.7875/first.author.2018.071
Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding.
Takeshi Kuroha, Keisuke Nagai, Rico Gamuyao, Diane R. Wang, Tomoyuki Furuta, Masanari Nakamori, Takuya Kitaoka, Keita Adachi, Anzu Minami, Yoshinao Mori, Kiyoshi Mashiguchi, Yoshiya Seto, Shinjiro Yamaguchi, Mikiko Kojima, Hitoshi Sakakibara, Jianzhong Wu, Kaworu Ebana, Nobutaka Mitsuda, Masaru Ohme-Takagi, Shuichi Yanagisawa, Masanori Yamasaki, Ryusuke Yokoyama, Kazuhiko Nishitani, Toshihiro Mochizuki, Gen Tamiya, Susan R. McCouch, Motoyuki Ashikari
Science, 361, 181-186 (2018)
動物と異なり移動することのできない植物は,過酷な環境に遭遇してもその場所において対応しなければならない.植物はさまざまな環境の変化に対し適応する機能を獲得することによりこのような難題を克服してきた.なかでも,“浮きイネ”とよばれるイネは定期的に発生する洪水に対し草丈を伸ばすというユニークな戦略により生き延びる.この研究において,筆者らは,浮きイネの洪水への適応にかかわる遺伝子として,植物ホルモンであるジベレリンを合成する酵素をコードするSD1遺伝子を同定した.また,洪水により別の植物ホルモンであるエチレンが蓄積することによりSD1遺伝子の発現が誘導され,ジベレリンのうち強い活性をもつGA4が効率よく産生されることにより浮きイネの草丈は伸長することがわかった.さらに,浮きイネのSD1遺伝子にみられた変異は栽培イネの祖先にすでに存在し,のちに人類によりバングラデシュにおいて浮きイネの栽培化に利用されたことが明らかにされた.
植物の生存に水は必須であるが,水没してしまうほどの大量の水は植物の生存をおびやかす.バングラデシュ,ミャンマー,タイ,カンボジアといった東南アジアの国々では,毎年,雨期になると継続的な降雨により水かさが増し,最終的に数メートルにもなる洪水が発生する.洪水は長期間におよび,4~5カ月にわたりつづくこともある.このような過酷な環境において多くの植物は生存できないが,アジアの洪水地帯において栽培される“浮きイネ”はユニークな進化をとげることによりこの環境に適応してきた.一般的なイネは洪水が発生すると水没し窒息により溺死してしまう.一方,浮きイネは乾期の浅水環境においては一般的なイネと同じような草姿をしているが,雨期になり水かさが増すと急激に草丈を伸ばして水面から葉を出し酸素を摂取することにより生き延びる1,2).この伸長の能力は非常に高く,1日に25 cmも伸長し,ときには草丈が数メートルにいたることもある.すなわち,浮きイネは完全に水没してしまうような洪水が長期間つづいても,伸長することにより生存に不利な環境を突破する.
これまで,浮きイネの環境への適応にエチレンやジベレリンといった植物ホルモンが関与することが考えられていたが,その分子機構や起源については明らかにされていなかった1,2).この研究において,筆者らは,浮きイネの環境への適応に関する遺伝子の同定をつうじ,その分子機構および進化的な起源の解明を試みた.
浮きイネの深水に応答した背丈の伸長には節間の急激な伸長が貢献することが知られている1,2).そこで,ゲノムワイド関連解析(genome wide association study:GWAS)により,浮きイネの深水に応答した節間の伸長を制御する遺伝子の探索を試みた.ゲノムワイド関連解析は,多数の系統についてゲノム塩基配列のデータと形質のデータを統計解析し,それぞれの形質に関連する遺伝子について染色体における位置を検出する手法である3).まず,アジア各国からさまざまな浮きイネ性をもつ68品種のイネを収集した.おのおのの品種について7日間の深水処理を施し,伸長したすべての節間の長さを合計した総節間長を測定した.おのおのの品種についてDNAマイクロアレイを用いてDNAマーカーのデータを取得し,得られた総節間長とのあいだでゲノムワイド関連解析を実施した結果,総節間長と強い相関のある領域が第1染色体に見い出された.そののち,非浮きイネと浮きイネの2品種の交配から得られた交雑集団を用いた高密度の連鎖解析により,第1染色体に存在する総節間長を制御する原因遺伝子を探索した.その結果,植物ホルモンであるジベレリンの合成にかかわるSD1(SEMIDWARF1)遺伝子が同定された.
ゲノムワイド関連解析に用いたイネの品種においてSD1遺伝子の塩基配列を比較したところ,高い総節間長の値をもつ浮きイネに共通して17の多型からなる変異がみられ,この変異をもつハプロタイプをDWH(deepwater rice-specific haplotype)と名づけた.以前の研究により,浮きイネの節間の伸長を促進的に制御する別の遺伝子として,SK1遺伝子およびSK2遺伝子とよばれる転写因子をコードする遺伝子が同定されていた4).そこで,DWHとSK1遺伝子およびSK2遺伝子との関連を調べた結果,DWHの関与する深水に応答した節間の伸長の促進にはSK1遺伝子およびSK2遺伝子が必須であり,さらに,DWHはSK1およびSK2と相乗的に機能することが明らかにされた.
DWHをもつイネともたないイネについて深水に応答したSD1遺伝子の発現を比較した.その結果,DWHをもたないイネにおいては深水処理によりSD1遺伝子の発現に変化はみられなかったのに対し,DWHをもつイネにおいては深水処理の1時間のちからSD1遺伝子の発現が顕著に上昇した.一方,イネは深水条件におかれるとガス状の植物ホルモンであるエチレンを体内に蓄積する1,2).また,浮きイネにエチレンを処理すると深水条件でなくとも節間の伸長がひき起こされるため,深水に応答した節間の伸長にはエチレンの関与することが予想されていた1,2).そこで,SD1遺伝子の発現におけるエチレンの処理の効果を比較したところ,DWHをもつイネにおいてのみSD1遺伝子の発現の顕著な上昇が観察された.さらなる分子生物学的な解析により,エチレンシグナル伝達に関与する転写因子EIN3 5) のイネにおけるホモログであるOsEIL1aが,SD1遺伝子のプロモーターに直接的に結合することによりSD1遺伝子の発現が誘導されることが明らかにされ,これをエチレン-ジベレリンリレーと名づけた(図1).また,SK1遺伝子およびSK2遺伝子もOsEIL1aにより直接的に発現が誘導されると考えられていたため4),SD1遺伝子とSK1遺伝子およびSK2遺伝子はともにOsEIL1aにより発現が制御されるといえた.さらに,DWHをもたないイネにおいてもOsEIL1aのSD1遺伝子のプロモーターへの結合能が検出されたことから,イネにはSD1遺伝子の発現を抑制する機構が存在し,DWHはその抑制を弱める効果のあることが考えられた.
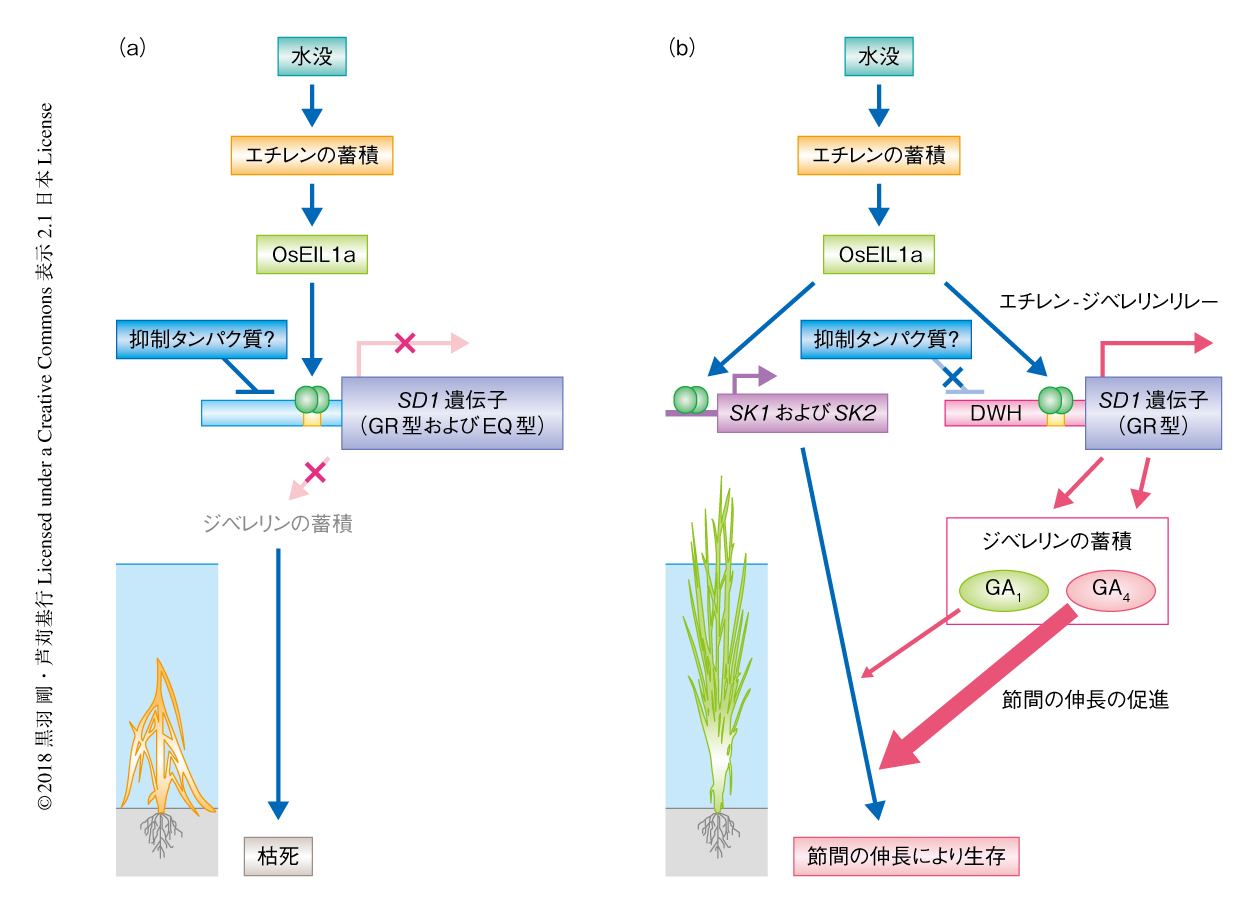
SD1遺伝子の産物であるSD1は,ジベレリン合成経路のうちジベレリン20酸化酵素の活性をもち,植物の体内において生理活性をもつ活性型のジベレリンであるGA1およびGA4の両方の合成にかかわる6).SD1遺伝子のタンパク質コード領域には2箇所の変異があり,イネのSD1にはEQ型およびGR型とよばれる2種類のタイプが存在する7).DWHをもつイネはすべてGR型のSD1をもっていたことから,EQ型のSD1およびGR型のSD1の酵素活性を比較したところ,GR型のSD1の酵素活性はEQ型のSD1よりも圧倒的に高かった.また,イネは通常の栄養成長の段階においてはおもにGA1を合成するが,GR型のSD1はGA1よりもGA4を約20倍も多く合成する能力をもっていた.さらに,浮きイネにおいては深水処理によりGA4の内生量が顕著に増加したとともに,GA4はGA1に比べ浮きイネの節間の伸長を約10倍も強くひき起こすことが外生投与の実験により明らかにされた.以上の結果から,浮きイネは水没することによりエチレンを介し高活性型であるGR型のSD1を多量に産生してGA4を効率よく産生し,節間の急激な伸長をひき起こすことが明らかにされた(図1).
浮きイネのもつハプロタイプであるDWHを手がかりに,さまざまなイネのゲノム塩基配列の情報を比較することにより浮きイネの起源にせまった.まず,アジアのさまざまな地域において栽培されているイネについてSD1遺伝子の塩基配列を比較した結果,DWHはバングラデシュの浮きイネにのみ存在する,栽培イネのなかでは希少なハプロタイプであることが明らかにされた.アジアの栽培イネは遺伝的な背景によりジャポニカ型やインディカ型といったグループに区分される8).DWHをもちかつ高い総節間長の値をもつ浮きイネは,ジャポニカ型やインディカ型を含む遺伝的に多様な背景をもっていた.これらの結果から,DWHがバングラデシュにおける浮きイネの育種に利用されてきたことが示唆された.
DWHにみられる変異の起源をさぐるため,栽培イネの祖先と考えられている野生イネ(Oryza rufipogon)のゲノム塩基配列の情報を解析した.その結果,バングラデシュのみならず南アジアや東南アジア各国の野生イネの一部においてDWHをもつ系統が見い出された.ゲノム塩基配列のさらなる解析の結果,DWHは野生イネの祖先集団にすでに存在していた多型に由来することが明らかにされた.
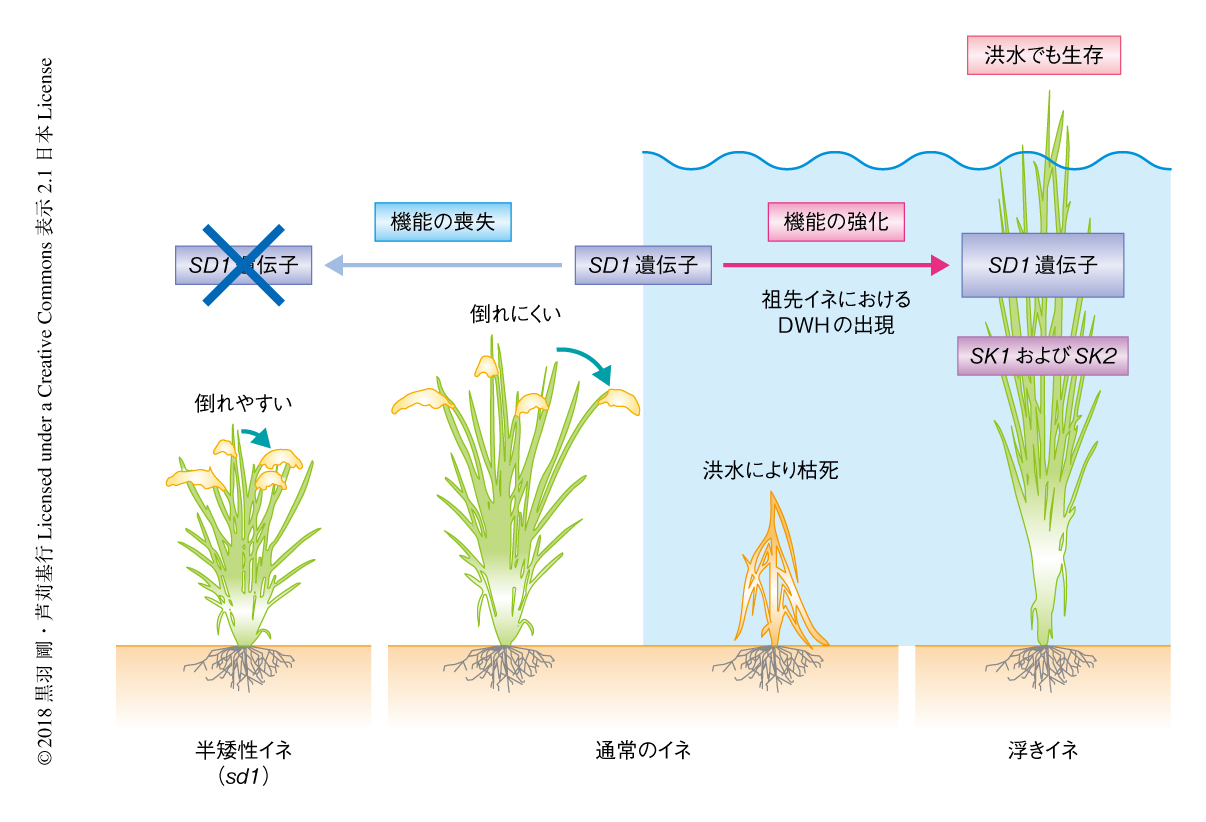
SD1遺伝子は“緑の革命遺伝子”の別名をもち,機能喪失型のsd1遺伝子をもつイネは半矮性イネとよばれ,ジベレリンの含量が減少することにより草丈が低くなり台風などの強風や豪雨でも倒れにくくなることが知られている9)(図2).1960年代以降,化学肥料を十分にあたえても背丈が低く保たれ倒伏せず高収量であることから,sd1遺伝子をもつ栽培イネの品種が数多く作出され,アジアで広く利用されて“緑の革命”となった.一方,この研究により,浮きイネは進化の過程においてSD1遺伝子の機能を逆に強化することにより洪水の際に草丈を急激に伸長させる機能を獲得したことが明らかにされた(図2).以上から,人類は同じ遺伝子の異なる変異を用いて,草丈を低くする方向だけでなく逆に高くする方向への育種にも利用したことが明らかにされた.近年,世界各地において降水量の減少による乾燥および砂漠化や,多雨による大洪水といった異常気象があいついで報告されている.この研究により得られた知見をもとに,浮きイネの環境への適応の機構をさらに解明することにより,東南アジアなどにおける高収量の浮きイネの品種の開発や,環境の変化に応じてイネの草丈を調整する技術の確立などへの応用が期待される.
略歴:2005年 筑波大学大学院生命環境科学研究科 修了,同年 理化学研究所植物科学研究センター 博士研究員,2008年 米国Washington大学 博士研究員,2010年 名古屋大学生物機能開発利用研究センター 博士研究員を経て,2015年より東北大学大学院生命科学研究科 助教.
研究テーマ:植物の環境に応答した形態の変化にかかわる分子機能.
芦苅 基行(Motoyuki Ashikari)
名古屋大学生物機能開発利用研究センター 教授.
研究室URL:http://motoashikari-lab.com/
© 2018 黒羽 剛・芦苅基行 Licensed under CC 表示 2.1 日本
(1東北大学大学院生命科学研究科 植物細胞壁分野,2名古屋大学生物機能開発利用研究センター 高次生体分子機能研究分野)
email:黒羽 剛,芦苅基行
DOI: 10.7875/first.author.2018.071
Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding.
Takeshi Kuroha, Keisuke Nagai, Rico Gamuyao, Diane R. Wang, Tomoyuki Furuta, Masanari Nakamori, Takuya Kitaoka, Keita Adachi, Anzu Minami, Yoshinao Mori, Kiyoshi Mashiguchi, Yoshiya Seto, Shinjiro Yamaguchi, Mikiko Kojima, Hitoshi Sakakibara, Jianzhong Wu, Kaworu Ebana, Nobutaka Mitsuda, Masaru Ohme-Takagi, Shuichi Yanagisawa, Masanori Yamasaki, Ryusuke Yokoyama, Kazuhiko Nishitani, Toshihiro Mochizuki, Gen Tamiya, Susan R. McCouch, Motoyuki Ashikari
Science, 361, 181-186 (2018)
要 約
動物と異なり移動することのできない植物は,過酷な環境に遭遇してもその場所において対応しなければならない.植物はさまざまな環境の変化に対し適応する機能を獲得することによりこのような難題を克服してきた.なかでも,“浮きイネ”とよばれるイネは定期的に発生する洪水に対し草丈を伸ばすというユニークな戦略により生き延びる.この研究において,筆者らは,浮きイネの洪水への適応にかかわる遺伝子として,植物ホルモンであるジベレリンを合成する酵素をコードするSD1遺伝子を同定した.また,洪水により別の植物ホルモンであるエチレンが蓄積することによりSD1遺伝子の発現が誘導され,ジベレリンのうち強い活性をもつGA4が効率よく産生されることにより浮きイネの草丈は伸長することがわかった.さらに,浮きイネのSD1遺伝子にみられた変異は栽培イネの祖先にすでに存在し,のちに人類によりバングラデシュにおいて浮きイネの栽培化に利用されたことが明らかにされた.
はじめに
植物の生存に水は必須であるが,水没してしまうほどの大量の水は植物の生存をおびやかす.バングラデシュ,ミャンマー,タイ,カンボジアといった東南アジアの国々では,毎年,雨期になると継続的な降雨により水かさが増し,最終的に数メートルにもなる洪水が発生する.洪水は長期間におよび,4~5カ月にわたりつづくこともある.このような過酷な環境において多くの植物は生存できないが,アジアの洪水地帯において栽培される“浮きイネ”はユニークな進化をとげることによりこの環境に適応してきた.一般的なイネは洪水が発生すると水没し窒息により溺死してしまう.一方,浮きイネは乾期の浅水環境においては一般的なイネと同じような草姿をしているが,雨期になり水かさが増すと急激に草丈を伸ばして水面から葉を出し酸素を摂取することにより生き延びる1,2).この伸長の能力は非常に高く,1日に25 cmも伸長し,ときには草丈が数メートルにいたることもある.すなわち,浮きイネは完全に水没してしまうような洪水が長期間つづいても,伸長することにより生存に不利な環境を突破する.
これまで,浮きイネの環境への適応にエチレンやジベレリンといった植物ホルモンが関与することが考えられていたが,その分子機構や起源については明らかにされていなかった1,2).この研究において,筆者らは,浮きイネの環境への適応に関する遺伝子の同定をつうじ,その分子機構および進化的な起源の解明を試みた.
1.浮きイネの深水に応答した節間の伸長を制御する遺伝子の同定
浮きイネの深水に応答した背丈の伸長には節間の急激な伸長が貢献することが知られている1,2).そこで,ゲノムワイド関連解析(genome wide association study:GWAS)により,浮きイネの深水に応答した節間の伸長を制御する遺伝子の探索を試みた.ゲノムワイド関連解析は,多数の系統についてゲノム塩基配列のデータと形質のデータを統計解析し,それぞれの形質に関連する遺伝子について染色体における位置を検出する手法である3).まず,アジア各国からさまざまな浮きイネ性をもつ68品種のイネを収集した.おのおのの品種について7日間の深水処理を施し,伸長したすべての節間の長さを合計した総節間長を測定した.おのおのの品種についてDNAマイクロアレイを用いてDNAマーカーのデータを取得し,得られた総節間長とのあいだでゲノムワイド関連解析を実施した結果,総節間長と強い相関のある領域が第1染色体に見い出された.そののち,非浮きイネと浮きイネの2品種の交配から得られた交雑集団を用いた高密度の連鎖解析により,第1染色体に存在する総節間長を制御する原因遺伝子を探索した.その結果,植物ホルモンであるジベレリンの合成にかかわるSD1(SEMIDWARF1)遺伝子が同定された.
2.深水に応答した節間の伸長におけるSD1遺伝子の機能
ゲノムワイド関連解析に用いたイネの品種においてSD1遺伝子の塩基配列を比較したところ,高い総節間長の値をもつ浮きイネに共通して17の多型からなる変異がみられ,この変異をもつハプロタイプをDWH(deepwater rice-specific haplotype)と名づけた.以前の研究により,浮きイネの節間の伸長を促進的に制御する別の遺伝子として,SK1遺伝子およびSK2遺伝子とよばれる転写因子をコードする遺伝子が同定されていた4).そこで,DWHとSK1遺伝子およびSK2遺伝子との関連を調べた結果,DWHの関与する深水に応答した節間の伸長の促進にはSK1遺伝子およびSK2遺伝子が必須であり,さらに,DWHはSK1およびSK2と相乗的に機能することが明らかにされた.
DWHをもつイネともたないイネについて深水に応答したSD1遺伝子の発現を比較した.その結果,DWHをもたないイネにおいては深水処理によりSD1遺伝子の発現に変化はみられなかったのに対し,DWHをもつイネにおいては深水処理の1時間のちからSD1遺伝子の発現が顕著に上昇した.一方,イネは深水条件におかれるとガス状の植物ホルモンであるエチレンを体内に蓄積する1,2).また,浮きイネにエチレンを処理すると深水条件でなくとも節間の伸長がひき起こされるため,深水に応答した節間の伸長にはエチレンの関与することが予想されていた1,2).そこで,SD1遺伝子の発現におけるエチレンの処理の効果を比較したところ,DWHをもつイネにおいてのみSD1遺伝子の発現の顕著な上昇が観察された.さらなる分子生物学的な解析により,エチレンシグナル伝達に関与する転写因子EIN3 5) のイネにおけるホモログであるOsEIL1aが,SD1遺伝子のプロモーターに直接的に結合することによりSD1遺伝子の発現が誘導されることが明らかにされ,これをエチレン-ジベレリンリレーと名づけた(図1).また,SK1遺伝子およびSK2遺伝子もOsEIL1aにより直接的に発現が誘導されると考えられていたため4),SD1遺伝子とSK1遺伝子およびSK2遺伝子はともにOsEIL1aにより発現が制御されるといえた.さらに,DWHをもたないイネにおいてもOsEIL1aのSD1遺伝子のプロモーターへの結合能が検出されたことから,イネにはSD1遺伝子の発現を抑制する機構が存在し,DWHはその抑制を弱める効果のあることが考えられた.
3.浮きイネにおけるSD1を介したジベレリンの合成の制御
SD1遺伝子の産物であるSD1は,ジベレリン合成経路のうちジベレリン20酸化酵素の活性をもち,植物の体内において生理活性をもつ活性型のジベレリンであるGA1およびGA4の両方の合成にかかわる6).SD1遺伝子のタンパク質コード領域には2箇所の変異があり,イネのSD1にはEQ型およびGR型とよばれる2種類のタイプが存在する7).DWHをもつイネはすべてGR型のSD1をもっていたことから,EQ型のSD1およびGR型のSD1の酵素活性を比較したところ,GR型のSD1の酵素活性はEQ型のSD1よりも圧倒的に高かった.また,イネは通常の栄養成長の段階においてはおもにGA1を合成するが,GR型のSD1はGA1よりもGA4を約20倍も多く合成する能力をもっていた.さらに,浮きイネにおいては深水処理によりGA4の内生量が顕著に増加したとともに,GA4はGA1に比べ浮きイネの節間の伸長を約10倍も強くひき起こすことが外生投与の実験により明らかにされた.以上の結果から,浮きイネは水没することによりエチレンを介し高活性型であるGR型のSD1を多量に産生してGA4を効率よく産生し,節間の急激な伸長をひき起こすことが明らかにされた(図1).
4.浮きイネのSD1遺伝子にみられる変異の起源
浮きイネのもつハプロタイプであるDWHを手がかりに,さまざまなイネのゲノム塩基配列の情報を比較することにより浮きイネの起源にせまった.まず,アジアのさまざまな地域において栽培されているイネについてSD1遺伝子の塩基配列を比較した結果,DWHはバングラデシュの浮きイネにのみ存在する,栽培イネのなかでは希少なハプロタイプであることが明らかにされた.アジアの栽培イネは遺伝的な背景によりジャポニカ型やインディカ型といったグループに区分される8).DWHをもちかつ高い総節間長の値をもつ浮きイネは,ジャポニカ型やインディカ型を含む遺伝的に多様な背景をもっていた.これらの結果から,DWHがバングラデシュにおける浮きイネの育種に利用されてきたことが示唆された.
DWHにみられる変異の起源をさぐるため,栽培イネの祖先と考えられている野生イネ(Oryza rufipogon)のゲノム塩基配列の情報を解析した.その結果,バングラデシュのみならず南アジアや東南アジア各国の野生イネの一部においてDWHをもつ系統が見い出された.ゲノム塩基配列のさらなる解析の結果,DWHは野生イネの祖先集団にすでに存在していた多型に由来することが明らかにされた.
おわりに
SD1遺伝子は“緑の革命遺伝子”の別名をもち,機能喪失型のsd1遺伝子をもつイネは半矮性イネとよばれ,ジベレリンの含量が減少することにより草丈が低くなり台風などの強風や豪雨でも倒れにくくなることが知られている9)(図2).1960年代以降,化学肥料を十分にあたえても背丈が低く保たれ倒伏せず高収量であることから,sd1遺伝子をもつ栽培イネの品種が数多く作出され,アジアで広く利用されて“緑の革命”となった.一方,この研究により,浮きイネは進化の過程においてSD1遺伝子の機能を逆に強化することにより洪水の際に草丈を急激に伸長させる機能を獲得したことが明らかにされた(図2).以上から,人類は同じ遺伝子の異なる変異を用いて,草丈を低くする方向だけでなく逆に高くする方向への育種にも利用したことが明らかにされた.近年,世界各地において降水量の減少による乾燥および砂漠化や,多雨による大洪水といった異常気象があいついで報告されている.この研究により得られた知見をもとに,浮きイネの環境への適応の機構をさらに解明することにより,東南アジアなどにおける高収量の浮きイネの品種の開発や,環境の変化に応じてイネの草丈を調整する技術の確立などへの応用が期待される.
文 献
- Hattori, Y., Nagai, K. & Ashikari, M.: Rice growth adapting to deepwater. Curr. Opin. Plant Biol., 14, 100-105 (2011)[PubMed]
- Kende, H., van der Knaap, E. & Cho, H. T.: Deepwater rice: a model plant to study stem elongation. Plant Physiol., 118, 1105-1110 (1998)[PubMed]
- Myles, S., Peiffer, J., Brown, P. J. et al.: Association mapping: critical considerations shift from genotyping to experimental design. Plant Cell, 21, 2194-2202 (2009)[PubMed]
- Hattori, Y., Nagai, K., Furukawa, S. et al.: The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature, 460, 1026-1030 (2009)[PubMed]
- Ju, C. & Chang, C.: Mechanistic insights in ethylene perception and signal transduction. Plant Physiol., 169, 85-95 (2015)[PubMed]
- Yamaguchi, S.: Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol., 59, 225-251 (2008)[PubMed]
- Asano, K., Yamasaki, M., Takuno, S. et al.: Artificial selection for a green revolution gene during japonica rice domestication. Proc. Natl. Acad. Sci. USA, 108, 11034-11039 (2011)[PubMed]
- Zhao, K., Tung, C. W., Eizenga, G. C. et al.: Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun., 2, 467 (2011)[PubMed]
- Sasaki, A., Ashikari, M., Ueguchi-Tanaka, M. et al.: Green revolution: a mutant gibberellin-synthesis gene in rice. Nature, 416, 701-702 (2002)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2005年 筑波大学大学院生命環境科学研究科 修了,同年 理化学研究所植物科学研究センター 博士研究員,2008年 米国Washington大学 博士研究員,2010年 名古屋大学生物機能開発利用研究センター 博士研究員を経て,2015年より東北大学大学院生命科学研究科 助教.
研究テーマ:植物の環境に応答した形態の変化にかかわる分子機能.
芦苅 基行(Motoyuki Ashikari)
名古屋大学生物機能開発利用研究センター 教授.
研究室URL:http://motoashikari-lab.com/
© 2018 黒羽 剛・芦苅基行 Licensed under CC 表示 2.1 日本
