乾燥ストレス応答において根から葉への長距離のシグナル伝達にかかわるペプチドはアブシジン酸を介して気孔の開閉を制御する
高橋史憲・篠崎一雄
(理化学研究所環境資源科学研究センター 機能開発研究グループ)
email:高橋史憲,篠崎一雄
DOI: 10.7875/first.author.2018.044
A small peptide modulates stomatal control via abscisic acid in long-distance signalling.
Fuminori Takahashi, Takehiro Suzuki, Yuriko Osakabe, Shigeyuki Betsuyaku, Yuki Kondo, Naoshi Dohmae, Hiroo Fukuda, Kazuko Yamaguchi-Shinozaki, Kazuo Shinozaki
Nature, 556, 235-238 (2018)
陸上植物における乾燥ストレスのもとでの根から地上部へのシグナル伝達は,蒸散による水分の損失をふせぎ,水分の欠乏に適応するために重要である.植物ホルモンであるアブシジン酸は水分の損失をふせぐための気孔の閉鎖にかかわることが知られている.しかし,葉におけるアブシジン酸の蓄積をひき起こす移動性のシグナル伝達物質については,いまだ明らかにされていない.この研究において,筆者らは,シロイヌナズナにおいてCLE25ペプチドが維管束柔組織を介して乾燥ストレスのシグナルを伝達し,葉においてその受容体であるBAMと結合することによりアブシジン酸の合成に影響を促進し,気孔を閉鎖することにより水分の蒸散を制御することを明らかにした.CLE25遺伝子は維管束柔組織に発現しており,乾燥ストレスに応答し根においてその発現が上昇した.根において合成されたCLE25ペプチドは乾燥ストレスに依存して根から葉へと移動し,葉においてアブシジン酸の蓄積を制御することにより気孔の閉鎖をひき起こし,乾燥ストレスへの耐性を上昇させた.BAMはCLE25ペプチドによりひき起こされる葉における乾燥ストレス応答に必要であり,CLE25-BAM複合体は乾燥ストレス応答において離れた組織のあいだでシグナルをやりとりするための移動性のシグナル伝達物質のひとつとして機能すると考えられた.この研究により,乾燥などに対する環境ストレス応答におけるシグナル伝達,とくに,長距離のシグナル伝達にペプチドがはたらくことが明らかにされ,複雑な植物の環境ストレス応答に新たな展開をもたらすことになった.
植物は芽をだした場所で一生をすごす.そのため,葉や根といったさまざまな組織や器官において環境の変化を認識し,細胞のレベルにおいて遺伝子の発現や代謝物質を変化させて外界に適応しながら生存する.とくに,乾燥ストレスのもとでは,土壌および大気の環境の変化を適切に認識し,蒸散による水分の損失をふせいだり,全身での乾燥ストレスへの耐性を獲得したりするため,根と葉のあいだでさまざまなシグナルが伝達されると考えられてきた1,2).植物の乾燥ストレス応答における重要な因子として,植物ホルモンのひとつであるアブシジン酸が広く知られている.アブシジン酸は乾燥ストレスを感知した植物の葉において合成され,葉の気孔の閉鎖を促進し体内から水分が失われるのをふせぐ3).また,アブシジン酸は乾燥ストレスへの耐性にかかわる遺伝子の発現の制御も担う.
乾燥ストレスのもと葉においてアブシジン酸を合成するのに必須な酵素としてNCED3が知られている.NCED3遺伝子は,シロイヌナズナのもつ9つのNCED遺伝子のなかで唯一,乾燥ストレスに依存して発現が上昇するとともに,タンパク質のレベルにて葉や胚軸の維管束柔組織に蓄積する4,5).NCED3遺伝子を破壊した植物は乾燥ストレスに弱く,過剰に発現した植物は乾燥ストレスに強いことから,NCED3遺伝子の発現の制御は乾燥ストレス応答および乾燥ストレスへの耐性の獲得に重要であることがわかる.
これまで,乾燥ストレスのもとで根から地上部へと移動するシグナル伝達因子として,水分の膨圧,Ca2+シグナル,アブシジン酸自体の移動などが知られていた.しかし,植物が土壌における水分の減少による乾燥ストレスを根において感受したのち,葉においてNCED3遺伝子の発現を介したアブシジン酸の合成が促進されるまでの分子機構については,ほとんど解明されていなかった.
近年,シロイヌナズナのゲノムにはsORF(small ORF)とよばれる300 bp以下の短い遺伝子が7000以上も存在することが示唆されている6).また,植物にも分泌型のペプチドが存在し,成長や花成の制御にかかわることが報告されている7,8).なかでも,CLV3ペプチドを含むCLEペプチドファミリーに属するいくつかのペプチドについては研究が進んでいる.CLV3ペプチドは茎頂分裂組織における恒常性の維持にかかわる.また,TDIFペプチド,CLE41ペプチド,CLE44ペプチドは維管束の形態形成および分化にかかわる.しかしこれまで,このようなペプチドやsORFが環境ストレス応答にかかわるという報告はなかった.
そこで,タイリングアレイ法により環境ストレスに応答して発現が上昇する既知のペプチド遺伝子およびsORFを探索した.その結果,CLEペプチドをコードするいくつかの遺伝子が環境ストレスに対する応答性を示すことが見い出された.このことから,CLEペプチドファミリーが環境ストレス応答にも関与する可能性が考えられた.そこで,27種類のCLEペプチドを人工合成して根に添加し,葉におけるNCED3遺伝子の発現の上昇,葉におけるアブシジン酸の蓄積,気孔の開閉について調べた.その結果,根へのCLE25ペプチドの添加により葉におけるアブシジン酸の蓄積および気孔の閉鎖がひき起こされた.これらの結果から,根で吸収されたCLE25ペプチドが葉に輸送されてNCED3遺伝子の発現を誘導し,葉においてアブシジン酸の蓄積を介した気孔の閉鎖にかかわることが示唆された.
CLE25遺伝子が発現する組織について解析した結果,根および葉の維管束柔組織,とくに,維管束前駆細胞において強く発現していることが明らかにされた.維管束柔組織はNCED3を含むアブシジン酸合成酵素,および,アブシジン酸のトランスポーターが強く発現する組織であることを考えると,この結果から,CLE25ペプチドおよび一連のアブシジン酸合成経路が維管束柔組織において機能することが示唆された.また,乾燥ストレスのもとでのCLE25遺伝子の発現の変動を根および葉において調べた結果,根においてのみ,乾燥ストレスによる顕著な誘導性が示された.
植物体におけるCLE25ペプチドの機能を明らかにするため,CRISPR-Cas9法によるゲノム編集技術を用いてcle25破壊植物を作製した.cle25破壊植物においては,乾燥ストレスによる誘導性を示すNCED3遺伝子や,アブシジン酸シグナル伝達経路のマーカー遺伝子であるLEA遺伝子およびRD29B遺伝子の発現が抑制されていた.また,cle25破壊植物は葉におけるアブシジン酸の蓄積量が低下しており,乾燥ストレスに弱かったことから,CLE25ペプチドはNCED3遺伝子の発現を介したアブシジン酸の合成および乾燥ストレスへの耐性を制御することが明らかにされた.一方,幼植物体を用いて脱水ストレスのもとでの蒸散量を測定したところ,cle25破壊植物はアブシジン酸の蓄積が確認された乾燥処理時間よりも早い時間帯において,野生型よりも高い蒸散量を示した.この結果から,CLE25ペプチドはアブシジン酸の合成を介した気孔の閉鎖だけでなく,ほかのシグナル伝達経路の関与する乾燥ストレス応答および乾燥ストレスへの耐性の獲得にもかかわることが示唆された.今後は,水分の膨圧やCa2+シグナルとの関係を解析する必要があるだろう.
CLE25ペプチドは乾燥ストレスに依存して根において発現が上昇したことから,ひとつの可能性として,CLE25ペプチドは根におけるアブシジン酸の合成に影響をおよぼし,根において合成されたアブシジン酸が地上部へと移動してNCED3遺伝子の発現および気孔の閉鎖をひき起こすことが考えられた.そこで,接木技術を使ってアブシジン酸の合成欠損植物と野生型の植物とを接木し,根からCLE25ペプチドを吸収させたのち,葉におけるNCED3遺伝子の発現について解析した.その結果,根がアブシジン酸の合成欠損植物であっても葉におけるNCED3遺伝子の発現の上昇が確認された.このことから,CLE25ペプチドは根におけるアブシジン酸の合成に依存せず,CLE25ペプチドは地上部へと移動し葉においてNCED3遺伝子の発現を介したアブシジン酸の合成を促進することが示唆された.そこで,野生型の植物とcle25破壊植物とを接木し内在性のCLE25ペプチドの移動を測定した結果,根において合成されたCLE25ペプチドは乾燥ストレスに依存して根から葉に移動することが証明された(図1).また,根から地上部へと移動したCLE25ペプチドは,葉においてNCED3遺伝子の発現にかかわることも明らかにされた.また,葉において発現したCLE25ペプチドもNCED3遺伝子の発現にかかわっていた.CLE25遺伝子の組織に特異的な発現の解析において,葉の維管束柔組織においてもCLE25遺伝子の発現がみられたことから,CLE25ペプチドは根と葉をつなぐ長距離のシグナル伝達因子としてだけでなく,葉における局所的な乾燥ストレス応答にもかかわると考えられた.
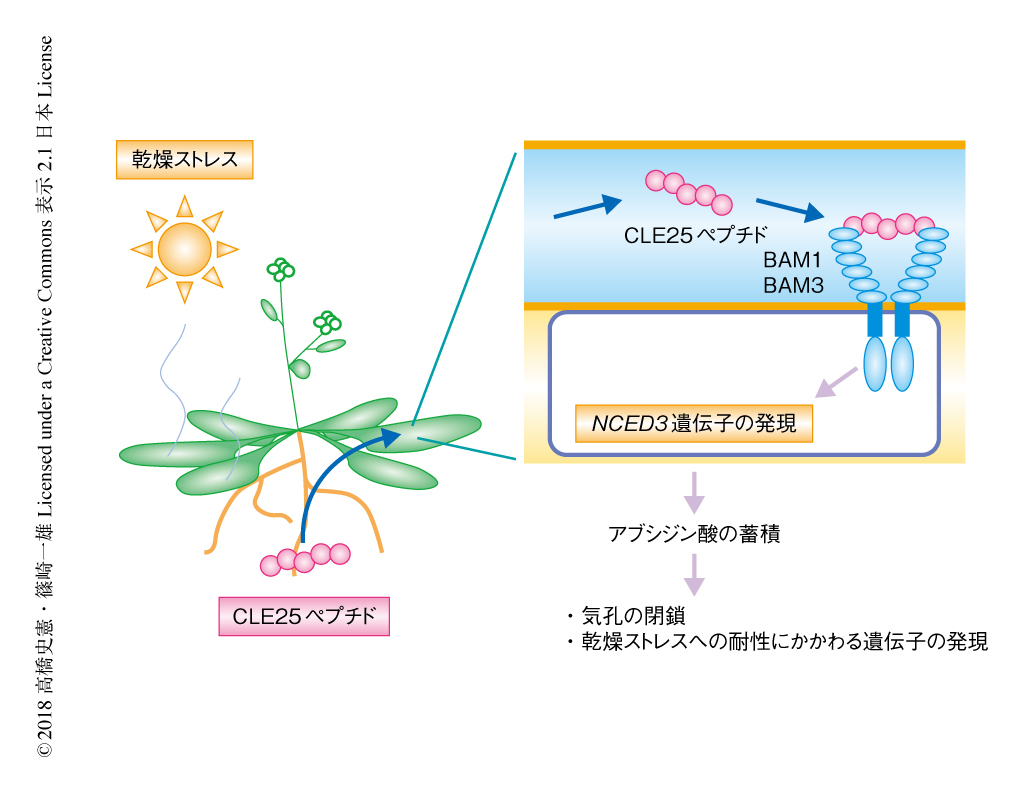
CLE25ペプチドを受容する受容体を探索した.これまでに報告されているCLEペプチドファミリーのいくつかは,サブクラスIXとよばれる一群の特徴をもつ受容体キナーゼと結合することが知られていた.そこで,このサブクラスを対象としてスクリーニングした結果,BAM1およびBAM3の2つがNCED3遺伝子の発現,アブシジン酸の蓄積,乾燥ストレスへの耐性の獲得にかかわることが見い出された.一方,これまで,BAM1はBAM2と塩基配列の相同性が高く,ともに葉の形態形成にかかわることが報告されていたが,BAM2は乾燥ストレス応答にはかかわらなかった.このことから,BAM1とBAM2は発現レベルでの組織の特異性,ヘテロ二量体の形成,リガンドの認識性などを介した,機能的な役割の分担のあることが示唆された.さらに,bam1 bam3二重変異植物を用いた接木の実験から,CLE25ペプチドが葉においてNCED3遺伝子の発現を誘導するにはBAM1およびBAM3が必要であることが証明された(図1).
この研究により,移動性のCLE25ペプチドが根と葉という離れた組織のあいだで乾燥ストレスのシグナルをやりとりするための要として機能しており,植物は環境ストレス応答においても全身的なシグナル伝達系を使った器官のあいだのコミュニケーション系をもつことが示された.とくに,CLE25ペプチドは乾燥ストレスに依存して細胞外へと放出されることから,CLE25-BAM複合体は外部の環境ストレスを統合的に感知して器官のあいだでシグナルを長距離で伝達する系の一部であると考えられた(図1).ストレスの長距離でのシグナルとして,ほかにも水分の欠乏による膨圧の変化やアブシジン酸自体もかかわることが考えられており,ペプチドの長距離の輸送との関係の解明が必要である.また,CLE25ペプチドは作物として重要であるイネ,コムギ,トマト,ダイズのゲノムにも保存されていることから,CLE25ペプチドによる乾燥ストレスの認識の機構,および,乾燥ストレスのシグナルを全身に伝達する系が広く保存されていることが示唆された.今後,移動性のペプチドによる乾燥ストレス応答の機構をさらに解明していくことにより,植物の器官のあいだのシグナル伝達に関する分子レベルでの理解が深まるだけでなく,応用への発展にも寄与することを期待したい.
略歴:2007年 筑波大学大学院生命環境科学研究科博士課程 修了,同年 理化学研究所植物科学研究センター 特別研究員,オーストラリアAdelaide大学 客員研究員,理化学研究所バイオマス工学研究プログラム 研究員を経て,2018年より理化学研究所環境資源科学研究センター 研究員.
研究テーマ:植物の環境ストレス応答にかかわる,離れた組織あるいは器官のあいだをつなぐシグナル伝達ネットワーク.
篠崎 一雄(Kazuo Shinozaki)
理化学研究所環境資源科学研究センター グループディレクター.
研究室URL:http://genediscovery.riken.jp/
© 2018 高橋史憲・篠崎一雄 Licensed under CC 表示 2.1 日本
(理化学研究所環境資源科学研究センター 機能開発研究グループ)
email:高橋史憲,篠崎一雄
DOI: 10.7875/first.author.2018.044
A small peptide modulates stomatal control via abscisic acid in long-distance signalling.
Fuminori Takahashi, Takehiro Suzuki, Yuriko Osakabe, Shigeyuki Betsuyaku, Yuki Kondo, Naoshi Dohmae, Hiroo Fukuda, Kazuko Yamaguchi-Shinozaki, Kazuo Shinozaki
Nature, 556, 235-238 (2018)
要 約
陸上植物における乾燥ストレスのもとでの根から地上部へのシグナル伝達は,蒸散による水分の損失をふせぎ,水分の欠乏に適応するために重要である.植物ホルモンであるアブシジン酸は水分の損失をふせぐための気孔の閉鎖にかかわることが知られている.しかし,葉におけるアブシジン酸の蓄積をひき起こす移動性のシグナル伝達物質については,いまだ明らかにされていない.この研究において,筆者らは,シロイヌナズナにおいてCLE25ペプチドが維管束柔組織を介して乾燥ストレスのシグナルを伝達し,葉においてその受容体であるBAMと結合することによりアブシジン酸の合成に影響を促進し,気孔を閉鎖することにより水分の蒸散を制御することを明らかにした.CLE25遺伝子は維管束柔組織に発現しており,乾燥ストレスに応答し根においてその発現が上昇した.根において合成されたCLE25ペプチドは乾燥ストレスに依存して根から葉へと移動し,葉においてアブシジン酸の蓄積を制御することにより気孔の閉鎖をひき起こし,乾燥ストレスへの耐性を上昇させた.BAMはCLE25ペプチドによりひき起こされる葉における乾燥ストレス応答に必要であり,CLE25-BAM複合体は乾燥ストレス応答において離れた組織のあいだでシグナルをやりとりするための移動性のシグナル伝達物質のひとつとして機能すると考えられた.この研究により,乾燥などに対する環境ストレス応答におけるシグナル伝達,とくに,長距離のシグナル伝達にペプチドがはたらくことが明らかにされ,複雑な植物の環境ストレス応答に新たな展開をもたらすことになった.
はじめに
植物は芽をだした場所で一生をすごす.そのため,葉や根といったさまざまな組織や器官において環境の変化を認識し,細胞のレベルにおいて遺伝子の発現や代謝物質を変化させて外界に適応しながら生存する.とくに,乾燥ストレスのもとでは,土壌および大気の環境の変化を適切に認識し,蒸散による水分の損失をふせいだり,全身での乾燥ストレスへの耐性を獲得したりするため,根と葉のあいだでさまざまなシグナルが伝達されると考えられてきた1,2).植物の乾燥ストレス応答における重要な因子として,植物ホルモンのひとつであるアブシジン酸が広く知られている.アブシジン酸は乾燥ストレスを感知した植物の葉において合成され,葉の気孔の閉鎖を促進し体内から水分が失われるのをふせぐ3).また,アブシジン酸は乾燥ストレスへの耐性にかかわる遺伝子の発現の制御も担う.
乾燥ストレスのもと葉においてアブシジン酸を合成するのに必須な酵素としてNCED3が知られている.NCED3遺伝子は,シロイヌナズナのもつ9つのNCED遺伝子のなかで唯一,乾燥ストレスに依存して発現が上昇するとともに,タンパク質のレベルにて葉や胚軸の維管束柔組織に蓄積する4,5).NCED3遺伝子を破壊した植物は乾燥ストレスに弱く,過剰に発現した植物は乾燥ストレスに強いことから,NCED3遺伝子の発現の制御は乾燥ストレス応答および乾燥ストレスへの耐性の獲得に重要であることがわかる.
これまで,乾燥ストレスのもとで根から地上部へと移動するシグナル伝達因子として,水分の膨圧,Ca2+シグナル,アブシジン酸自体の移動などが知られていた.しかし,植物が土壌における水分の減少による乾燥ストレスを根において感受したのち,葉においてNCED3遺伝子の発現を介したアブシジン酸の合成が促進されるまでの分子機構については,ほとんど解明されていなかった.
1.乾燥ストレス応答にかかわるペプチドの探索
近年,シロイヌナズナのゲノムにはsORF(small ORF)とよばれる300 bp以下の短い遺伝子が7000以上も存在することが示唆されている6).また,植物にも分泌型のペプチドが存在し,成長や花成の制御にかかわることが報告されている7,8).なかでも,CLV3ペプチドを含むCLEペプチドファミリーに属するいくつかのペプチドについては研究が進んでいる.CLV3ペプチドは茎頂分裂組織における恒常性の維持にかかわる.また,TDIFペプチド,CLE41ペプチド,CLE44ペプチドは維管束の形態形成および分化にかかわる.しかしこれまで,このようなペプチドやsORFが環境ストレス応答にかかわるという報告はなかった.
そこで,タイリングアレイ法により環境ストレスに応答して発現が上昇する既知のペプチド遺伝子およびsORFを探索した.その結果,CLEペプチドをコードするいくつかの遺伝子が環境ストレスに対する応答性を示すことが見い出された.このことから,CLEペプチドファミリーが環境ストレス応答にも関与する可能性が考えられた.そこで,27種類のCLEペプチドを人工合成して根に添加し,葉におけるNCED3遺伝子の発現の上昇,葉におけるアブシジン酸の蓄積,気孔の開閉について調べた.その結果,根へのCLE25ペプチドの添加により葉におけるアブシジン酸の蓄積および気孔の閉鎖がひき起こされた.これらの結果から,根で吸収されたCLE25ペプチドが葉に輸送されてNCED3遺伝子の発現を誘導し,葉においてアブシジン酸の蓄積を介した気孔の閉鎖にかかわることが示唆された.
2.CLE25ペプチドは乾燥ストレス応答および乾燥ストレスへの耐性にかかわる
CLE25遺伝子が発現する組織について解析した結果,根および葉の維管束柔組織,とくに,維管束前駆細胞において強く発現していることが明らかにされた.維管束柔組織はNCED3を含むアブシジン酸合成酵素,および,アブシジン酸のトランスポーターが強く発現する組織であることを考えると,この結果から,CLE25ペプチドおよび一連のアブシジン酸合成経路が維管束柔組織において機能することが示唆された.また,乾燥ストレスのもとでのCLE25遺伝子の発現の変動を根および葉において調べた結果,根においてのみ,乾燥ストレスによる顕著な誘導性が示された.
植物体におけるCLE25ペプチドの機能を明らかにするため,CRISPR-Cas9法によるゲノム編集技術を用いてcle25破壊植物を作製した.cle25破壊植物においては,乾燥ストレスによる誘導性を示すNCED3遺伝子や,アブシジン酸シグナル伝達経路のマーカー遺伝子であるLEA遺伝子およびRD29B遺伝子の発現が抑制されていた.また,cle25破壊植物は葉におけるアブシジン酸の蓄積量が低下しており,乾燥ストレスに弱かったことから,CLE25ペプチドはNCED3遺伝子の発現を介したアブシジン酸の合成および乾燥ストレスへの耐性を制御することが明らかにされた.一方,幼植物体を用いて脱水ストレスのもとでの蒸散量を測定したところ,cle25破壊植物はアブシジン酸の蓄積が確認された乾燥処理時間よりも早い時間帯において,野生型よりも高い蒸散量を示した.この結果から,CLE25ペプチドはアブシジン酸の合成を介した気孔の閉鎖だけでなく,ほかのシグナル伝達経路の関与する乾燥ストレス応答および乾燥ストレスへの耐性の獲得にもかかわることが示唆された.今後は,水分の膨圧やCa2+シグナルとの関係を解析する必要があるだろう.
3.CLE25ペプチドは根から葉へと移動してアブシジン酸の合成を制御する
CLE25ペプチドは乾燥ストレスに依存して根において発現が上昇したことから,ひとつの可能性として,CLE25ペプチドは根におけるアブシジン酸の合成に影響をおよぼし,根において合成されたアブシジン酸が地上部へと移動してNCED3遺伝子の発現および気孔の閉鎖をひき起こすことが考えられた.そこで,接木技術を使ってアブシジン酸の合成欠損植物と野生型の植物とを接木し,根からCLE25ペプチドを吸収させたのち,葉におけるNCED3遺伝子の発現について解析した.その結果,根がアブシジン酸の合成欠損植物であっても葉におけるNCED3遺伝子の発現の上昇が確認された.このことから,CLE25ペプチドは根におけるアブシジン酸の合成に依存せず,CLE25ペプチドは地上部へと移動し葉においてNCED3遺伝子の発現を介したアブシジン酸の合成を促進することが示唆された.そこで,野生型の植物とcle25破壊植物とを接木し内在性のCLE25ペプチドの移動を測定した結果,根において合成されたCLE25ペプチドは乾燥ストレスに依存して根から葉に移動することが証明された(図1).また,根から地上部へと移動したCLE25ペプチドは,葉においてNCED3遺伝子の発現にかかわることも明らかにされた.また,葉において発現したCLE25ペプチドもNCED3遺伝子の発現にかかわっていた.CLE25遺伝子の組織に特異的な発現の解析において,葉の維管束柔組織においてもCLE25遺伝子の発現がみられたことから,CLE25ペプチドは根と葉をつなぐ長距離のシグナル伝達因子としてだけでなく,葉における局所的な乾燥ストレス応答にもかかわると考えられた.
4.CLE25ペプチドの受容体であるBAMはCLE25ペプチドにより制御される乾燥ストレス応答にかかわる
CLE25ペプチドを受容する受容体を探索した.これまでに報告されているCLEペプチドファミリーのいくつかは,サブクラスIXとよばれる一群の特徴をもつ受容体キナーゼと結合することが知られていた.そこで,このサブクラスを対象としてスクリーニングした結果,BAM1およびBAM3の2つがNCED3遺伝子の発現,アブシジン酸の蓄積,乾燥ストレスへの耐性の獲得にかかわることが見い出された.一方,これまで,BAM1はBAM2と塩基配列の相同性が高く,ともに葉の形態形成にかかわることが報告されていたが,BAM2は乾燥ストレス応答にはかかわらなかった.このことから,BAM1とBAM2は発現レベルでの組織の特異性,ヘテロ二量体の形成,リガンドの認識性などを介した,機能的な役割の分担のあることが示唆された.さらに,bam1 bam3二重変異植物を用いた接木の実験から,CLE25ペプチドが葉においてNCED3遺伝子の発現を誘導するにはBAM1およびBAM3が必要であることが証明された(図1).
おわりに
この研究により,移動性のCLE25ペプチドが根と葉という離れた組織のあいだで乾燥ストレスのシグナルをやりとりするための要として機能しており,植物は環境ストレス応答においても全身的なシグナル伝達系を使った器官のあいだのコミュニケーション系をもつことが示された.とくに,CLE25ペプチドは乾燥ストレスに依存して細胞外へと放出されることから,CLE25-BAM複合体は外部の環境ストレスを統合的に感知して器官のあいだでシグナルを長距離で伝達する系の一部であると考えられた(図1).ストレスの長距離でのシグナルとして,ほかにも水分の欠乏による膨圧の変化やアブシジン酸自体もかかわることが考えられており,ペプチドの長距離の輸送との関係の解明が必要である.また,CLE25ペプチドは作物として重要であるイネ,コムギ,トマト,ダイズのゲノムにも保存されていることから,CLE25ペプチドによる乾燥ストレスの認識の機構,および,乾燥ストレスのシグナルを全身に伝達する系が広く保存されていることが示唆された.今後,移動性のペプチドによる乾燥ストレス応答の機構をさらに解明していくことにより,植物の器官のあいだのシグナル伝達に関する分子レベルでの理解が深まるだけでなく,応用への発展にも寄与することを期待したい.
文 献
- Steudle, E.: The cohesion-tension mechanism and the acquisition of water by plant root. Annu. Rev. Physiol., 52, 847-875 (2001)[PubMed]
- Christmann, A., Grill, E. & Huang, J.: Hydraulic signals in long-distance signaling. Curr. Opin. Plant Biol., 16, 293-300 (2013)[PubMed]
- Kim, T. H., Bohmer, M., Hu, H. et al.: Guard cell signal transduction network: advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol., 61, 561-591 (2010)[PubMed]
- Iuchi, S., Kobayashi, M., Taji, T. et al.: Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J., 27, 325-333 (2001)[PubMed]
- Endo, A., Sawada, Y., Takahashi, H. et al.: Drought induction of Arabidopsis 9-cis-epoxycarotenoid dioxygenase occurs in vascular parenchyma cells. Plant Physiol., 147, 1984-1993 (2008)[PubMed]
- Hanada, K., Zhang, X., Borevitz, J. O. et al.: A large number of novel coding small open reading frames in the intergenic regions of the Arabidopsis thaliana genome are transcribed and/or under purifying selection. Genome Res., 17, 632-640 (2007)[PubMed]
- Matsubayashi, Y.: Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol., 65, 385-413 (2014)[PubMed]
- Betsuyaku, S., Sawa, S. & Yamada, M.: The function of the CLE peptides in plant development and plant-microbe interactions. Arabidopsis Book, 9, e0149 (2011)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2007年 筑波大学大学院生命環境科学研究科博士課程 修了,同年 理化学研究所植物科学研究センター 特別研究員,オーストラリアAdelaide大学 客員研究員,理化学研究所バイオマス工学研究プログラム 研究員を経て,2018年より理化学研究所環境資源科学研究センター 研究員.
研究テーマ:植物の環境ストレス応答にかかわる,離れた組織あるいは器官のあいだをつなぐシグナル伝達ネットワーク.
篠崎 一雄(Kazuo Shinozaki)
理化学研究所環境資源科学研究センター グループディレクター.
研究室URL:http://genediscovery.riken.jp/
© 2018 高橋史憲・篠崎一雄 Licensed under CC 表示 2.1 日本
