マカク属サルの第4次視覚野における色と傾きの処理にかかわる機能的な構造
谷川 久
(米国Vanderbilt大学Department of Psychology)
email:谷川 久
DOI: 10.7875/first.author.2010.062
Functional organization for color and orientation in macaque V4.
Hisashi Tanigawa, Haidong D. Lu & Anna W. Roe
Nature Neuroscience, 13, 1542-1548 (2010)
第4次視覚野は色や形の知覚に重要なはたらきをもつ大脳皮質領野である.この領野は,発見当初はおもに色の視覚情報の処理にはたらく領野として解釈されたが,のちの研究から色だけでなく傾きや両眼視差(奥行き),さらには,より複雑な図形パターンに選択的に応答するニューロンがみつかってきたことから,この領野の視覚における役割については多くの議論がなされてきた.この研究において筆者らは,サルの第4次視覚野における視覚刺激に対する脳活動の分布を内因性光計測法を用いて調べた.その結果,第4次視覚野において,色に選択的な活動を示す領域と傾きに選択的な活動を示す領域とがほぼ分離していることが明らかになった.この発見は,第4次視覚野の機能的な多様性や領野区分に関する長年にわたる論争に一石を投じ,色や形の脳内表現が第4次視覚野において空間的に偏って分布していることを示した.
外界の視覚情報は網膜において神経信号へと変換されたのち,視床の外側膝状体をへて大脳皮質視覚野へと伝達される.視覚野は複数の領野から構成されており,視覚情報は初期視覚野である第1次視覚野,第2次視覚野をへて高次視覚野へと伝達される.過去の生理学的あるいは解剖学的な研究から,初期視覚野において,色の情報(色相や明るさ)を処理する部位と形の情報(輪郭の傾き)を処理する部位がほぼ分離していることがわかっている1,2).具体的には,第1次視覚野ではblob領域とinterblob領域が,第2次視覚野ではthin stripe領域とpale stripe領域があり,それぞれの領野で前者が色の情報を,後者が形の情報をおもに処理しているものと考えられている.厳密には色や形の情報は初期視覚野で完全に分離して処理されているわけではなく,両方の視覚情報をもったニューロンの存在や両方の領域のあいだでの神経連絡も多く報告されている3).第1次視覚野あるいは第2次視覚野より高次の領野においても色と形の情報処理を担う部位が分離しているのかどうかは明らかにされていない.
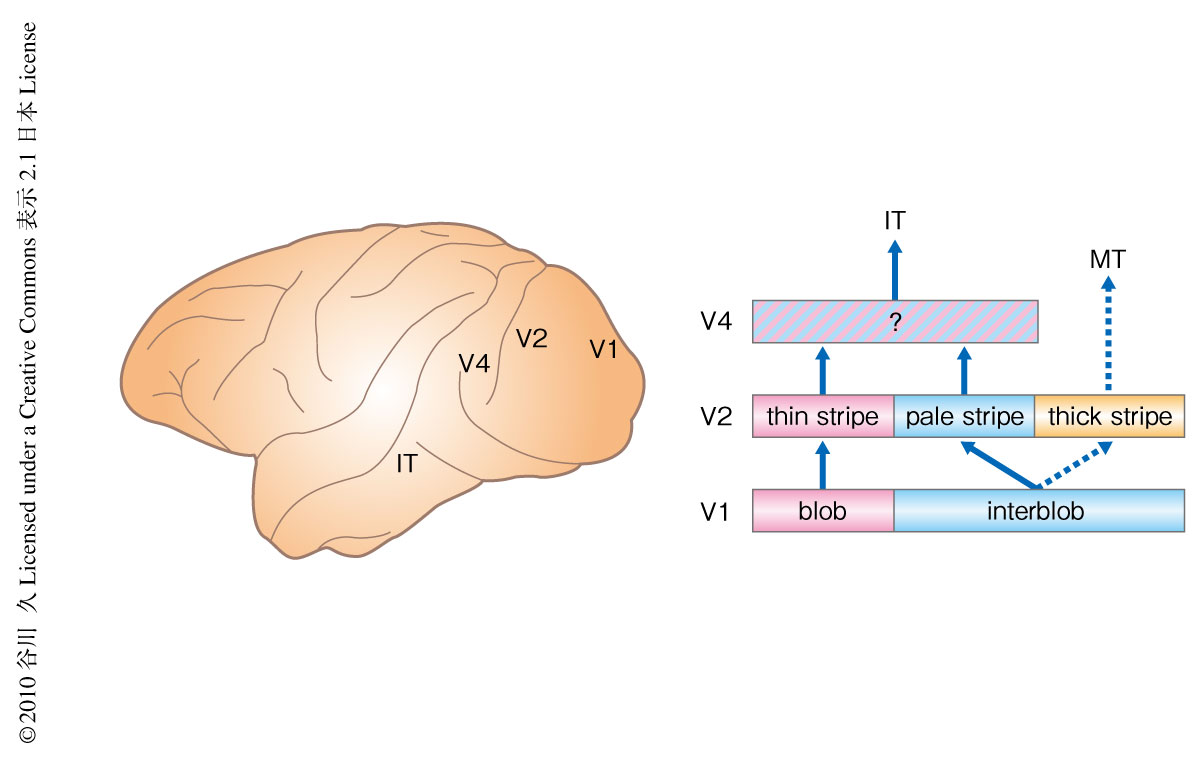
第4次視覚野は初期視覚野において色や形を処理している領域からの神経投射を多くうけており,また,物体認識に重要なはたらきをもつとされる下側頭葉皮質への神経投射を多く行っている4)(図1).そのため,初期視覚野から下側頭葉皮質への物体視にかかわる視覚情報の処理経路の中間段階に位置している.第4次視覚野は,サルにおいて発見された当初,記録されたニューロンのほとんどが色に対して選択的な応答を示したことから,色の情報処理を専門に行なっている領野であると解釈された5).しかしながら,のちの研究から,色以外にも形(輪郭の傾き)や奥行きに対して選択的に応答するニューロンが第4次視覚野から多くみつかり,現在では,さまざまな視覚属性の情報を含んだより高次な視覚情報を処理している領野であると考えられている6).しかしながら,第4次視覚野においても異なった視覚属性(たとえば,色と形)の情報処理を担う部位が分離しているのかどうかはわかっていない.また,過去の生理学的あるいは解剖学的な研究から第4次視覚野の領野区分に関してさまざまな説が提案されているが7),いまだに意見の一致をみていない.第4次視覚野のニューロンの機能的な多様性や領野区分に関する長年にわたる論争を解決し視覚系における第4次視覚野の役割を知るうえで,第4次視覚野の機能的な構造を知ることは重要である.筆者らはこの問いに答えるため,内因性光計測法を用いてマカク属サルの第4次視覚野における色と傾きの処理にかかわる機能的な構造を調べた.
内因性光計測(intrinsic signal optical imaging:ISOI)法は,機能的磁気共鳴画像(functional magnetic resonance imaging:fMRI)法と同様に,ニューロンの活動にともなう脳血液の動態変化を計測し脳活動部位を調べる方法である.ただし,この方法は侵襲的であり,開頭し脳の表面を露出する必要がある.また,計測信号としては還元型ヘモグロビンの局所的な増大による吸光度の変化を利用している8).機能的磁気共鳴画像法に比べて空間解像度が高く,脳の活動を最小で50μmの分解能で観察することができる.そのため,これまでにも大脳皮質の機能的な構造の可視化に多く用いられてきた.有名な例では,第1次視覚野の傾き選択性地図の可視化およびその地図におけるピンホイール構造(ある点を中心に異なる傾きに選択性をもつ部位が放射状に分布する構造)の発見にはこの方法が用いられた9).この研究において筆者らは,内因性光計測法を覚醒サルに適用した.覚醒サルへの内因性光計測法の適応は世界でも限られた研究室でのみ可能となっている技術であり,さまざまな行動課題を遂行中のサルから高解像度の脳機能マッピングを行うことを可能としている.
コンピューターモニター上の点を数秒間にわたり注視することを訓練されたサルに対し,注視中にさまざまな視覚刺激を提示し,刺激によってひき起こされた第4次視覚野の脳活動を内因性光計測法を用いて記録した.第4次視覚野の脳表面は事前に麻酔手術において露出しており,露出部位(約1 cm四方)を透明な人工硬膜と記録用チャンバーによって保護した.露出した脳表面に波長630 nmの光を照射し,その反射光を計測することで神経活動にともなう吸光度の変化を生じた脳部位を同定した.視覚刺激として,等輝度の赤と緑の2色で構成された縞模様(赤緑縞模様),もしくは,無彩色の白と黒で構成された縞模様(白黒縞模様)を用い,灰色の背景のうえに提示した.また,縞模様は水平方向から45度もしくは135度,反時計回りに傾いたものを用いた.
おのおのの視覚刺激によってひき起こされた第4次視覚野の脳活動を調べてみると,部位ごとに強い応答をひき起こす刺激が異なっていることがわかった.そこで,傾きにかかわらず赤緑縞模様か白黒縞模様かで応答の違いを生じる領域を統計的に同定した.その結果,そのような色感受性応答を示す領域は第4次視覚野において幅数ミリの帯状に分布していることがわかった(図2a).同様な方法で,赤緑縞模様か白黒縞模様かにかかわらず縞模様の傾きの違いで応答が異なる領域を調べたところ,そのような傾き感受性応答を示す領域も第4次視覚野で帯状に分布していることがわかった.さらに,このような色感受性領域と傾き感受性領域は第4次視覚野で空間的にほぼ分離していることがわかった.そこで,これらの第4次視覚野の色感受性領域と傾き感受性領域をそれぞれcolor bandとorientation bandと名づけた.これらの領域は色もしくは形の情報処理に関与しているものと考えられた.
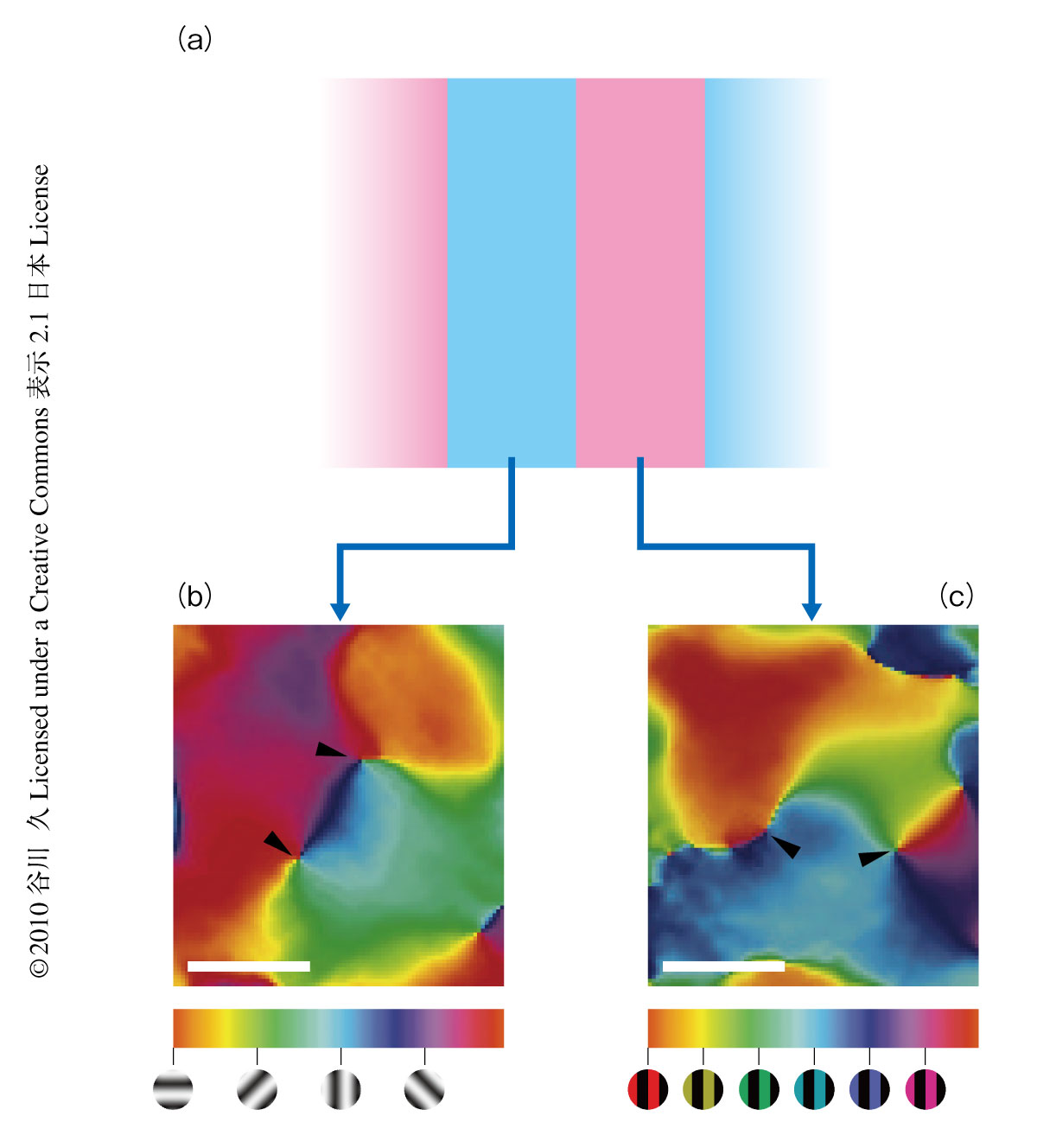
傾き感受性応答を示す領域において,特定の傾きの縞模様に選択的に応答する部位の分布を調べるため,初期視覚野の研究でも用いられている傾き選択性地図(orientation preference map)を作成した(図2b).視覚刺激としては白黒縞模様を用い,その傾きは水平方向から0度,45度,90度もしくは135度,反時計回りに傾いたものを用いた.この地図において,ある色の部位はある特定の傾きの縞模様により強く(選択的に)応答した脳部位を示す.作成された傾き選択性地図から,異なった傾きは傾き感受性領域の異なった脳部位を活性化させることがわかった.このことは,異なった傾きの視覚情報は傾き感受性領域の異なった部位で処理されていることを示唆した.また,第4次視覚野の傾き選択性地図にも第1次視覚野と同様にピンホイール構造が存在することが明らかになった.特定の傾きに選択的に応答する部位(傾き選択性ドメインまたはカラム)は第4次視覚野において平均幅570μmであり,同時に計測された第2次視覚野における傾き選択性ドメイン(400μm)より有意に大きかった.
色感受性応答を示す領域において,特定の色相の縞模様に選択的に応答する部位の分布を調べるため,傾き選択性地図の原理を用いて新たに色相選択性地図(hue preference map)を作成した(図2c).視覚刺激としてはある特定の色と黒とで構成された縞模様を用いた.色は,等輝度の赤,黄,緑,シアン(緑と青の混色),青,マゼンタ(青と赤の混色)のいずれかを使用した.作成された色相選択性地図から,異なった色相は色感受性領域の異なった脳部位を活性化させることがわかった.このことは,異なった色相の情報は色感受性領域の異なった部位で処理されていることを示唆した.また,第4次視覚野の色相選択性地図にもピンホイール構造が存在していた.特定の色相に選択的に応答する部位(色相選択性ドメインまたはカラム)は第4次視覚野において平均幅600μmであり,傾き選択性ドメインと似た大きさであった.また,同時に計測された第2次視覚野における色相選択性ドメイン(370μm)より有意に大きかった.
色相に対するドメインの応答が色の輝度を変化させても変わらないのかどうかを調べるため,同じ色相で異なる輝度をもった縞模様に対する応答を調べた.その結果,少なくともいくつかの色相選択性ドメインに関しては輝度を変化させても色相に対する応答の強度は変化しないことがわかった.このことは,これらの色相選択性ドメインは明るさにかかわらない純粋な色合いの情報の処理を担っていることを示唆していた.
この研究によって,初期視覚野である第1次視覚野や第2次視覚野と同様に,第4次視覚野が色と傾きの処理にかかわる機能区分を構築していることが明らかになった.これまでの電気生理学的な研究から第4次視覚野のニューロンがより複雑な視覚情報を処理していることがわかっており6),色と傾きといった比較的単純な視覚情報の処理のため第4次視覚野が区分されていることは意外なことであった.筆者らはここで,個々の第4次視覚野のニューロンは複雑な視覚情報を処理しているが,その処理している視覚情報が色の情報を含んでいるかそれとも傾きの情報を含んでいるかで,ニューロンがそれぞれ別々の集団を形成しているのではないかと提案している.また,第4次視覚野の神経活動は視覚的な注意により影響をうけることが知られている10).この研究で明らかにされた機能区分はある特定の色や傾きに注意をむける際に神経活動が影響をうける単位となっているのかもしれない.
略歴:2001年 大阪大学大学院医学研究科にて博士(医学)取得,2002年 理化学研究所脳科学総合研究センター 博士研究員を経て,2006年より米国Vanderbilt大学 博士研究員.
研究テーマ:大脳皮質の高次視覚野の機能構築.
抱負:複雑な脳の視覚情報処理の機構を理解するため,細胞レベルから領野レベルまで,機能から構造までを横断的に研究したい.
© 2010 谷川 久 Licensed under CC 表示 2.1 日本
(米国Vanderbilt大学Department of Psychology)
email:谷川 久
DOI: 10.7875/first.author.2010.062
Functional organization for color and orientation in macaque V4.
Hisashi Tanigawa, Haidong D. Lu & Anna W. Roe
Nature Neuroscience, 13, 1542-1548 (2010)
要 約
第4次視覚野は色や形の知覚に重要なはたらきをもつ大脳皮質領野である.この領野は,発見当初はおもに色の視覚情報の処理にはたらく領野として解釈されたが,のちの研究から色だけでなく傾きや両眼視差(奥行き),さらには,より複雑な図形パターンに選択的に応答するニューロンがみつかってきたことから,この領野の視覚における役割については多くの議論がなされてきた.この研究において筆者らは,サルの第4次視覚野における視覚刺激に対する脳活動の分布を内因性光計測法を用いて調べた.その結果,第4次視覚野において,色に選択的な活動を示す領域と傾きに選択的な活動を示す領域とがほぼ分離していることが明らかになった.この発見は,第4次視覚野の機能的な多様性や領野区分に関する長年にわたる論争に一石を投じ,色や形の脳内表現が第4次視覚野において空間的に偏って分布していることを示した.
はじめに
外界の視覚情報は網膜において神経信号へと変換されたのち,視床の外側膝状体をへて大脳皮質視覚野へと伝達される.視覚野は複数の領野から構成されており,視覚情報は初期視覚野である第1次視覚野,第2次視覚野をへて高次視覚野へと伝達される.過去の生理学的あるいは解剖学的な研究から,初期視覚野において,色の情報(色相や明るさ)を処理する部位と形の情報(輪郭の傾き)を処理する部位がほぼ分離していることがわかっている1,2).具体的には,第1次視覚野ではblob領域とinterblob領域が,第2次視覚野ではthin stripe領域とpale stripe領域があり,それぞれの領野で前者が色の情報を,後者が形の情報をおもに処理しているものと考えられている.厳密には色や形の情報は初期視覚野で完全に分離して処理されているわけではなく,両方の視覚情報をもったニューロンの存在や両方の領域のあいだでの神経連絡も多く報告されている3).第1次視覚野あるいは第2次視覚野より高次の領野においても色と形の情報処理を担う部位が分離しているのかどうかは明らかにされていない.
第4次視覚野は初期視覚野において色や形を処理している領域からの神経投射を多くうけており,また,物体認識に重要なはたらきをもつとされる下側頭葉皮質への神経投射を多く行っている4)(図1).そのため,初期視覚野から下側頭葉皮質への物体視にかかわる視覚情報の処理経路の中間段階に位置している.第4次視覚野は,サルにおいて発見された当初,記録されたニューロンのほとんどが色に対して選択的な応答を示したことから,色の情報処理を専門に行なっている領野であると解釈された5).しかしながら,のちの研究から,色以外にも形(輪郭の傾き)や奥行きに対して選択的に応答するニューロンが第4次視覚野から多くみつかり,現在では,さまざまな視覚属性の情報を含んだより高次な視覚情報を処理している領野であると考えられている6).しかしながら,第4次視覚野においても異なった視覚属性(たとえば,色と形)の情報処理を担う部位が分離しているのかどうかはわかっていない.また,過去の生理学的あるいは解剖学的な研究から第4次視覚野の領野区分に関してさまざまな説が提案されているが7),いまだに意見の一致をみていない.第4次視覚野のニューロンの機能的な多様性や領野区分に関する長年にわたる論争を解決し視覚系における第4次視覚野の役割を知るうえで,第4次視覚野の機能的な構造を知ることは重要である.筆者らはこの問いに答えるため,内因性光計測法を用いてマカク属サルの第4次視覚野における色と傾きの処理にかかわる機能的な構造を調べた.
1.内因性光計測法の覚醒サルへの適用
内因性光計測(intrinsic signal optical imaging:ISOI)法は,機能的磁気共鳴画像(functional magnetic resonance imaging:fMRI)法と同様に,ニューロンの活動にともなう脳血液の動態変化を計測し脳活動部位を調べる方法である.ただし,この方法は侵襲的であり,開頭し脳の表面を露出する必要がある.また,計測信号としては還元型ヘモグロビンの局所的な増大による吸光度の変化を利用している8).機能的磁気共鳴画像法に比べて空間解像度が高く,脳の活動を最小で50μmの分解能で観察することができる.そのため,これまでにも大脳皮質の機能的な構造の可視化に多く用いられてきた.有名な例では,第1次視覚野の傾き選択性地図の可視化およびその地図におけるピンホイール構造(ある点を中心に異なる傾きに選択性をもつ部位が放射状に分布する構造)の発見にはこの方法が用いられた9).この研究において筆者らは,内因性光計測法を覚醒サルに適用した.覚醒サルへの内因性光計測法の適応は世界でも限られた研究室でのみ可能となっている技術であり,さまざまな行動課題を遂行中のサルから高解像度の脳機能マッピングを行うことを可能としている.
2.第4次視覚野において色感受性応答を示す領域と傾き感受性応答を示す領域はほぼ分離している
コンピューターモニター上の点を数秒間にわたり注視することを訓練されたサルに対し,注視中にさまざまな視覚刺激を提示し,刺激によってひき起こされた第4次視覚野の脳活動を内因性光計測法を用いて記録した.第4次視覚野の脳表面は事前に麻酔手術において露出しており,露出部位(約1 cm四方)を透明な人工硬膜と記録用チャンバーによって保護した.露出した脳表面に波長630 nmの光を照射し,その反射光を計測することで神経活動にともなう吸光度の変化を生じた脳部位を同定した.視覚刺激として,等輝度の赤と緑の2色で構成された縞模様(赤緑縞模様),もしくは,無彩色の白と黒で構成された縞模様(白黒縞模様)を用い,灰色の背景のうえに提示した.また,縞模様は水平方向から45度もしくは135度,反時計回りに傾いたものを用いた.
おのおのの視覚刺激によってひき起こされた第4次視覚野の脳活動を調べてみると,部位ごとに強い応答をひき起こす刺激が異なっていることがわかった.そこで,傾きにかかわらず赤緑縞模様か白黒縞模様かで応答の違いを生じる領域を統計的に同定した.その結果,そのような色感受性応答を示す領域は第4次視覚野において幅数ミリの帯状に分布していることがわかった(図2a).同様な方法で,赤緑縞模様か白黒縞模様かにかかわらず縞模様の傾きの違いで応答が異なる領域を調べたところ,そのような傾き感受性応答を示す領域も第4次視覚野で帯状に分布していることがわかった.さらに,このような色感受性領域と傾き感受性領域は第4次視覚野で空間的にほぼ分離していることがわかった.そこで,これらの第4次視覚野の色感受性領域と傾き感受性領域をそれぞれcolor bandとorientation bandと名づけた.これらの領域は色もしくは形の情報処理に関与しているものと考えられた.
3.傾き感受性領域において異なった傾きは異なった脳部位を活性化させる
傾き感受性応答を示す領域において,特定の傾きの縞模様に選択的に応答する部位の分布を調べるため,初期視覚野の研究でも用いられている傾き選択性地図(orientation preference map)を作成した(図2b).視覚刺激としては白黒縞模様を用い,その傾きは水平方向から0度,45度,90度もしくは135度,反時計回りに傾いたものを用いた.この地図において,ある色の部位はある特定の傾きの縞模様により強く(選択的に)応答した脳部位を示す.作成された傾き選択性地図から,異なった傾きは傾き感受性領域の異なった脳部位を活性化させることがわかった.このことは,異なった傾きの視覚情報は傾き感受性領域の異なった部位で処理されていることを示唆した.また,第4次視覚野の傾き選択性地図にも第1次視覚野と同様にピンホイール構造が存在することが明らかになった.特定の傾きに選択的に応答する部位(傾き選択性ドメインまたはカラム)は第4次視覚野において平均幅570μmであり,同時に計測された第2次視覚野における傾き選択性ドメイン(400μm)より有意に大きかった.
4.色感受性領域において異なった色相は異なった脳部位を活性化させる
色感受性応答を示す領域において,特定の色相の縞模様に選択的に応答する部位の分布を調べるため,傾き選択性地図の原理を用いて新たに色相選択性地図(hue preference map)を作成した(図2c).視覚刺激としてはある特定の色と黒とで構成された縞模様を用いた.色は,等輝度の赤,黄,緑,シアン(緑と青の混色),青,マゼンタ(青と赤の混色)のいずれかを使用した.作成された色相選択性地図から,異なった色相は色感受性領域の異なった脳部位を活性化させることがわかった.このことは,異なった色相の情報は色感受性領域の異なった部位で処理されていることを示唆した.また,第4次視覚野の色相選択性地図にもピンホイール構造が存在していた.特定の色相に選択的に応答する部位(色相選択性ドメインまたはカラム)は第4次視覚野において平均幅600μmであり,傾き選択性ドメインと似た大きさであった.また,同時に計測された第2次視覚野における色相選択性ドメイン(370μm)より有意に大きかった.
5.色相に対する応答は輝度を変化させても変わらない
色相に対するドメインの応答が色の輝度を変化させても変わらないのかどうかを調べるため,同じ色相で異なる輝度をもった縞模様に対する応答を調べた.その結果,少なくともいくつかの色相選択性ドメインに関しては輝度を変化させても色相に対する応答の強度は変化しないことがわかった.このことは,これらの色相選択性ドメインは明るさにかかわらない純粋な色合いの情報の処理を担っていることを示唆していた.
おわりに
この研究によって,初期視覚野である第1次視覚野や第2次視覚野と同様に,第4次視覚野が色と傾きの処理にかかわる機能区分を構築していることが明らかになった.これまでの電気生理学的な研究から第4次視覚野のニューロンがより複雑な視覚情報を処理していることがわかっており6),色と傾きといった比較的単純な視覚情報の処理のため第4次視覚野が区分されていることは意外なことであった.筆者らはここで,個々の第4次視覚野のニューロンは複雑な視覚情報を処理しているが,その処理している視覚情報が色の情報を含んでいるかそれとも傾きの情報を含んでいるかで,ニューロンがそれぞれ別々の集団を形成しているのではないかと提案している.また,第4次視覚野の神経活動は視覚的な注意により影響をうけることが知られている10).この研究で明らかにされた機能区分はある特定の色や傾きに注意をむける際に神経活動が影響をうける単位となっているのかもしれない.
文 献
- Livingstone, M. S. & Hubel, D. H: Anatomy and physiology of a color system in the primate visual cortex. J. Neurosci., 4, 309-356 (1984)[PubMed]
- Hubel, D. H. & Livingstone, M. S: Segregation of form, color, and stereopsis in primate area 18. J. Neurosci., 7, 3378-3415 (1987)[PubMed]
- Sincich, L. C. & Horton, J. C: The circuitry of V1 and V2: integration of color, form, and motion. Annu. Rev. Neurosci., 28, 303-326 (2005)[PubMed]
- Felleman, D. J. & Van Essen, D. C: Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex, 1, 1-47 (1991)[PubMed]
- Zeki, S. M: Colour coding in rhesus monkey prestriate cortex. Brain Res., 53, 422-427 (1973)[PubMed]
- Orban, G. A: Higher order visual processing in macaque extrastriate cortex. Physiol. Rev., 88, 59-89 (2008)[PubMed]
- Stepniewska, I., Collins, C. E. & Kaas, J. H: Reappraisal of DL/V4 boundaries based on connectivity patterns of dorsolateral visual cortex in macaques. Cereb. Cortex, 809-822 (2005)[PubMed]
- Bonhoeffer, T. & Grinvald, A: Optical imaging based on intrinsic signals: the methodology. in Brain Mapping: the Methods (eds. Toga, A. W. & Mazziotta, J. C.), pp.55-97, Academic Press, New York (1996)
- Bonhoeffer, T. & Grinvald, A: Iso-orientation domains in cat visual cortex are arranged in pinwheel-like patterns. Nature, 353, 429-431 (1991)[PubMed]
- Maunsell, J. H. & Treue, S: Feature-based attention in visual cortex. Trends Neurosci., 29, 317-322 (2006)[PubMed]
著者プロフィール
略歴:2001年 大阪大学大学院医学研究科にて博士(医学)取得,2002年 理化学研究所脳科学総合研究センター 博士研究員を経て,2006年より米国Vanderbilt大学 博士研究員.
研究テーマ:大脳皮質の高次視覚野の機能構築.
抱負:複雑な脳の視覚情報処理の機構を理解するため,細胞レベルから領野レベルまで,機能から構造までを横断的に研究したい.
© 2010 谷川 久 Licensed under CC 表示 2.1 日本
