神経幹細胞はトポロジカル欠陥に集積する
川口喬吾1・影山龍一郎2・佐野雅己3
(1米国Harvard Medical School,Department of Systems Biology,2京都大学ウイルス・再生医科学研究所 増殖制御システム分野,3東京大学理学系研究科物理学専攻)
email:川口喬吾
DOI: 10.7875/first.author.2017.040
Topological defects control collective dynamics in neural progenitor cell cultures.
Kyogo Kawaguchi, Ryoichiro Kageyama, Masaki Sano
Nature, 545, 327-331 (2017)
培養皿において増殖する神経幹細胞は生体と同様に棒状のかたちをしており,高密度になると,となりどうしの細胞が向きをそろえる.この研究においては,神経幹細胞が向きをそろえあうことにより生じるパターンがネマチック液晶などにみられるパターンと同じであることに着目し,数万のスケールの神経幹細胞を長時間にわたり観察した.その結果,神経幹細胞は既存の液晶とは違いパターンのなかで激しく自発運動しており,局所的に細胞の集団の流れが生じるばかりでなく,細胞が集積しやすい場所や離散しやすい場所が現われることがわかった.これらの現象は,トポロジカル欠陥とよばれる細胞の配列の特異点の位置により予測でき,液晶におけるアクティブマターの理論の枠組みにより自然な説明することができた.このように,細胞の集団がそれ自体でつくる配列のパターンに誘起されて細胞流が生じる現象ははじめてみつかったもので,成体の脳における新生したニューロンの移動をはじめとして,生体における細胞の移動の機構にかかわる可能性がある.
ヒトを含む多細胞生物においては,発生の過程だけでなく成体においても細胞は新生し移動しつづけている.にもかかわらず,個体のあいだで器官の形状や機能に大きな差はなく,一生のうちほとんどは恒常性が維持されている.このような多細胞のレベルでの動的な組織の形成および恒常性の維持の機構を理解するにあたり,多体系の物理学の手法は有効であると思われる.多体系の物理学(あるいは,統計物理学)とは,複雑すぎてそのままでは手におえない多体問題から本質的な部分をぬきだし,要素の詳細によらない一般的な枠組みをつくることをめざす試みである.多体系の物理学は,半導体や超伝導体を含む固体や対流現象や乱流現象を生む流体だけでなく,液晶やゲルなどのソフトな物質の理解および制御にも役だってきた.しかし,生体の組織においては,細胞が運動や増殖をしていたり,いっけん静止している場合にも細胞骨格に依存した内在的な力がはたらいていたりするため,典型的な多体系の物理学の適用の範囲を逸脱してしまう場合が多く,昔から,単純な応用問題ではないことが知られていた.そこで近年では,既存の多体系の物理学の理論をどのように拡張すれば細胞の集団の運動や生物の群れの運動が説明できるか,また,拡張によりこれまでにない新しい現象の予測ができるか,といった方向でさかんに研究が進められており1),広く“アクティブマター”とよばれる学際的な研究領域が形成されている.
アクティブ(能動的,つまり,構成要素が自発運動する)であることにより,既存のパッシブ(受動的)な系とどのように違った現象が現われるのか.1990年代の理論研究により明らかにされたのは2),パッシブな系では不可能な2次元の長距離秩序(マクロな距離スケールにわたり粒子が同じ向きにそろった状態)がアクティブな系では自発的に達成されうることや,アクティブな系では秩序構造のなかでの粒子数のゆらぎが異常に大きくなることである.これらの予測は,棒状の物質をしきつめた系に振動をあたえる実験や細菌の集団運動において検証されてきた(理論からの予測に忠実な実験系における検証は,じつは,ごく最近の研究3) までなかった).1990年代の後半から2000年代にかけて,細胞骨格の示す自己組織的な性質が実験的および理論的に解明されはじめたほか4),培養皿において線維芽細胞の形成するパターンを調べた研究なども現われ5),いよいよ,アクティブマターの理論の生命の理解への本格的な応用が進むかにみえた.
ところが,2010年代に入ると,アクティブマターの枠組みにおける新現象の発見および実験系の精緻化の方向への発展がめだった.とくに,細胞から取り出した微小管やアクチンの高濃度での2次元パターンを観察した研究は6-8),息をのむほど美しい集団の運動の動画を提供したが,実際の細胞において起こっていることからはかけ離れた現象であることは明らかだった.細胞分裂などの実際の生命現象にアクティブマターの理論を適用する意欲的な研究も現われはじめたが9,10),多細胞生物の現象にアクティブマターの理論を拡張する研究は,最近になりようやく再始動した感がある.
神経幹細胞はニューロン,オリゴデンドロサイト,アストロサイトへの分化能をもつ幹細胞であり,おもにマウスの胎仔や成体から取り出し2次元の培養皿において長期間の培養が可能な簡便な培養細胞系である.細胞の形態は生体と同様に細長く,増殖して培養皿の2次元平面を埋めつくすほど高密度になると,細胞どうしが向きをそろえあう状況がみられる.
神経幹細胞の向きのそろう現象がどのように生じるのかをみるため,顕微鏡により長時間にわたり撮影した.その結果,神経幹細胞は低密度においては方向性のないランダムな運動を示すものの,高密度になるにしたがい細胞どうしがぶつかりあい,形状だけでなく運動の方向もしだいにそろっていくことがわかった.ここで配列の方向とは,頭と尻尾とを区別しない方向のことである.ヒストンH2Bと蛍光タンパク質mCherryとの融合タンパク質を発現する神経幹細胞株を作製し核の運動を追跡した結果,細胞の運動は高密度になってもまったく減速しない代わりに,パターンを形成したのちには配列の方向に沿うようになり,進行の方向を反転しうる双極的なものに変化することもわかった.
高密度において発生したパターンを数mmにわたる大きな領域において観察したところ,ネマチック液晶とよばれる物質が2次元において形成するパターンとよく似たものがみられた(図1a).ネマチック液晶を薄いセルのなかに閉じ込め偏光フィルターを用いて観察すると,シュリーレン模様とよばれる光学的なパターンがみられる(図1b).このパターンは棒状の粒子である液晶分子が場所により違った向きに配列することに起因しており,分子のそろう向きに依存して光のとおしやすさが違うことから明暗の模様が生じる.画像解析により神経幹細胞の配列の方向を色分けして可視化するとシュリーレン模様と同様なパターンがみられ,局所的にはある色が選ばれ,少し離れた箇所では違う色に変化しており,それが連続的に移り変わっているようすがみられた(図1a).

培養皿を神経幹細胞が自由に埋めつくす場合には,場所によりランダムに定まった配列の方向が生じるため,細胞は空間的に離れた点では違う向きにそろっている.液晶ディスプレイに使われる素子などでは,液晶を閉じ込めるセルの表面に適切な処理を施すことにより液晶分子が自発的に特定の向きにそろうようあらかじめ制御する.神経幹細胞の場合にも同様に,細胞の集団に対し境界条件を設置することにより配列の方向をそろえられないだろうか.
神経幹細胞が培養皿の表面につけた傷のなかに侵入できないことを利用して,2つの直線状の傷を短い間隔でつけ,そのあいだに閉じ込められた細胞の運動を観察した.傷が境界となり神経幹細胞はこの境界に沿って配列したが,その配列の方向は傷から離れた細胞にも伝達していき,結果的に,細長い環境において細胞をすべて同じ向きに自発的に配列させることができた.このようなパターンは数mmの長さにわたり継続することが示され,境界を用意するかぎりいくらでも長いパターンを形成できると考えられた.
細長い環境に閉じ込められた神経幹細胞は,さきと同様に,細胞の伸長方向への速い運動や双極的な進行方向の切り替えを示した.このようすは,成体の脳において新生したニューロンが嗅球に移動する際にみられる細長い流路(rostral migratory stream)における運動とよく似ていた11).この細長い流路において重要な細胞接着因子であるNCAMを神経幹細胞においてノックダウンしたところ,細胞間の相互作用が影響をうけ,高密度において神経幹細胞が向きをそろえあう現象そのものが破壊された.成体の脳における移動流にかかわる神経芽細胞は神経幹細胞よりも分化の進んだ細胞であるが,形状や移動にかかわる細胞間の相互作用は似ていると推測される.一般的なアクティブマターの理論から,自発的な細胞の運動がランダムあるいは頭と尻尾の方向に対称であっても,配列のパターンや境界条件の設定により集団的な流れを一方向に整流させられることがわかるが,生体における細胞の移動においても,このように誘引物質にたよらない集団の運動の機構がはたらく可能性がある.
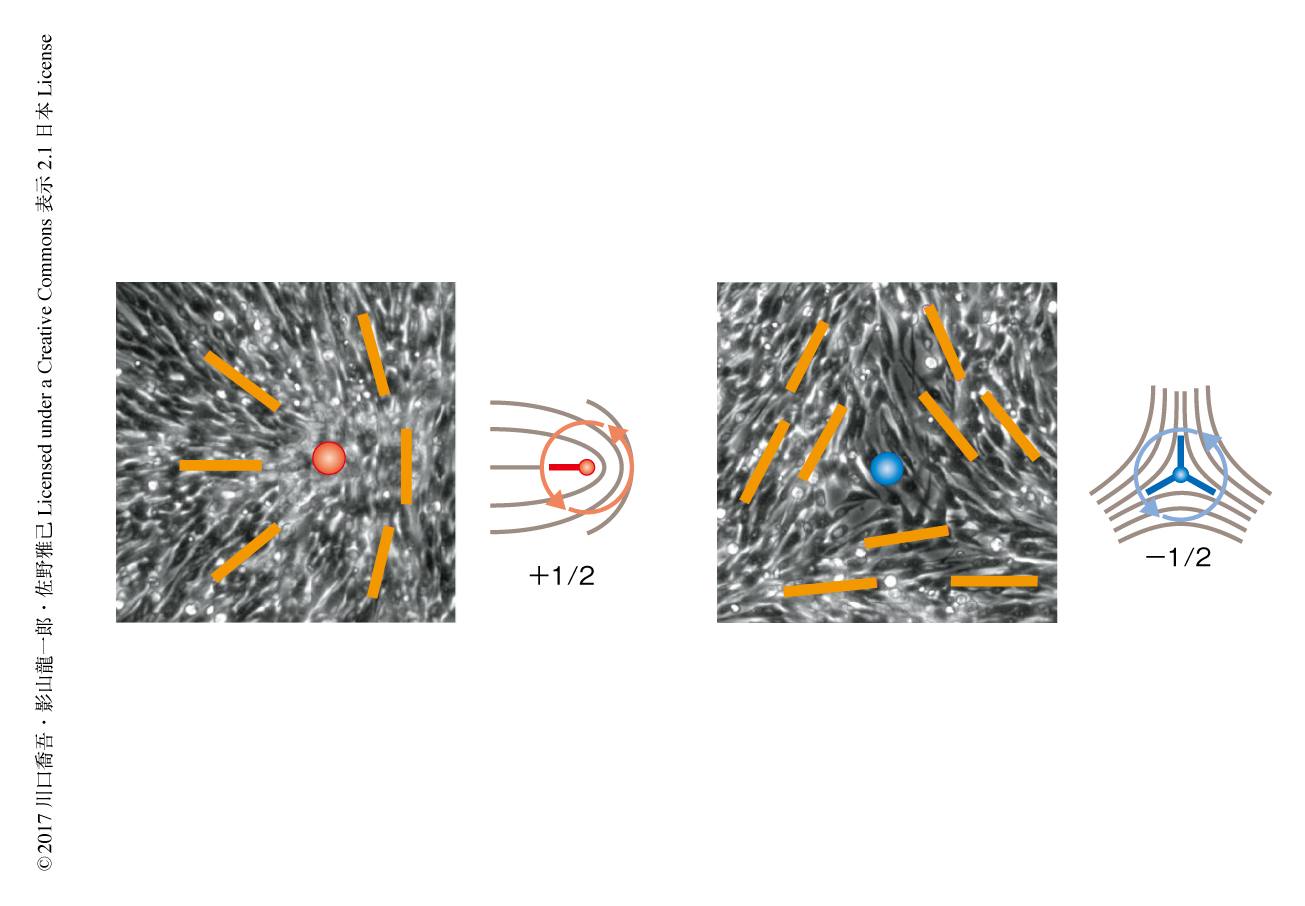
神経幹細胞の集団の配列の方向を可視化すると,色が滑らかに変化していない特異点がいくつも存在するのがみえる.これは一般にトポロジカル欠陥とよばれる構造で,細胞がうまく向きをそろえられていない箇所でありながら,周囲を配列のそろった状態にかこまれて“保護”されるため容易に解消されない安定な構造である.観察されたトポロジカル欠陥は+1/2欠陥と-1/2欠陥の2種類で,特異点のまわりを左回りに1周したとき,細胞の向きが左回りに1/2回転するものが+1/2欠陥,右回りに1/2回転するものが-1/2欠陥である(図2).このように,半整数のトポロジカル欠陥が現われるのはネマチック液晶と同様であり,細胞間の相互作用が頭と尻尾を区別しないもの(これを,ネマチックとよぶ)であることに起因する.
さらに,トポロジカル欠陥の付近における神経幹細胞の集団の挙動を長時間にわたり観察したところ,+1/2欠陥には細胞が集積することがわかった.細胞が+1/2欠陥に流れ込むことにより細胞の局所的な密度が上昇し,最終的には3次元的に盛りあがったマウンドが形成された.より詳細な画像解析により,神経幹細胞は+1/2欠陥に集積するだけでなく,-1/2欠陥から平均として離散することも確認された.
最近の微小管を用いた実験により6),アクティブな系においてトポロジカル欠陥そのものが遊走する現象がみつかったことにより,トポロジカル欠陥のふるまいがアクティブな系とパッシブな系とで明確に違うことが認識され注目をあつめていた.ところが,半整数のトポロジカル欠陥において要素が集積するような現象は,これまで,理論や数値シミュレーションにおいても知られていなかった.
トポロジカル欠陥は細胞の集団それ自体が形成するパターンの帰結であるが,集団のパターンが細胞の挙動を決めるのはどのような機構によるのか.トポロジカル欠陥のようにパターンのなかに非対称性がある場所では,個々の細胞の運動が積み重なってマクロな集団の運動が生じうる.これが微小管を用いた実験などでみられたトポロジカル欠陥の遊走の原因であるが,この集団の運動が細胞の配列の方向と垂直な場合には運動を押しとどめようとする“摩擦”の効果がはたらき,せき止められた細胞は集積するはずである.実際に,この直観的な描像から得られるもっとも単純な式を解くことにより,トポロジカル欠陥の付近において要素が集積あるいは離散する現象を説明できることが示された.この理論のカギは,アクティブな要素と配列に依存した摩擦の効果の組合せを考えることにあるが,同じ条件さえそろえば,神経幹細胞のほかでも同様に集積および離散の現象が現われうることが示唆された.
この研究は,筆者のひとり(川口)が神経幹細胞を用いた実験を習っていたところ思いがけずおもしろい現象をみつけたため,それを解析しまとめたものである.神経幹細胞の培養はほかの細胞に比べ少しだけ厄介な点や,トポロジカル欠陥の自動検出や細胞の追跡など画像解析の手法の開発がやや面倒であった点を除けば,標準的な実験設備で可能な素朴な研究であるといえる.たとえば,細胞を細長い環境に閉じ込める実験は,培養皿を文房具のカッターを用いて手書きで傷をつけて観察しただけのもので,正攻法であるマイクロパターニングなどと比べかなり初等的な方法である.近年,生体試料の観察の技術はめまぐるしく発展しているが,最先端の技術によりはじめてとらえられる観察の困難な現象だけでなく,いまどきの実験室では日常化してしまった技術により観察できる現象のなかにも,非自明なものが多く存在しているように思う.
この論文と同時に同じNature誌に出版された論文12) において,MDCK細胞の表皮シートにおけるトポロジカル欠陥が調べられ,トポロジカル欠陥の付近では細胞が押し出されやすいという現象が報告された.MDCK細胞と神経幹細胞とでは運動の様式などが大きく異なるものの,アクティブな系ゆえにトポロジカル欠陥に細胞が集積するという広い意味では同様の現象であるといえる.表皮細胞の押し出しにせよ神経幹細胞の移動流にせよ,生体において実際に起こっていることとどの程度の関連があるのかは未知数であるが,多体系の物理学を拡張して多細胞生物の現象の研究にもちこみ生体組織のダイナミクスの理解に役だてようという機運は高まっており,これらの研究がその第一歩となるものと期待される.
略歴:2015年 東京大学大学院理学系研究科物理学専攻博士課程 修了,同年より米国Harvard Medical School博士研究員.
研究テーマ:生物物理の理論と実験.
影山 龍一郎(Ryoichiro Kageyama)
京都大学ウイルス研究所 教授.
佐野 雅己(Masaki Sano)
東京大学理学系研究科物理学専攻 教授.
© 2017 川口喬吾・影山龍一郎・佐野雅己 Licensed under CC 表示 2.1 日本
(1米国Harvard Medical School,Department of Systems Biology,2京都大学ウイルス・再生医科学研究所 増殖制御システム分野,3東京大学理学系研究科物理学専攻)
email:川口喬吾
DOI: 10.7875/first.author.2017.040
Topological defects control collective dynamics in neural progenitor cell cultures.
Kyogo Kawaguchi, Ryoichiro Kageyama, Masaki Sano
Nature, 545, 327-331 (2017)
要 約
培養皿において増殖する神経幹細胞は生体と同様に棒状のかたちをしており,高密度になると,となりどうしの細胞が向きをそろえる.この研究においては,神経幹細胞が向きをそろえあうことにより生じるパターンがネマチック液晶などにみられるパターンと同じであることに着目し,数万のスケールの神経幹細胞を長時間にわたり観察した.その結果,神経幹細胞は既存の液晶とは違いパターンのなかで激しく自発運動しており,局所的に細胞の集団の流れが生じるばかりでなく,細胞が集積しやすい場所や離散しやすい場所が現われることがわかった.これらの現象は,トポロジカル欠陥とよばれる細胞の配列の特異点の位置により予測でき,液晶におけるアクティブマターの理論の枠組みにより自然な説明することができた.このように,細胞の集団がそれ自体でつくる配列のパターンに誘起されて細胞流が生じる現象ははじめてみつかったもので,成体の脳における新生したニューロンの移動をはじめとして,生体における細胞の移動の機構にかかわる可能性がある.
はじめに
ヒトを含む多細胞生物においては,発生の過程だけでなく成体においても細胞は新生し移動しつづけている.にもかかわらず,個体のあいだで器官の形状や機能に大きな差はなく,一生のうちほとんどは恒常性が維持されている.このような多細胞のレベルでの動的な組織の形成および恒常性の維持の機構を理解するにあたり,多体系の物理学の手法は有効であると思われる.多体系の物理学(あるいは,統計物理学)とは,複雑すぎてそのままでは手におえない多体問題から本質的な部分をぬきだし,要素の詳細によらない一般的な枠組みをつくることをめざす試みである.多体系の物理学は,半導体や超伝導体を含む固体や対流現象や乱流現象を生む流体だけでなく,液晶やゲルなどのソフトな物質の理解および制御にも役だってきた.しかし,生体の組織においては,細胞が運動や増殖をしていたり,いっけん静止している場合にも細胞骨格に依存した内在的な力がはたらいていたりするため,典型的な多体系の物理学の適用の範囲を逸脱してしまう場合が多く,昔から,単純な応用問題ではないことが知られていた.そこで近年では,既存の多体系の物理学の理論をどのように拡張すれば細胞の集団の運動や生物の群れの運動が説明できるか,また,拡張によりこれまでにない新しい現象の予測ができるか,といった方向でさかんに研究が進められており1),広く“アクティブマター”とよばれる学際的な研究領域が形成されている.
アクティブ(能動的,つまり,構成要素が自発運動する)であることにより,既存のパッシブ(受動的)な系とどのように違った現象が現われるのか.1990年代の理論研究により明らかにされたのは2),パッシブな系では不可能な2次元の長距離秩序(マクロな距離スケールにわたり粒子が同じ向きにそろった状態)がアクティブな系では自発的に達成されうることや,アクティブな系では秩序構造のなかでの粒子数のゆらぎが異常に大きくなることである.これらの予測は,棒状の物質をしきつめた系に振動をあたえる実験や細菌の集団運動において検証されてきた(理論からの予測に忠実な実験系における検証は,じつは,ごく最近の研究3) までなかった).1990年代の後半から2000年代にかけて,細胞骨格の示す自己組織的な性質が実験的および理論的に解明されはじめたほか4),培養皿において線維芽細胞の形成するパターンを調べた研究なども現われ5),いよいよ,アクティブマターの理論の生命の理解への本格的な応用が進むかにみえた.
ところが,2010年代に入ると,アクティブマターの枠組みにおける新現象の発見および実験系の精緻化の方向への発展がめだった.とくに,細胞から取り出した微小管やアクチンの高濃度での2次元パターンを観察した研究は6-8),息をのむほど美しい集団の運動の動画を提供したが,実際の細胞において起こっていることからはかけ離れた現象であることは明らかだった.細胞分裂などの実際の生命現象にアクティブマターの理論を適用する意欲的な研究も現われはじめたが9,10),多細胞生物の現象にアクティブマターの理論を拡張する研究は,最近になりようやく再始動した感がある.
1.神経幹細胞は高密度になると向きをそろえあいネマチック液晶と同じパターンを形成する
神経幹細胞はニューロン,オリゴデンドロサイト,アストロサイトへの分化能をもつ幹細胞であり,おもにマウスの胎仔や成体から取り出し2次元の培養皿において長期間の培養が可能な簡便な培養細胞系である.細胞の形態は生体と同様に細長く,増殖して培養皿の2次元平面を埋めつくすほど高密度になると,細胞どうしが向きをそろえあう状況がみられる.
神経幹細胞の向きのそろう現象がどのように生じるのかをみるため,顕微鏡により長時間にわたり撮影した.その結果,神経幹細胞は低密度においては方向性のないランダムな運動を示すものの,高密度になるにしたがい細胞どうしがぶつかりあい,形状だけでなく運動の方向もしだいにそろっていくことがわかった.ここで配列の方向とは,頭と尻尾とを区別しない方向のことである.ヒストンH2Bと蛍光タンパク質mCherryとの融合タンパク質を発現する神経幹細胞株を作製し核の運動を追跡した結果,細胞の運動は高密度になってもまったく減速しない代わりに,パターンを形成したのちには配列の方向に沿うようになり,進行の方向を反転しうる双極的なものに変化することもわかった.
高密度において発生したパターンを数mmにわたる大きな領域において観察したところ,ネマチック液晶とよばれる物質が2次元において形成するパターンとよく似たものがみられた(図1a).ネマチック液晶を薄いセルのなかに閉じ込め偏光フィルターを用いて観察すると,シュリーレン模様とよばれる光学的なパターンがみられる(図1b).このパターンは棒状の粒子である液晶分子が場所により違った向きに配列することに起因しており,分子のそろう向きに依存して光のとおしやすさが違うことから明暗の模様が生じる.画像解析により神経幹細胞の配列の方向を色分けして可視化するとシュリーレン模様と同様なパターンがみられ,局所的にはある色が選ばれ,少し離れた箇所では違う色に変化しており,それが連続的に移り変わっているようすがみられた(図1a).
2.細長い環境に閉じ込められた神経幹細胞は長距離にわたり配列する
培養皿を神経幹細胞が自由に埋めつくす場合には,場所によりランダムに定まった配列の方向が生じるため,細胞は空間的に離れた点では違う向きにそろっている.液晶ディスプレイに使われる素子などでは,液晶を閉じ込めるセルの表面に適切な処理を施すことにより液晶分子が自発的に特定の向きにそろうようあらかじめ制御する.神経幹細胞の場合にも同様に,細胞の集団に対し境界条件を設置することにより配列の方向をそろえられないだろうか.
神経幹細胞が培養皿の表面につけた傷のなかに侵入できないことを利用して,2つの直線状の傷を短い間隔でつけ,そのあいだに閉じ込められた細胞の運動を観察した.傷が境界となり神経幹細胞はこの境界に沿って配列したが,その配列の方向は傷から離れた細胞にも伝達していき,結果的に,細長い環境において細胞をすべて同じ向きに自発的に配列させることができた.このようなパターンは数mmの長さにわたり継続することが示され,境界を用意するかぎりいくらでも長いパターンを形成できると考えられた.
細長い環境に閉じ込められた神経幹細胞は,さきと同様に,細胞の伸長方向への速い運動や双極的な進行方向の切り替えを示した.このようすは,成体の脳において新生したニューロンが嗅球に移動する際にみられる細長い流路(rostral migratory stream)における運動とよく似ていた11).この細長い流路において重要な細胞接着因子であるNCAMを神経幹細胞においてノックダウンしたところ,細胞間の相互作用が影響をうけ,高密度において神経幹細胞が向きをそろえあう現象そのものが破壊された.成体の脳における移動流にかかわる神経芽細胞は神経幹細胞よりも分化の進んだ細胞であるが,形状や移動にかかわる細胞間の相互作用は似ていると推測される.一般的なアクティブマターの理論から,自発的な細胞の運動がランダムあるいは頭と尻尾の方向に対称であっても,配列のパターンや境界条件の設定により集団的な流れを一方向に整流させられることがわかるが,生体における細胞の移動においても,このように誘引物質にたよらない集団の運動の機構がはたらく可能性がある.
3.神経幹細胞の集団はトポロジカル欠陥のうち+1/2欠陥に集積し-1/2欠陥から離散する
神経幹細胞の集団の配列の方向を可視化すると,色が滑らかに変化していない特異点がいくつも存在するのがみえる.これは一般にトポロジカル欠陥とよばれる構造で,細胞がうまく向きをそろえられていない箇所でありながら,周囲を配列のそろった状態にかこまれて“保護”されるため容易に解消されない安定な構造である.観察されたトポロジカル欠陥は+1/2欠陥と-1/2欠陥の2種類で,特異点のまわりを左回りに1周したとき,細胞の向きが左回りに1/2回転するものが+1/2欠陥,右回りに1/2回転するものが-1/2欠陥である(図2).このように,半整数のトポロジカル欠陥が現われるのはネマチック液晶と同様であり,細胞間の相互作用が頭と尻尾を区別しないもの(これを,ネマチックとよぶ)であることに起因する.
さらに,トポロジカル欠陥の付近における神経幹細胞の集団の挙動を長時間にわたり観察したところ,+1/2欠陥には細胞が集積することがわかった.細胞が+1/2欠陥に流れ込むことにより細胞の局所的な密度が上昇し,最終的には3次元的に盛りあがったマウンドが形成された.より詳細な画像解析により,神経幹細胞は+1/2欠陥に集積するだけでなく,-1/2欠陥から平均として離散することも確認された.
最近の微小管を用いた実験により6),アクティブな系においてトポロジカル欠陥そのものが遊走する現象がみつかったことにより,トポロジカル欠陥のふるまいがアクティブな系とパッシブな系とで明確に違うことが認識され注目をあつめていた.ところが,半整数のトポロジカル欠陥において要素が集積するような現象は,これまで,理論や数値シミュレーションにおいても知られていなかった.
トポロジカル欠陥は細胞の集団それ自体が形成するパターンの帰結であるが,集団のパターンが細胞の挙動を決めるのはどのような機構によるのか.トポロジカル欠陥のようにパターンのなかに非対称性がある場所では,個々の細胞の運動が積み重なってマクロな集団の運動が生じうる.これが微小管を用いた実験などでみられたトポロジカル欠陥の遊走の原因であるが,この集団の運動が細胞の配列の方向と垂直な場合には運動を押しとどめようとする“摩擦”の効果がはたらき,せき止められた細胞は集積するはずである.実際に,この直観的な描像から得られるもっとも単純な式を解くことにより,トポロジカル欠陥の付近において要素が集積あるいは離散する現象を説明できることが示された.この理論のカギは,アクティブな要素と配列に依存した摩擦の効果の組合せを考えることにあるが,同じ条件さえそろえば,神経幹細胞のほかでも同様に集積および離散の現象が現われうることが示唆された.
おわりに
この研究は,筆者のひとり(川口)が神経幹細胞を用いた実験を習っていたところ思いがけずおもしろい現象をみつけたため,それを解析しまとめたものである.神経幹細胞の培養はほかの細胞に比べ少しだけ厄介な点や,トポロジカル欠陥の自動検出や細胞の追跡など画像解析の手法の開発がやや面倒であった点を除けば,標準的な実験設備で可能な素朴な研究であるといえる.たとえば,細胞を細長い環境に閉じ込める実験は,培養皿を文房具のカッターを用いて手書きで傷をつけて観察しただけのもので,正攻法であるマイクロパターニングなどと比べかなり初等的な方法である.近年,生体試料の観察の技術はめまぐるしく発展しているが,最先端の技術によりはじめてとらえられる観察の困難な現象だけでなく,いまどきの実験室では日常化してしまった技術により観察できる現象のなかにも,非自明なものが多く存在しているように思う.
この論文と同時に同じNature誌に出版された論文12) において,MDCK細胞の表皮シートにおけるトポロジカル欠陥が調べられ,トポロジカル欠陥の付近では細胞が押し出されやすいという現象が報告された.MDCK細胞と神経幹細胞とでは運動の様式などが大きく異なるものの,アクティブな系ゆえにトポロジカル欠陥に細胞が集積するという広い意味では同様の現象であるといえる.表皮細胞の押し出しにせよ神経幹細胞の移動流にせよ,生体において実際に起こっていることとどの程度の関連があるのかは未知数であるが,多体系の物理学を拡張して多細胞生物の現象の研究にもちこみ生体組織のダイナミクスの理解に役だてようという機運は高まっており,これらの研究がその第一歩となるものと期待される.
文 献
- Marchetti, M. C., Joanny, J. -F., Ramaswamy, S. et al.: Hydrodynamics of soft active matter. Rev. Mod. Phys., 85, 1143 (2013)
- Vicsek, T. & Zafeiris, A.: Collective motion. Phys. Rep., 517, 71-140 (2012)
- Nishiguchi, D., Nagai, K. H., Chate, H. et al.: Long-range nematic order and anomalous fluctuations in suspensions of swimming filamentous bacteria. Phys. Rev. E, 95, 020601 (2017)[PubMed]
- Nedelec, F. J., Surrey, T., Maggs, A. C. et al.: Self-organization of microtubules and motors. Nature, 389, 305-308 (1997)[PubMed]
- Kemkemer, R., Teichgraber, V., Schrank-Kaufmann, S. et al.: Nematic order-disorder state transition in a liquid crystal analogue formed by oriented and migrating amoeboid cells. Eur. Phys. J. E, 3, 101 (2000)
- Sanchez, T., Chen, D. T., DeCamp, S. J. et al.: Spontaneous motion in hierarchically assembled active matter. Nature, 491, 431-434 (2012)[PubMed]
- Schaller, V., Weber, C., Semmrich, C. et al.: Polar patterns of driven filaments. Nature, 467, 73-77 (2010)[PubMed]
- Sumino, Y., Nagai, K., Shitaka, Y. et al.: Large-scale vortex lattice emerging from collectively moving microtubules. Nature, 483, 448-452 (2012)[PubMed]
- Brugues, J. & Needleman, D.: Physical basis of spindle self-organization. Proc. Natl. Acad. Sci. USA, 111, 18496-18500 (2014)[PubMed]
- Prost, J., Julicher, F. & Joanny, J. -F.: Active gel physics. Nat. Phys., 11, 111 (2015)
- Lois, C., Garcia-Verdugo, J. -M., Alvarez-Buylla, A. et al.: Chain migration of neuronal precursors. Science, 271, 978-981 (1996)[PubMed]
- Saw, T. B., Doostmohammadi, A., Nier, V. et al.: Topological defects in epithelia govern cell death and extrusion. Nature, 544, 212-216 (2017)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2015年 東京大学大学院理学系研究科物理学専攻博士課程 修了,同年より米国Harvard Medical School博士研究員.
研究テーマ:生物物理の理論と実験.
影山 龍一郎(Ryoichiro Kageyama)
京都大学ウイルス研究所 教授.
佐野 雅己(Masaki Sano)
東京大学理学系研究科物理学専攻 教授.
© 2017 川口喬吾・影山龍一郎・佐野雅己 Licensed under CC 表示 2.1 日本
