細胞質流動の発生および逆転は小胞体のネットワークにより支配される
木村健二・木村 暁
(国立遺伝学研究所 構造遺伝学研究センター細胞建築研究室)
email:木村健二
DOI: 10.7875/first.author.2017.029
Endoplasmic-reticulum-mediated microtubule alignment governs cytoplasmic streaming.
Kenji Kimura, Alexandre Mamane, Tohru Sasaki, Kohta Sato, Jun Takagi, Ritsuya Niwayama, Lars Hufnagel, Yuta Shimamoto, Jean-François Joanny, Seiichi Uchida, Akatsuki Kimura
Nature Cell Biology, 19, 399-406 (2017)
細胞質流動は細胞の内部における大規模な流れであり,植物の成長や動物の胚の発生において重要な役割を担う.線虫の受精卵においては,受精の直後にキネシンおよび微小管に依存した細胞質流動が生じる.この細胞質流動の方向はあらかじめ定まっておらず,途中で逆転することもある.しかしながら,なぜ受精の直後の卵において集団的な流れが自発的に発生するのか,その機構は不明であった.今回,筆者らは,細胞質流動の際のオルガネラおよび微小管を定量的に解析し,キネシンに依存した小胞体ネットワークの輸送が微小管の整列および集団的な流れに必要であることを発見した.さらに,得られた結果をもとに細胞質流動のふるまいを再現かつ予測する数理モデルを構築し,予測のひとつを実験的に検証することにも成功した.提案された数理モデルはさまざまなタイプの細胞質流動の説明に応用されるかもしれない.
細胞質流動は細胞質の集団的な流れであり,1774年,植物において発見された1).細胞質流動は動物にもみられ,とくに受精の前後の卵において顕著である.細胞質流動の方向はもともとの細胞極性によりあらかじめ決まっている場合が多い.しかし一方で,方向の決まっていない細胞質流動もある.線虫Caenorhabditis elegansの受精卵においては,まだ細胞極性が確立していない減数分裂期に細胞質流動が生じる.この細胞質流動はおもに卵長軸を中心とした回転運動だが,その回転の方向は決まっておらず,途中で方向が逆転することもある.
2010年,減数分裂期の細胞質流動は微小管のプラス端へと移動するモータータンパク質のひとつkinesin-1および微小管に依存することが見い出された2).減数分裂期において微小管はおもに卵の表層から伸びており,多数が流れの方向に近い角度をむき整列している.これら整列した微小管のつくるレールのうえでkinesin-1がカーゴである卵黄顆粒を輸送することにより,集団的な流れが生じると考えられた.つまり,より多くの微小管が同じ方向をむくことにより,安定した細胞質流動が生じる.しかしながら,なぜ微小管が細胞極性のない状況で自発的に整列するのか,その機構は謎であった.
植物における細胞質流動においては細胞に密なネットワーク構造を形成する小胞体の重要性が示唆されてきた.植物の細胞質流動において必要なモータータンパク質は小胞体を輸送するミオシンである3).ミオシンをノックダウンするとそのレールであるアクチン線維の整列が乱れ細胞質流動も生じない.これらのことから,小胞体が輸送されることにより広範囲で効率的に細胞質の流れが生じ,それが隣接するアクチン線維の方向をそろえるのではないかと提案された3).しかし,このような機構が線虫における減数分裂期の細胞質流動に必要な微小管の整列にも寄与するかどうかは不明であった.
この研究においては,線虫における減数分裂期の細胞質流動において微小管が自発的に整列する機構を解明するため,小胞体を含むさまざまなオルガネラの動きおよび微小管の角度を定量的に解析した.さらに,得られた結果をもとに減数分裂期の細胞質流動の発生を説明する数理モデルを構築し,実験的に検証した.
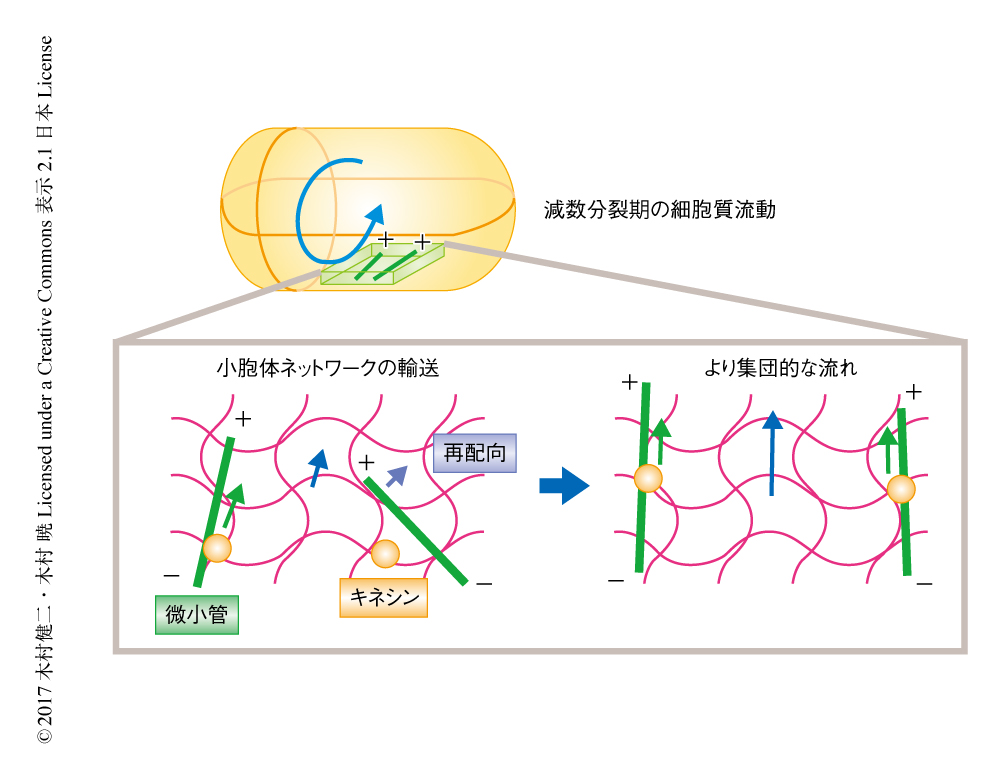
線虫の受精卵において,小胞体は網目状のネットワーク構造を形成し,減数分裂期の細胞質流動の際にダイナミックに流れていることが見い出された.その流れは卵黄顆粒や表層顆粒といったほかのオルガネラの動きとほぼ同じであった.この小胞体の動きはkinesin-1をノックダウンするとみられなくなった.これらの結果から,減数分裂期の細胞質流動の機構として,以下のような正のフィードバックモデルが考案された(図1).1)小胞体が卵の表層から伸びる微小管のうえをkinesin-1により輸送され,局所的な流れが生じる.2)この流れにより周囲の微小管をその方向へむかせるような力が生じる.3)このような正のフィードバックのくり返しがより多くの微小管を整列させる.このモデルにおいて,小胞体のネットワーク構造は重要であった.なぜなら,網目状に広がる小胞体のほうが,顆粒状のオルガネラよりもより効率的に細胞質の流れを生じる力を伝達すると考えられたためである.
減数分裂期の細胞質流動における小胞体の重要性について検証するため,小胞体のネットワーク構造を断片化することにより細胞質流動が阻害されるかどうか確認した.線虫において小胞体はret-1遺伝子およびyop-1遺伝子を同時に抑制することにより断片化される4).ret-1遺伝子およびyop-1遺伝子をRNAi法によりノックダウンした結果,小胞体のネットワーク構造は断片化し,減数分裂期の細胞質流動は阻害された.さらに,アフリカツメガエルの卵抽出液を用いて細胞質の流れに対する小胞体ネットワークの寄与について検証した.卵抽出液において小胞体ネットワークの存在する領域に微小針を刺して引っ張ると,細胞質の顆粒もいっしょに動いた.一方,小胞体ネットワークのない領域に微小針を刺して動かしても細胞質の顆粒は動かなかった.これらの結果から,小胞体ネットワークは細胞質における粘性を高めることが示され,減数分裂期の細胞質流動の機構の正のフィードバックモデルが支持された.
線虫における減数分裂期の細胞質流動において,小胞体ネットワークの流れがないと微小管は整列しなくなるのだろうか? この点について検証するため,小胞体の流動の方向と微小管の角度との関係を画像解析により定量的に調べた.小胞体は卵長軸を中心とした回転運動,つまり,卵短軸の方向にそって流れていた.このとき,卵の表層の付近の微小管は同様におもに卵短軸の方向をむいており,小胞体の流動の方向と一致した.小胞体の流動の方向と微小管の角度との相関は局所的にもみられた.これらのことから,微小管のうえを小胞体が輸送されることが支持された.一方,kinesin-1をノックダウンするとこの微小管の整列は消失し,微小管はランダムな方向をむくようになった.ret-1遺伝子およびyop-1遺伝子をノックダウンして小胞体のネットワーク構造を断片化した際にも微小管の整列は消失した.この結果から,小胞体ネットワークの流れが微小管を整列させるという減数分裂期の細胞質流動の機構の正のフィードバックモデルが支持された.
この正のフィードバックモデルが減数分裂期の細胞質流動を発生させるのに十分かどうか検証するため,新たに数理モデルを構築した.この数理モデルは,1)kinesin-1により微小管にそって小胞体を含む細胞質を動かす力が生じる,2)その力が小胞体を含む細胞質の流れをつくる,3)細胞の表層にアンカーされた微小管はこの流れにより方向を変える(正のフィードバック),4)微小管は伸長と崩壊とを確率的にくり返す,という,これまでの観察の結果にもとづいたものであった.構築された数理モデルは,実際の生理的な範囲のパラメーターのセットにより細胞質流動が発生することを示した.さらに,この数理モデルはパラメーターを変えたときの細胞質流動のふるまいの変化も予測した.
構築された数理モデルは減数分裂期の細胞質流動における方向の逆転についても説明した.これまで,細胞質流動の逆転を説明する数理モデルは存在しなかった.さきに述べたとおり,減数分裂期の細胞質流動は卵長軸を中心とした回転運動であり,これがときおり逆回転する.この逆転は微小管の整列の方向の逆転により起こりうる.微小管の崩壊および伸長の頻度に影響するような細胞におけるノイズを考慮することにより,微小管の整列の方向の確率的な逆転が説明された.
構築された数理モデルの信頼性を確認するため,数理モデルによる予測を実験的に検証した.予測のひとつは,微小管の崩壊の頻度を低下させて微小管を長くすることにより細胞質流動の速度が上昇し,さらに,方向が逆転する頻度も低下するというものであった.微小管を長くするため微小管の切断酵素として知られるkataninのサブユニットをコードするmei-1遺伝子をノックダウンしたところ,微小管は平均で約15%長くなった.このとき,細胞質流動の速度は約60%上昇し,逆転の頻度も1/3に低下した.これらの結果は予測された傾向と一致し,この数理モデルが支持された.
線虫における減数分裂期の細胞質流動の生理的な意義について検証した.線虫の受精卵においては,細胞質に分布する表層顆粒が細胞の外へと分泌されることにより受精卵に浸透圧ストレスに対する耐性能をあたえる5,6).減数分裂期の細胞質流動は表層顆粒が分泌される時期に起こる.そこで,kinesin-1あるいはret-1遺伝子およびyop-1遺伝子をノックダウンし,減数分裂期の細胞質流動が起こらないようにして表層顆粒の分泌を観察した.その結果,一部の表層顆粒は細胞質に残留し,表層までたどりつかず分泌されなかった.この結果と一致して,減数分裂期の細胞質流動を抑制した受精卵の20%は浸透圧ストレスに対し弱くなっていた.これらのことから,減数分裂期の細胞質流動は表層顆粒の動きを促進し,表層の付近まで移動するチャンスをあたえることで分泌に寄与することが示された.
この研究により,小胞体のネットワーク構造が減数分裂期の細胞質流動における集団的な流れの発生に重要な役割をはたすことが明らかにされた.今回の結果から,小胞体ネットワークの媒介する正のフィードバックが微小管を整列させることにより細胞質流動が生じるという正のフィードバックモデルを提案する(図1).小胞体の重要性は植物の細胞質流動におけるアクチン線維の整列においても示されていたことから,動物と植物の細胞質流動に共通する機構が存在するのかもしれない.
細胞質流動はその流れのパターンにより7つに分類され1,7),そのうち,“saltation”(あまりまとまりのない流れ),“circulation”(ときおり方向の変わる流れ),“rotation”(一方向への周回運動)がよくみられる.これらのあいだにあまり明確な区別はなく,植物においてはこれらのパターンのあいだの推移もみられる.ショウジョウバエの卵母細胞における細胞質流動においても,発生のステージによりsaltationからrotationへとパターンが変化する8,9).今回,このような変化が遺伝学的な操作により誘導されることも示した.すなわち,減数分裂期の細胞質流動はときおり方向が逆転することからcirculationである.ret-1遺伝子およびyop-1遺伝子をノックダウンすると集団的な流れは失われsaltationになる.mei-1遺伝子をノックダウンすると逆転の頻度は大幅に低下してrotationになる.構築された数理モデルにおいて,パラメーターを変えることによりこれら3つのタイプの発生の条件が説明された.この研究は,さまざまなタイプの細胞質流動に理論的な枠組みをあたえ,受精卵における微小管の自発的な整列という自己組織化の具体例を示したものである.
略歴:2007年 金沢大学大学院自然科学研究科 修了,同年 国立遺伝学研究所 博士研究員を経て,2012年より同 助教.
研究テーマ:細胞においてオルガネラが適材適所に配置するしくみ.
抱負:今後も細胞においておもしろい動きをみつけ,そのしくみを明らかにしていきたい.
木村 暁(Akatsuki Kimura)
国立遺伝学研究所 教授.
研究室URL:https://www.nig.ac.jp/labs/CelArchi/home_jp.html
© 2017 木村健二・木村 暁 Licensed under CC 表示 2.1 日本
(国立遺伝学研究所 構造遺伝学研究センター細胞建築研究室)
email:木村健二
DOI: 10.7875/first.author.2017.029
Endoplasmic-reticulum-mediated microtubule alignment governs cytoplasmic streaming.
Kenji Kimura, Alexandre Mamane, Tohru Sasaki, Kohta Sato, Jun Takagi, Ritsuya Niwayama, Lars Hufnagel, Yuta Shimamoto, Jean-François Joanny, Seiichi Uchida, Akatsuki Kimura
Nature Cell Biology, 19, 399-406 (2017)
要 約
細胞質流動は細胞の内部における大規模な流れであり,植物の成長や動物の胚の発生において重要な役割を担う.線虫の受精卵においては,受精の直後にキネシンおよび微小管に依存した細胞質流動が生じる.この細胞質流動の方向はあらかじめ定まっておらず,途中で逆転することもある.しかしながら,なぜ受精の直後の卵において集団的な流れが自発的に発生するのか,その機構は不明であった.今回,筆者らは,細胞質流動の際のオルガネラおよび微小管を定量的に解析し,キネシンに依存した小胞体ネットワークの輸送が微小管の整列および集団的な流れに必要であることを発見した.さらに,得られた結果をもとに細胞質流動のふるまいを再現かつ予測する数理モデルを構築し,予測のひとつを実験的に検証することにも成功した.提案された数理モデルはさまざまなタイプの細胞質流動の説明に応用されるかもしれない.
はじめに
細胞質流動は細胞質の集団的な流れであり,1774年,植物において発見された1).細胞質流動は動物にもみられ,とくに受精の前後の卵において顕著である.細胞質流動の方向はもともとの細胞極性によりあらかじめ決まっている場合が多い.しかし一方で,方向の決まっていない細胞質流動もある.線虫Caenorhabditis elegansの受精卵においては,まだ細胞極性が確立していない減数分裂期に細胞質流動が生じる.この細胞質流動はおもに卵長軸を中心とした回転運動だが,その回転の方向は決まっておらず,途中で方向が逆転することもある.
2010年,減数分裂期の細胞質流動は微小管のプラス端へと移動するモータータンパク質のひとつkinesin-1および微小管に依存することが見い出された2).減数分裂期において微小管はおもに卵の表層から伸びており,多数が流れの方向に近い角度をむき整列している.これら整列した微小管のつくるレールのうえでkinesin-1がカーゴである卵黄顆粒を輸送することにより,集団的な流れが生じると考えられた.つまり,より多くの微小管が同じ方向をむくことにより,安定した細胞質流動が生じる.しかしながら,なぜ微小管が細胞極性のない状況で自発的に整列するのか,その機構は謎であった.
植物における細胞質流動においては細胞に密なネットワーク構造を形成する小胞体の重要性が示唆されてきた.植物の細胞質流動において必要なモータータンパク質は小胞体を輸送するミオシンである3).ミオシンをノックダウンするとそのレールであるアクチン線維の整列が乱れ細胞質流動も生じない.これらのことから,小胞体が輸送されることにより広範囲で効率的に細胞質の流れが生じ,それが隣接するアクチン線維の方向をそろえるのではないかと提案された3).しかし,このような機構が線虫における減数分裂期の細胞質流動に必要な微小管の整列にも寄与するかどうかは不明であった.
この研究においては,線虫における減数分裂期の細胞質流動において微小管が自発的に整列する機構を解明するため,小胞体を含むさまざまなオルガネラの動きおよび微小管の角度を定量的に解析した.さらに,得られた結果をもとに減数分裂期の細胞質流動の発生を説明する数理モデルを構築し,実験的に検証した.
1.減数分裂期の細胞質流動には小胞体のネットワーク構造が必要である
線虫の受精卵において,小胞体は網目状のネットワーク構造を形成し,減数分裂期の細胞質流動の際にダイナミックに流れていることが見い出された.その流れは卵黄顆粒や表層顆粒といったほかのオルガネラの動きとほぼ同じであった.この小胞体の動きはkinesin-1をノックダウンするとみられなくなった.これらの結果から,減数分裂期の細胞質流動の機構として,以下のような正のフィードバックモデルが考案された(図1).1)小胞体が卵の表層から伸びる微小管のうえをkinesin-1により輸送され,局所的な流れが生じる.2)この流れにより周囲の微小管をその方向へむかせるような力が生じる.3)このような正のフィードバックのくり返しがより多くの微小管を整列させる.このモデルにおいて,小胞体のネットワーク構造は重要であった.なぜなら,網目状に広がる小胞体のほうが,顆粒状のオルガネラよりもより効率的に細胞質の流れを生じる力を伝達すると考えられたためである.
減数分裂期の細胞質流動における小胞体の重要性について検証するため,小胞体のネットワーク構造を断片化することにより細胞質流動が阻害されるかどうか確認した.線虫において小胞体はret-1遺伝子およびyop-1遺伝子を同時に抑制することにより断片化される4).ret-1遺伝子およびyop-1遺伝子をRNAi法によりノックダウンした結果,小胞体のネットワーク構造は断片化し,減数分裂期の細胞質流動は阻害された.さらに,アフリカツメガエルの卵抽出液を用いて細胞質の流れに対する小胞体ネットワークの寄与について検証した.卵抽出液において小胞体ネットワークの存在する領域に微小針を刺して引っ張ると,細胞質の顆粒もいっしょに動いた.一方,小胞体ネットワークのない領域に微小針を刺して動かしても細胞質の顆粒は動かなかった.これらの結果から,小胞体ネットワークは細胞質における粘性を高めることが示され,減数分裂期の細胞質流動の機構の正のフィードバックモデルが支持された.
2.微小管の整列には小胞体の流れが必要である
線虫における減数分裂期の細胞質流動において,小胞体ネットワークの流れがないと微小管は整列しなくなるのだろうか? この点について検証するため,小胞体の流動の方向と微小管の角度との関係を画像解析により定量的に調べた.小胞体は卵長軸を中心とした回転運動,つまり,卵短軸の方向にそって流れていた.このとき,卵の表層の付近の微小管は同様におもに卵短軸の方向をむいており,小胞体の流動の方向と一致した.小胞体の流動の方向と微小管の角度との相関は局所的にもみられた.これらのことから,微小管のうえを小胞体が輸送されることが支持された.一方,kinesin-1をノックダウンするとこの微小管の整列は消失し,微小管はランダムな方向をむくようになった.ret-1遺伝子およびyop-1遺伝子をノックダウンして小胞体のネットワーク構造を断片化した際にも微小管の整列は消失した.この結果から,小胞体ネットワークの流れが微小管を整列させるという減数分裂期の細胞質流動の機構の正のフィードバックモデルが支持された.
3.細胞質流動を説明する新しい数理モデルの構築および予測
この正のフィードバックモデルが減数分裂期の細胞質流動を発生させるのに十分かどうか検証するため,新たに数理モデルを構築した.この数理モデルは,1)kinesin-1により微小管にそって小胞体を含む細胞質を動かす力が生じる,2)その力が小胞体を含む細胞質の流れをつくる,3)細胞の表層にアンカーされた微小管はこの流れにより方向を変える(正のフィードバック),4)微小管は伸長と崩壊とを確率的にくり返す,という,これまでの観察の結果にもとづいたものであった.構築された数理モデルは,実際の生理的な範囲のパラメーターのセットにより細胞質流動が発生することを示した.さらに,この数理モデルはパラメーターを変えたときの細胞質流動のふるまいの変化も予測した.
構築された数理モデルは減数分裂期の細胞質流動における方向の逆転についても説明した.これまで,細胞質流動の逆転を説明する数理モデルは存在しなかった.さきに述べたとおり,減数分裂期の細胞質流動は卵長軸を中心とした回転運動であり,これがときおり逆回転する.この逆転は微小管の整列の方向の逆転により起こりうる.微小管の崩壊および伸長の頻度に影響するような細胞におけるノイズを考慮することにより,微小管の整列の方向の確率的な逆転が説明された.
構築された数理モデルの信頼性を確認するため,数理モデルによる予測を実験的に検証した.予測のひとつは,微小管の崩壊の頻度を低下させて微小管を長くすることにより細胞質流動の速度が上昇し,さらに,方向が逆転する頻度も低下するというものであった.微小管を長くするため微小管の切断酵素として知られるkataninのサブユニットをコードするmei-1遺伝子をノックダウンしたところ,微小管は平均で約15%長くなった.このとき,細胞質流動の速度は約60%上昇し,逆転の頻度も1/3に低下した.これらの結果は予測された傾向と一致し,この数理モデルが支持された.
4.減数分裂期の細胞質流動の生理的な意義
線虫における減数分裂期の細胞質流動の生理的な意義について検証した.線虫の受精卵においては,細胞質に分布する表層顆粒が細胞の外へと分泌されることにより受精卵に浸透圧ストレスに対する耐性能をあたえる5,6).減数分裂期の細胞質流動は表層顆粒が分泌される時期に起こる.そこで,kinesin-1あるいはret-1遺伝子およびyop-1遺伝子をノックダウンし,減数分裂期の細胞質流動が起こらないようにして表層顆粒の分泌を観察した.その結果,一部の表層顆粒は細胞質に残留し,表層までたどりつかず分泌されなかった.この結果と一致して,減数分裂期の細胞質流動を抑制した受精卵の20%は浸透圧ストレスに対し弱くなっていた.これらのことから,減数分裂期の細胞質流動は表層顆粒の動きを促進し,表層の付近まで移動するチャンスをあたえることで分泌に寄与することが示された.
おわりに
この研究により,小胞体のネットワーク構造が減数分裂期の細胞質流動における集団的な流れの発生に重要な役割をはたすことが明らかにされた.今回の結果から,小胞体ネットワークの媒介する正のフィードバックが微小管を整列させることにより細胞質流動が生じるという正のフィードバックモデルを提案する(図1).小胞体の重要性は植物の細胞質流動におけるアクチン線維の整列においても示されていたことから,動物と植物の細胞質流動に共通する機構が存在するのかもしれない.
細胞質流動はその流れのパターンにより7つに分類され1,7),そのうち,“saltation”(あまりまとまりのない流れ),“circulation”(ときおり方向の変わる流れ),“rotation”(一方向への周回運動)がよくみられる.これらのあいだにあまり明確な区別はなく,植物においてはこれらのパターンのあいだの推移もみられる.ショウジョウバエの卵母細胞における細胞質流動においても,発生のステージによりsaltationからrotationへとパターンが変化する8,9).今回,このような変化が遺伝学的な操作により誘導されることも示した.すなわち,減数分裂期の細胞質流動はときおり方向が逆転することからcirculationである.ret-1遺伝子およびyop-1遺伝子をノックダウンすると集団的な流れは失われsaltationになる.mei-1遺伝子をノックダウンすると逆転の頻度は大幅に低下してrotationになる.構築された数理モデルにおいて,パラメーターを変えることによりこれら3つのタイプの発生の条件が説明された.この研究は,さまざまなタイプの細胞質流動に理論的な枠組みをあたえ,受精卵における微小管の自発的な整列という自己組織化の具体例を示したものである.
文 献
- Kamiya, N.: Protoplasmic Streaming. Springer-Verlag, Vienna (1959)
- McNally, K. L., Martin, J. L., Ellefson, M. et al.: Kinesin-dependent transport results in polarized migration of the nucleus in oocytes and inward movement of yolk granules in meiotic embryos. Dev. Biol., 339, 126-140 (2010)[PubMed]
- Ueda, H., Yokota, E., Kutsuna, N. et al.: Myosin-dependent endoplasmic reticulum motility and F-actin organization in plant cells. Proc. Natl. Acad. Sci. USA, 107, 6894-6899 (2010)[PubMed]
- Audhya, A., Desai, A. & Oegema, K.: A role for Rab5 in structuring the endoplasmic reticulum. J. Cell Biol., 178, 43-56 (2007)[PubMed]
- Kimura, K. & Kimura, A.: Rab6 is required for the exocytosis of cortical granules and the recruitment of separase to the granules during the oocyte-to-embryo transition in Caenorhabditis elegans. J. Cell Sci., 125, 5897-5905 (2012)[PubMed]
- Sato, M., Grant, B. D., Harada, A. et al.: Rab11 is required for synchronous secretion of chondroitin proteoglycans after fertilization in Caenorhabditis elegans. J. Cell Sci., 121, 3177-3186 (2008)[PubMed]
- Allen, N. S. & Allen, R. D.: Cytoplasmic streaming in green plants. Annu. Rev. Biophys. Bioeng., 7, 497-526 (1978)[PubMed]
- Bohrmann, J. & Biber, K.: Cytoskeleton-dependent transport of cytoplasmic particles in previtellogenic to mid-vitellogenic ovarian follicles of Drosophila: time-lapse analysis using video-enhanced contrast microscopy. J. Cell Sci., 107, 849-858 (1994)[PubMed]
- Gutzeit, H. O. & Koppa, R.: Time-lapse film analysis of cytoplasmic streaming during late oogenesis of Drosophila. J. Embryol. Exp. Morphol., 67, 101-111 (1982)
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2007年 金沢大学大学院自然科学研究科 修了,同年 国立遺伝学研究所 博士研究員を経て,2012年より同 助教.
研究テーマ:細胞においてオルガネラが適材適所に配置するしくみ.
抱負:今後も細胞においておもしろい動きをみつけ,そのしくみを明らかにしていきたい.
木村 暁(Akatsuki Kimura)
国立遺伝学研究所 教授.
研究室URL:https://www.nig.ac.jp/labs/CelArchi/home_jp.html
© 2017 木村健二・木村 暁 Licensed under CC 表示 2.1 日本
