機械刺激の受容にもとづく匂い情報の時間コーディング
岩田 遼・今井 猛
(理化学研究所多細胞システム形成研究センター 感覚神経回路形成研究チーム)
email:今井 猛
DOI: 10.7875/first.author.2017.144
Mechanosensory-based phase coding of odor identity in the olfactory bulb.
Ryo Iwata, Hiroshi Kiyonari, Takeshi Imai
Neuron, 96, 1139-1152.e7 (2017)
脳において感覚情報はニューロンの発火の頻度および発火のタイミングにより表現される.哺乳類の嗅球の僧帽細胞においては匂い情報が入力すると発火の頻度が変化するだけでなく発火のタイミングも変化する.この研究において,筆者らは,嗅細胞において受容される匂い刺激と機械刺激が僧帽細胞の発火のタイミングにもとづき区別されることを見い出した.吸気にともない嗅細胞に機械刺激が入力すると嗅球へと伝達され,嗅球の僧帽細胞において呼吸の周期に同期した神経活動の振動が生じた.神経活動の振動の位相すなわち発火のタイミングは吸気の流速を変化させても変わらなかったが,匂い刺激をあたえると変化した.匂い刺激により生じる匂いに固有の発火のタイミングは,複数の呼吸の周期にわたり匂いを嗅がせた場合においても,匂いの濃度を変化させた場合においても,発火の頻度より安定であった.また,発火のタイミングのほうが発火の頻度よりも正確に匂いの種類をコードした.さらに,機械刺激にもとづく神経活動の振動を人為的に消去したところ,匂い刺激に対する発火のタイミングは不正確になった.以上の結果から,匂いの種類に関する情報は発火の頻度よりも発火のタイミングにより安定に表現されること,発火のタイミングの正確性は嗅細胞における機械刺激の受容により保証されることが明らかにされた.
脳における感覚情報の処理においては,外界のノイズにみちた感覚の刺激のなかから特定の情報だけが抽出される.近年,感覚情報はニューロンの発火の頻度の変化により表現されるだけではなく(発火頻度コーディング),発火のタイミングの変化によっても表現されること(時間コーディング)が明らかにされつつある1).しかしながら,感覚情報の処理における発火のタイミングの意義については不明な点が多い.
マウスの嗅覚系において,匂い分子は嗅上皮の嗅細胞により検出される.個々の嗅細胞は約1000種類ある嗅覚受容体のなかから単一の受容体だけを選択的に発現する.同一の嗅覚受容体を発現する嗅細胞は,嗅球において1対の糸球体へと軸索を収斂させる.したがって,匂い情報は嗅球にある約2000個の糸球体の時空間的な神経活動のパターンにより表現される.糸球体において,嗅細胞の軸索は投射ニューロンである僧帽細胞にくわえ,糸球体層の介在ニューロンと接続する2).僧帽細胞は嗅皮質へと軸索を投射し匂い情報を伝達する.
空気に含まれる匂い分子は,呼吸あるいは積極的な吸気にともなう空気の流れにより嗅上皮へと到達する.げっ歯類は1回の呼吸で匂いの種類を識別できるため3),そのあいだの神経活動のパターンに匂いを識別できるだけの十分な情報が含まれると考えられる.僧帽細胞においては,匂い刺激によりその発火の頻度が変化するだけでなく,匂いの種類に固有なタイミングで発火が生じる4)(図1a).さらに近年,僧帽細胞の発火のタイミングを光遺伝学的な手法により変化させるとマウスの個体が実際にそれを識別できることや5),2つの糸球体をさまざまなタイミングで光遺伝学的な手法により刺激すると嗅皮質における応答が変わることが報告された6).こうしたことから,発火のタイミングが匂い情報の処理になんらかの役割をはたすと考えられるようになった.しかし,匂い情報の処理における発火のタイミングの役割やその制御機構についてはよくわかっていなかった.
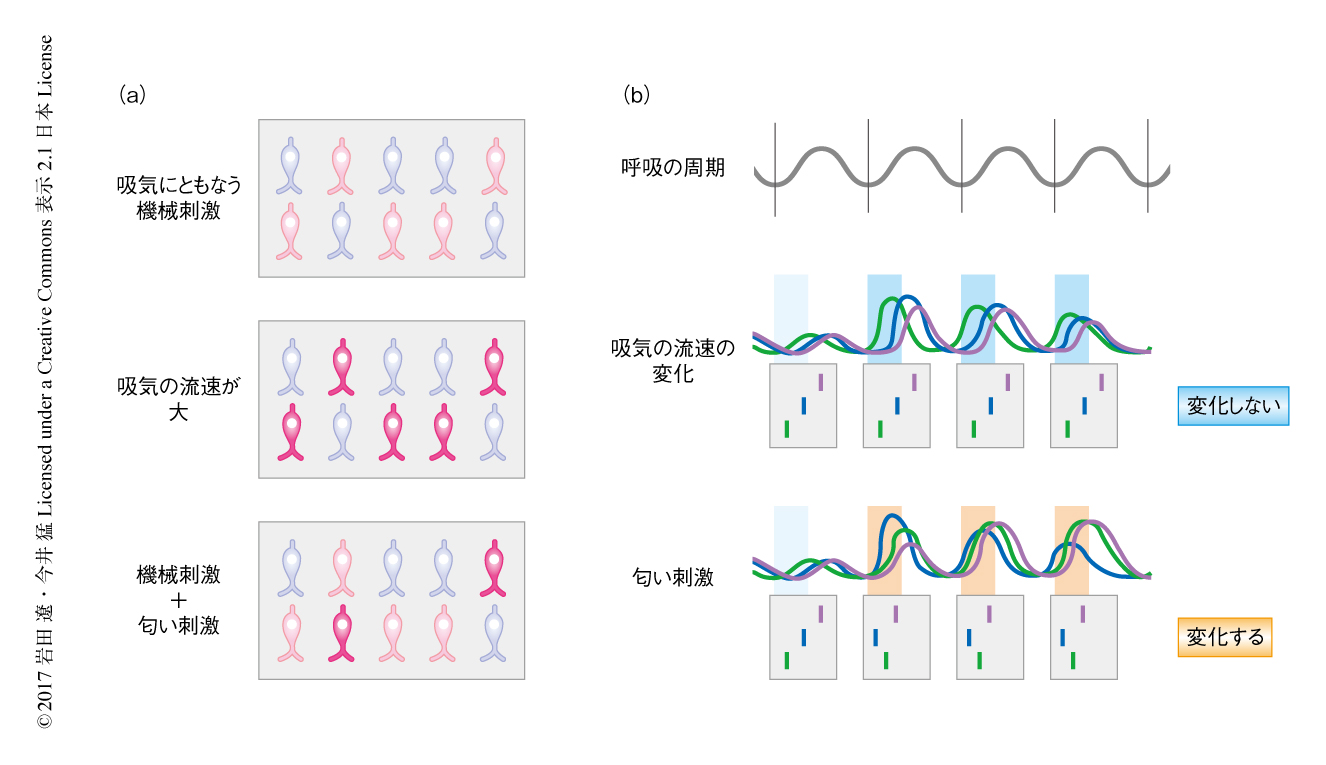
げっ歯類の嗅細胞は匂い分子だけでなく機械刺激にも応答することが知られている7).この機械刺激の受容は吸気にともなう空気の流れを検出しているという説がとなえられているが,脳において匂い刺激と機械刺激がどのように区別されるのかはよくわかっていなかった.嗅細胞により検出された機械刺激がどのように処理されるのかをさぐるため,Ca2+のプローブであるGCaMP3あるいはGCaMP6fを嗅細胞あるいは僧帽細胞に特異的に発現する遺伝子改変マウスを利用し,生体における神経活動を2光子顕微鏡により観察した.鼻腔において人工的に吸気の条件を再現したところ,嗅上皮において多くの嗅細胞が応答した(図1a).嗅球においては,嗅細胞の軸索の末端および僧帽細胞の樹状突起において,それぞれ,約5割および約8割の糸球体が有意な興奮を示した.
自律的な呼吸にともなう機械刺激に対する応答の意義をさぐる目的で,自発的に呼吸しているマウスに対し片鼻を閉塞した.鼻腔を閉塞した条件においては糸球体の自発的な発火はみられたものの特定のリズムは見い出されなかった.一方,鼻腔を開いた通常の条件においては多くの糸球体が呼吸の周期に同期した発火のパターンを示した.この呼吸の周期に同期した神経活動の振動においては,神経活動の振動の位相すなわち発火のタイミングが糸球体ごとに異なっていた.嗅球の背側の表面を広く解析したところ,神経活動の振動の位相はおおむねランダムに分布することが判明した(図1b).また,発火のタイミングは覚醒しているマウスにおいて呼吸のリズムが変化してもほぼ一定であった.こうしたことから,嗅細胞における機械刺激の受容は糸球体に固有かつ呼吸に同期した神経活動の振動を起こすのに必要だと考えられた.
行動している動物は匂いに対する感度を制御するため吸気の流速を積極的に変化させることが知られている.そのような状況においては,吸気にともなう機械刺激により多くの糸球体が応答すると考えられる.吸気がもたらす機械刺激と匂い刺激は脳においてどのように区別されるのだろうか.吸気の条件を人工的に制御しつつ,機械刺激あるいは匂い刺激に対する応答を記録しその様式の違いを解析した.吸気の流速を上昇させたところ,機械刺激の上昇にともない多くの糸球体において発火の頻度が上昇したが,この刺激に対し発火のタイミングはほとんど変化せず一定であった.吸気の流速を一定に保った状態で匂い刺激をあたえると,発火の頻度の変化だけでなく,呼吸の周期における発火のタイミングも変化した.この結果から,発火頻度コーディングにおいては機械刺激と匂い刺激の両方に対し変化がみられるのに対し,時間コーディングにおいては匂い刺激だけを抽出できることがわかった(図1b).
自然界で動物が匂いをたどって餌などの匂いの源を探す際には,匂いをくり返し嗅いで探索する必要がある.さらに,そのあいだには匂いの濃度も大きく変化するであろう.このあいだに匂いの種類に関する情報がどのようにして脳において安定的に表現されるのかは大きな謎であった.そこで,匂いをくり返し嗅ぐ状況および匂いの濃度が変化する状況において,僧帽細胞の発火の頻度および発火のタイミングがどのように変化するかをさぐった.
僧帽細胞の発火の頻度は匂いを嗅ぎつづけるとダイナミックに変化していくことが知られているが,発火のタイミングのふるまいについてはよくわかっていなかった.そこで,20回の呼吸の周期にわたり匂い刺激をあたえ,呼吸のおのおのの周期における発火の頻度および発火のタイミングを測定した.約20個の糸球体の集団について,呼吸の1回目の周期における応答のパターンを基準として20回目までのおのおのの周期における相関係数を計算し,応答のパターンの継時的な安定性を評価した.8種類の匂い分子について検討した結果,発火の頻度については相関係数が低下した一方,発火のタイミングについては相関係数が高く維持され,発火の頻度よりも安定であることが判明した.
異なる濃度の匂いを嗅がせた際の発火の頻度および発火のタイミングの安定性について,14種類の匂い分子を用いて検討した.もっとも高い濃度に対する応答のパターンを基準として,それ以外の濃度における応答のパターンの相関係数を計算し,匂いの濃度の変化に対する安定性を評価した.その結果,発火の頻度については匂いの濃度が低くなるにつれて相関係数が低下した.これに対し,発火のタイミングについては相関係数が高く維持され,発火の頻度よりも安定であることが判明した.さらに,実験で得られた発火の頻度の情報と発火のタイミングの情報のどちらを使うと匂いの濃度の違いによらずに匂いの種類を正確に判別できるか,デコーディング解析を行った.ユークリッド距離にもとづく線形分類器を用いた場合あるいはサポートベクターマシンを用いた場合のいずれにおいても,発火のタイミングの情報を用いたほうがより正確に匂いの種類を予測できることがわかった.
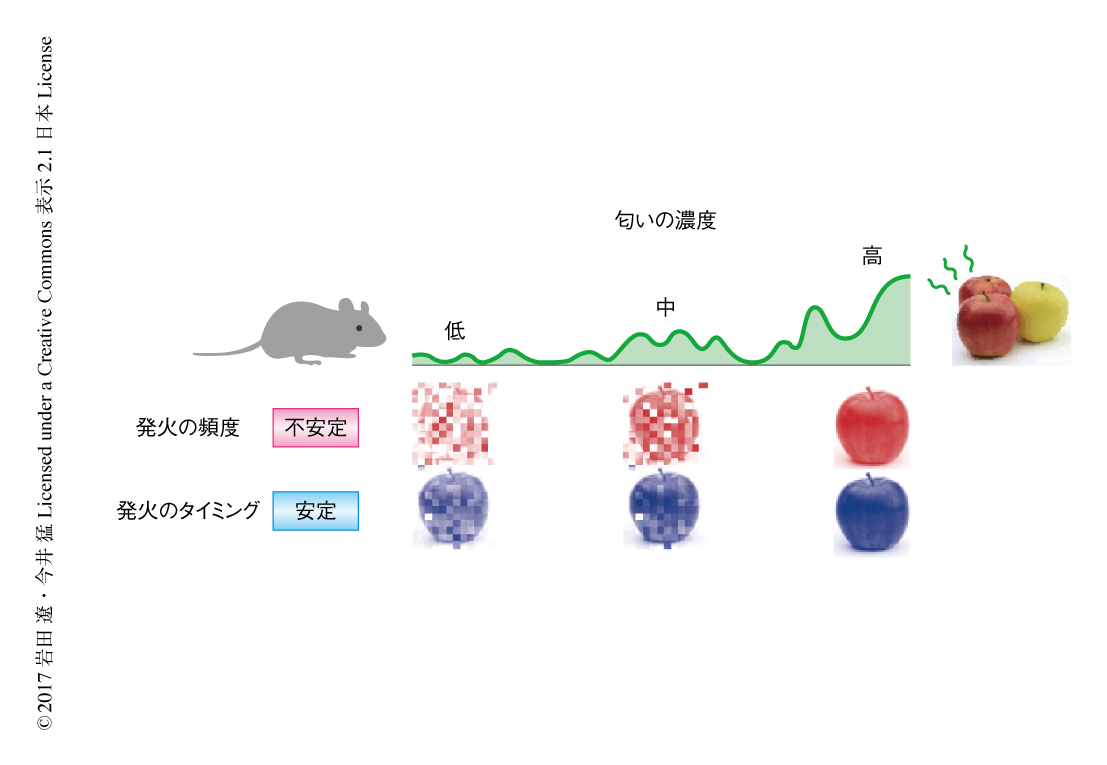
これらの結果から,僧帽細胞においては匂いの種類の情報は発火のタイミングにより表現されると考えられた.たとえば,動物が餌を探すときにはその匂いをくり返し嗅いで匂いの源を探しあてる必要がある.匂いをくり返し嗅いだり匂いの濃度が変わったりすると僧帽細胞の発火の頻度は大きく変化するが,発火のタイミングが保持されるため匂いの同一性は保たれ,匂いの探索が可能になるのではないかと考えられた(図2).
機械刺激の受容が僧帽細胞の発火のタイミングにもとづき匂い情報と識別が可能なことはわかったが,匂い情報の処理において積極的な役割を担うのだろうか.外界の匂い分子は吸気にともない周期的に鼻腔に取り込まれることから,この周期的な匂い分子の取り込みだけでも匂い情報の時間コーディングはなりたつ可能性も考えられた.この点について明らかにするためには,機械刺激がない条件において匂い刺激に対する応答を観察すればよい.しかしながら,機械刺激の受容の分子機構は匂い刺激の受容とかなり共通しており,現在のところ,機械刺激の受容のみを遺伝学的に除去するのは困難である.また,吸気を止めてしまうとそもそも匂い分子が嗅上皮に到達しないため,匂い刺激に対する応答は得られない.そこで,つぎの2つの条件を比較することにより機械刺激の受容の意義について検証した.1)匂い刺激と機械刺激の両方を周期的にあたえる条件(生理的な条件),および,2)吸気を連続的にあたえて機械刺激に由来する神経活動の振動を除去しつつ匂い刺激をパルス状にあたえる条件,である.この2つの条件において匂い刺激に対する応答を6回ずつ記録した.試行ごとに発火のタイミングを測定し,6回の測定値における標準偏差を発火のタイミングの正確性の指標とした.その結果,2)の条件においては発火のタイミングがより不正確になることが判明した.2)の条件において観察された発火のタイミングの不正確性は,匂いの濃度が低い場合とくに顕著であった.
さらに,2つの条件において得られた発火のタイミングの情報を用いて,匂いの種類をどれだけ正確に判別できるかというデコーディング解析を行った.その結果,発火のタイミングが不正確になる2)の条件においては,匂いの種類を正確に予測できなくなることが判明した.したがって,機械刺激の受容により生じた神経活動の振動は匂い情報の正確な時間コーディングを手助けする,いわば,ペースメーカーとしての役割があることが明らかにされた.
従来から,嗅球の僧帽細胞において匂い情報が発火の頻度だけでなく発火のタイミングにも表現されることは知られていたが,その意義についてはよくわかっていなかった.今回の研究において,僧帽細胞の発火のタイミングは吸気の条件が変化しても変化せず一定に保たれることが明らかにされた.さらに,匂いの濃度によらず匂いの種類の同一性が認識できるしくみは長年の難問であったが,発火のタイミングの安定性により説明されることが判明した.したがって,ノイズにみちた入力から匂いの種類に関する情報だけを抽出することが僧帽細胞における時間コーディングの役割であると考えられた.発火のタイミングを厳密に制御するための神経回路の機構の解明が今後の課題である.
神経回路における情報処理において,神経活動の振動や発火のタイミングの重要性は海馬などにおいてよく知られてきた8).ほかには,げっ歯類の体性感覚系においてもよく似た機構が報告されている.げっ歯類の体性感覚系においては,ヒゲを自ら動かすことにより生じる機械刺激と,ヒゲがなんらかの物体と接触することにより生じる機械刺激とは,発火のタイミングにもとづき区別されることが知られている9,10).したがって,感覚系における時間コーティングのひとつの役割は,能動的な感覚情報の取得にともない生じる刺激と,真に外界からもたらされる感覚情報とを区別することであるといえるかもしれない.
略歴:2011年 東京大学大学院理学系研究科博士課程 修了,同年より理化学研究所多細胞システム形成研究センター 研究員.
研究テーマ:脳における匂い情報の処理.
関心事:呼吸の周期に同期した神経活動の振動は嗅球だけではなく脳のいたるところで観察されるらしい.いったい何をしているのだろうか.
今井 猛(Takeshi Imai)
九州大学大学院医学研究院 教授.
研究室URL:http://dn.med.kyushu-u.ac.jp/
© 2017 岩田 遼・今井 猛 Licensed under CC 表示 2.1 日本
(理化学研究所多細胞システム形成研究センター 感覚神経回路形成研究チーム)
email:今井 猛
DOI: 10.7875/first.author.2017.144
Mechanosensory-based phase coding of odor identity in the olfactory bulb.
Ryo Iwata, Hiroshi Kiyonari, Takeshi Imai
Neuron, 96, 1139-1152.e7 (2017)
要 約
脳において感覚情報はニューロンの発火の頻度および発火のタイミングにより表現される.哺乳類の嗅球の僧帽細胞においては匂い情報が入力すると発火の頻度が変化するだけでなく発火のタイミングも変化する.この研究において,筆者らは,嗅細胞において受容される匂い刺激と機械刺激が僧帽細胞の発火のタイミングにもとづき区別されることを見い出した.吸気にともない嗅細胞に機械刺激が入力すると嗅球へと伝達され,嗅球の僧帽細胞において呼吸の周期に同期した神経活動の振動が生じた.神経活動の振動の位相すなわち発火のタイミングは吸気の流速を変化させても変わらなかったが,匂い刺激をあたえると変化した.匂い刺激により生じる匂いに固有の発火のタイミングは,複数の呼吸の周期にわたり匂いを嗅がせた場合においても,匂いの濃度を変化させた場合においても,発火の頻度より安定であった.また,発火のタイミングのほうが発火の頻度よりも正確に匂いの種類をコードした.さらに,機械刺激にもとづく神経活動の振動を人為的に消去したところ,匂い刺激に対する発火のタイミングは不正確になった.以上の結果から,匂いの種類に関する情報は発火の頻度よりも発火のタイミングにより安定に表現されること,発火のタイミングの正確性は嗅細胞における機械刺激の受容により保証されることが明らかにされた.
はじめに
脳における感覚情報の処理においては,外界のノイズにみちた感覚の刺激のなかから特定の情報だけが抽出される.近年,感覚情報はニューロンの発火の頻度の変化により表現されるだけではなく(発火頻度コーディング),発火のタイミングの変化によっても表現されること(時間コーディング)が明らかにされつつある1).しかしながら,感覚情報の処理における発火のタイミングの意義については不明な点が多い.
マウスの嗅覚系において,匂い分子は嗅上皮の嗅細胞により検出される.個々の嗅細胞は約1000種類ある嗅覚受容体のなかから単一の受容体だけを選択的に発現する.同一の嗅覚受容体を発現する嗅細胞は,嗅球において1対の糸球体へと軸索を収斂させる.したがって,匂い情報は嗅球にある約2000個の糸球体の時空間的な神経活動のパターンにより表現される.糸球体において,嗅細胞の軸索は投射ニューロンである僧帽細胞にくわえ,糸球体層の介在ニューロンと接続する2).僧帽細胞は嗅皮質へと軸索を投射し匂い情報を伝達する.
空気に含まれる匂い分子は,呼吸あるいは積極的な吸気にともなう空気の流れにより嗅上皮へと到達する.げっ歯類は1回の呼吸で匂いの種類を識別できるため3),そのあいだの神経活動のパターンに匂いを識別できるだけの十分な情報が含まれると考えられる.僧帽細胞においては,匂い刺激によりその発火の頻度が変化するだけでなく,匂いの種類に固有なタイミングで発火が生じる4)(図1a).さらに近年,僧帽細胞の発火のタイミングを光遺伝学的な手法により変化させるとマウスの個体が実際にそれを識別できることや5),2つの糸球体をさまざまなタイミングで光遺伝学的な手法により刺激すると嗅皮質における応答が変わることが報告された6).こうしたことから,発火のタイミングが匂い情報の処理になんらかの役割をはたすと考えられるようになった.しかし,匂い情報の処理における発火のタイミングの役割やその制御機構についてはよくわかっていなかった.
1.嗅細胞における機械刺激の受容により嗅球において呼吸の周期に同期した神経活動の振動が生じる
げっ歯類の嗅細胞は匂い分子だけでなく機械刺激にも応答することが知られている7).この機械刺激の受容は吸気にともなう空気の流れを検出しているという説がとなえられているが,脳において匂い刺激と機械刺激がどのように区別されるのかはよくわかっていなかった.嗅細胞により検出された機械刺激がどのように処理されるのかをさぐるため,Ca2+のプローブであるGCaMP3あるいはGCaMP6fを嗅細胞あるいは僧帽細胞に特異的に発現する遺伝子改変マウスを利用し,生体における神経活動を2光子顕微鏡により観察した.鼻腔において人工的に吸気の条件を再現したところ,嗅上皮において多くの嗅細胞が応答した(図1a).嗅球においては,嗅細胞の軸索の末端および僧帽細胞の樹状突起において,それぞれ,約5割および約8割の糸球体が有意な興奮を示した.
自律的な呼吸にともなう機械刺激に対する応答の意義をさぐる目的で,自発的に呼吸しているマウスに対し片鼻を閉塞した.鼻腔を閉塞した条件においては糸球体の自発的な発火はみられたものの特定のリズムは見い出されなかった.一方,鼻腔を開いた通常の条件においては多くの糸球体が呼吸の周期に同期した発火のパターンを示した.この呼吸の周期に同期した神経活動の振動においては,神経活動の振動の位相すなわち発火のタイミングが糸球体ごとに異なっていた.嗅球の背側の表面を広く解析したところ,神経活動の振動の位相はおおむねランダムに分布することが判明した(図1b).また,発火のタイミングは覚醒しているマウスにおいて呼吸のリズムが変化してもほぼ一定であった.こうしたことから,嗅細胞における機械刺激の受容は糸球体に固有かつ呼吸に同期した神経活動の振動を起こすのに必要だと考えられた.
2.僧帽細胞の発火のタイミングにより機械刺激と匂い刺激とが区別される
行動している動物は匂いに対する感度を制御するため吸気の流速を積極的に変化させることが知られている.そのような状況においては,吸気にともなう機械刺激により多くの糸球体が応答すると考えられる.吸気がもたらす機械刺激と匂い刺激は脳においてどのように区別されるのだろうか.吸気の条件を人工的に制御しつつ,機械刺激あるいは匂い刺激に対する応答を記録しその様式の違いを解析した.吸気の流速を上昇させたところ,機械刺激の上昇にともない多くの糸球体において発火の頻度が上昇したが,この刺激に対し発火のタイミングはほとんど変化せず一定であった.吸気の流速を一定に保った状態で匂い刺激をあたえると,発火の頻度の変化だけでなく,呼吸の周期における発火のタイミングも変化した.この結果から,発火頻度コーディングにおいては機械刺激と匂い刺激の両方に対し変化がみられるのに対し,時間コーディングにおいては匂い刺激だけを抽出できることがわかった(図1b).
3.匂い情報は僧帽細胞の発火のタイミングにより安定に表現される
自然界で動物が匂いをたどって餌などの匂いの源を探す際には,匂いをくり返し嗅いで探索する必要がある.さらに,そのあいだには匂いの濃度も大きく変化するであろう.このあいだに匂いの種類に関する情報がどのようにして脳において安定的に表現されるのかは大きな謎であった.そこで,匂いをくり返し嗅ぐ状況および匂いの濃度が変化する状況において,僧帽細胞の発火の頻度および発火のタイミングがどのように変化するかをさぐった.
僧帽細胞の発火の頻度は匂いを嗅ぎつづけるとダイナミックに変化していくことが知られているが,発火のタイミングのふるまいについてはよくわかっていなかった.そこで,20回の呼吸の周期にわたり匂い刺激をあたえ,呼吸のおのおのの周期における発火の頻度および発火のタイミングを測定した.約20個の糸球体の集団について,呼吸の1回目の周期における応答のパターンを基準として20回目までのおのおのの周期における相関係数を計算し,応答のパターンの継時的な安定性を評価した.8種類の匂い分子について検討した結果,発火の頻度については相関係数が低下した一方,発火のタイミングについては相関係数が高く維持され,発火の頻度よりも安定であることが判明した.
異なる濃度の匂いを嗅がせた際の発火の頻度および発火のタイミングの安定性について,14種類の匂い分子を用いて検討した.もっとも高い濃度に対する応答のパターンを基準として,それ以外の濃度における応答のパターンの相関係数を計算し,匂いの濃度の変化に対する安定性を評価した.その結果,発火の頻度については匂いの濃度が低くなるにつれて相関係数が低下した.これに対し,発火のタイミングについては相関係数が高く維持され,発火の頻度よりも安定であることが判明した.さらに,実験で得られた発火の頻度の情報と発火のタイミングの情報のどちらを使うと匂いの濃度の違いによらずに匂いの種類を正確に判別できるか,デコーディング解析を行った.ユークリッド距離にもとづく線形分類器を用いた場合あるいはサポートベクターマシンを用いた場合のいずれにおいても,発火のタイミングの情報を用いたほうがより正確に匂いの種類を予測できることがわかった.
これらの結果から,僧帽細胞においては匂いの種類の情報は発火のタイミングにより表現されると考えられた.たとえば,動物が餌を探すときにはその匂いをくり返し嗅いで匂いの源を探しあてる必要がある.匂いをくり返し嗅いだり匂いの濃度が変わったりすると僧帽細胞の発火の頻度は大きく変化するが,発火のタイミングが保持されるため匂いの同一性は保たれ,匂いの探索が可能になるのではないかと考えられた(図2).
4.機械刺激の受容にもとづく神経活動の振動は正確な時間コーディングに必要である
機械刺激の受容が僧帽細胞の発火のタイミングにもとづき匂い情報と識別が可能なことはわかったが,匂い情報の処理において積極的な役割を担うのだろうか.外界の匂い分子は吸気にともない周期的に鼻腔に取り込まれることから,この周期的な匂い分子の取り込みだけでも匂い情報の時間コーディングはなりたつ可能性も考えられた.この点について明らかにするためには,機械刺激がない条件において匂い刺激に対する応答を観察すればよい.しかしながら,機械刺激の受容の分子機構は匂い刺激の受容とかなり共通しており,現在のところ,機械刺激の受容のみを遺伝学的に除去するのは困難である.また,吸気を止めてしまうとそもそも匂い分子が嗅上皮に到達しないため,匂い刺激に対する応答は得られない.そこで,つぎの2つの条件を比較することにより機械刺激の受容の意義について検証した.1)匂い刺激と機械刺激の両方を周期的にあたえる条件(生理的な条件),および,2)吸気を連続的にあたえて機械刺激に由来する神経活動の振動を除去しつつ匂い刺激をパルス状にあたえる条件,である.この2つの条件において匂い刺激に対する応答を6回ずつ記録した.試行ごとに発火のタイミングを測定し,6回の測定値における標準偏差を発火のタイミングの正確性の指標とした.その結果,2)の条件においては発火のタイミングがより不正確になることが判明した.2)の条件において観察された発火のタイミングの不正確性は,匂いの濃度が低い場合とくに顕著であった.
さらに,2つの条件において得られた発火のタイミングの情報を用いて,匂いの種類をどれだけ正確に判別できるかというデコーディング解析を行った.その結果,発火のタイミングが不正確になる2)の条件においては,匂いの種類を正確に予測できなくなることが判明した.したがって,機械刺激の受容により生じた神経活動の振動は匂い情報の正確な時間コーディングを手助けする,いわば,ペースメーカーとしての役割があることが明らかにされた.
おわりに
従来から,嗅球の僧帽細胞において匂い情報が発火の頻度だけでなく発火のタイミングにも表現されることは知られていたが,その意義についてはよくわかっていなかった.今回の研究において,僧帽細胞の発火のタイミングは吸気の条件が変化しても変化せず一定に保たれることが明らかにされた.さらに,匂いの濃度によらず匂いの種類の同一性が認識できるしくみは長年の難問であったが,発火のタイミングの安定性により説明されることが判明した.したがって,ノイズにみちた入力から匂いの種類に関する情報だけを抽出することが僧帽細胞における時間コーディングの役割であると考えられた.発火のタイミングを厳密に制御するための神経回路の機構の解明が今後の課題である.
神経回路における情報処理において,神経活動の振動や発火のタイミングの重要性は海馬などにおいてよく知られてきた8).ほかには,げっ歯類の体性感覚系においてもよく似た機構が報告されている.げっ歯類の体性感覚系においては,ヒゲを自ら動かすことにより生じる機械刺激と,ヒゲがなんらかの物体と接触することにより生じる機械刺激とは,発火のタイミングにもとづき区別されることが知られている9,10).したがって,感覚系における時間コーティングのひとつの役割は,能動的な感覚情報の取得にともない生じる刺激と,真に外界からもたらされる感覚情報とを区別することであるといえるかもしれない.
文 献
- Buzsaki, G. & Draguhn, A.: Neuronal oscillations in cortical networks. Science, 304, 1926-1929 (2004)[PubMed]
- Imai, T.: Construction of functional neuronal circuitry in the olfactory bulb. Semin. Cell Dev. Biol., 35, 180-188 (2014)[PubMed]
- Uchida, N. & Mainen, Z. F.: Speed and accuracy of olfactory discrimination in the rat. Nat. Neurosci., 6, 1224-1229 (2003)[PubMed]
- Uchida, N., Poo, C. & Haddad, R.: Coding and transformations in the olfactory system. Annu. Rev. Neurosci., 37, 363-385 (2014)[PubMed]
- Smear, M., Resulaj, A., Zhang, J. et al.: Multiple perceptible signals from a single olfactory glomerulus. Nat. Neurosci., 16, 1687-1691 (2013)[PubMed]
- Haddad, R., Lanjuin, A., Madisen, L. et al.: Olfactory cortical neurons read out a relative time code in the olfactory bulb. Nat. Neurosci., 949-957 (2013)[PubMed]
- Grosmaitre, X., Santarelli, L. C., Tan, J. et al.: Dual functions of mammalian olfactory sensory neurons as odor detectors and mechanical sensors. Nat. Neurosci., 10, 348-354 (2007)[PubMed]
- Buzsaki, G. & Moser, E. I.: Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci., 16, 130-138 (2013)[PubMed]
- Kleinfeld, D. & Deschenes, M.: Neuronal basis for object location in the vibrissa scanning sensorimotor system. Neuron, 72, 455-468 (2011)[PubMed]
- Severson, K. S., Xu, D., Van de Loo, M. et al.: Active touch and self-motion encoding by Merkel cell-associated afferents. Neuron, 94, 666-676.e9 (2017)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2011年 東京大学大学院理学系研究科博士課程 修了,同年より理化学研究所多細胞システム形成研究センター 研究員.
研究テーマ:脳における匂い情報の処理.
関心事:呼吸の周期に同期した神経活動の振動は嗅球だけではなく脳のいたるところで観察されるらしい.いったい何をしているのだろうか.
今井 猛(Takeshi Imai)
九州大学大学院医学研究院 教授.
研究室URL:http://dn.med.kyushu-u.ac.jp/
© 2017 岩田 遼・今井 猛 Licensed under CC 表示 2.1 日本
