炎症応答の収束にはマクロファージにおける脂肪酸の代謝のリプログラミングが重要である
大石由美子1・Nathanael J. Spann 2・Christopher K. Glass 2
(1東京医科歯科大学難治疾患研究所 細胞分子医学分野,2米国California大学San Diego校School of Medicine,Department of Cellular and Molecular Medicine)
email:大石由美子
DOI: 10.7875/first.author.2017.008
SREBP1 contributes to resolution of pro-inflammatory TLR4 signaling by reprogramming fatty acid metabolism.
Yumiko Oishi, Nathanael J. Spann, Verena M. Link, Evan D. Muse, Tobias Strid, Chantle Edillor, Matthew J. Kolar, Takashi Matsuzaka, Sumio Hayakawa, Jenhan Tao, Minna U. Kaikkonen, Aaron F. Carlin, Michael T. Lam, Ichiro Manabe, Hitoshi Shimano, Alan Saghatelian, Christopher K. Glass
Cell Metabolism, 25, 412-427 (2017)
マクロファージは病原体の感染に対する防御だけでなく,肥満や糖尿病など生活習慣病の発症につながる慢性炎症の制御においても重要である.この研究において,マクロファージは炎症応答の後期において炎症を抑制するはたらきをもつ不飽和脂肪酸を活発に合成すること,合成された不飽和脂肪酸は身体や組織に生じた炎症応答を抑制し収束させるのに重要であることが明らかにされた.このように,マクロファージの細胞機能としての炎症応答は細胞における脂肪酸の代謝と密接に関連していた.この研究の成果は,マクロファージにおける脂質の合成や代謝を標的とする新しい生活習慣病の治療法および予防法の開発への道を拓くものと期待される.
肥満,糖尿病,動脈硬化症などの生活習慣病や発がんに共通した病態として,慢性炎症が注目されている.慢性炎症は内外のストレスにより生じた炎症応答が適切に収束されずにくすぶった状態と考えられており,組織に障害をひき起こす要因になる.ところが,炎症の慢性化の分子機構は明らかにされていない.慢性炎症においては,多彩な機能をもつマクロファージが炎症の惹起および収束の両面において重要な役割をはたす1).
生体の恒常性をつかさどる免疫系と代謝系は,個体および組織のレベルにおいて密接に連携する.たとえば,肥満においては脂肪組織,肝臓,骨格筋に浸潤する炎症性マクロファージの数が増加し,インスリンシグナルを抑制してインスリン抵抗性をひき起こす2,3).さらに,細胞のレベルにおいても炎症は代謝と密接に関連する.マクロファージは炎症をひき起こす刺激をうけ活性化するとすみやかにHIF-1αやNF-κBを介して解糖系を活性化し,酸素の濃度の低い炎症組織においても活動し殺菌作用を示すようになる.一方,組織の修復やリモデリングをつかさどるマクロファージは細胞における代謝を酸化的リン酸化にシフトし炎症を抑制する表現型を呈する4).
また,マクロファージは種々の脂質を合成することが明らかにされている5).炎症の刺激をうけ活性化されたマクロファージは一過性にエイコサノイドの合成を増加させるが,炎症応答の後期にはスフィンゴ脂質やステロールを合成するようになる.また,エイコサヘキサエン酸やドコサヘキサエン酸などの不飽和脂肪酸はマクロファージにおいて抗炎症性を示すことが知られている6).この研究においては,マクロファージの機能としての炎症応答が代謝と協調して制御され炎症の慢性化にも影響をおよぼすとの仮説を検証した.
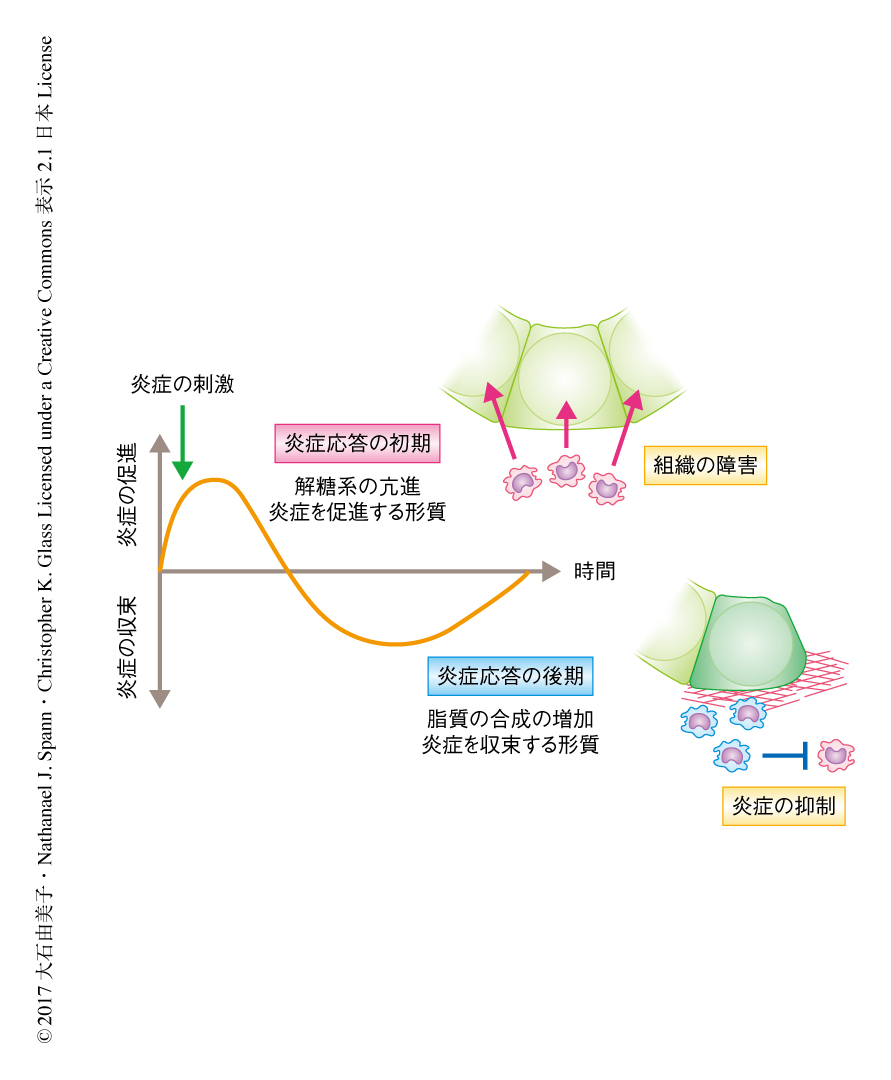
初代培養マクロファージにおいてToll様受容体(Toll-like receptor:TLR)のひとつTLR4を活性化し炎症応答を惹起した際の代謝を観察した.その結果,炎症応答の初期においては解糖系を亢進させ炎症を促進する形質を示したが,炎症応答の後期においては脂肪酸の合成を増加させ炎症を収束する形質に変化した(図1).炎症応答の過程において脂質の合成がダイナミックに変化したことから,炎症応答においてマクロファージにおける種々の脂肪酸の量の変化をリピドミクス法を用いて網羅的に定量した.炎症の刺激をうけたマクロファージにおいては脂肪酸の量は一過性に減少したが,炎症応答の後期になると不飽和脂肪酸の合成が増加に転じた.さらに,RNA-seq法による解析の結果,Scd2遺伝子やFadsなど脂肪酸の不飽和化に関連する遺伝子の発現についても,炎症応答の初期に低下し後期に上昇した.
炎症応答における脂肪酸の合成の制御の分子機構を明らかにするため,ChIP-seq法およびRNA-seq法を用いて炎症応答における全体的な転写およびエピゲノムの動態について解析した.定常状態においてはScd2遺伝子やFads遺伝子など脂肪酸の不飽和化に関連する遺伝子の発現制御領域には核内受容体LXRが結合し発現を維持していた.ところが,炎症応答の初期においてはLXRの機能はNF-κBに依存してすみやかに抑制され,Scd2遺伝子やFadsの発現は低下し抗炎症性の脂肪酸の合成は一過性に抑制された.このことから,炎症応答の初期における脂肪酸の合成の減少は,定常状態において脂肪酸の合成を担うLXRの機能が一時的に抑制されるため生じることが明らかにされた.
炎症応答の後期において不飽和脂肪酸の合成が増加に転じる分子機構を明らかにするため,LXRを欠損したマクロファージ7) において炎症応答における遺伝子の発現の変化を観察した.炎症応答の後期における脂肪酸の合成の増加もLXRに依存的であるなら,LXRを欠損したマクロファージにおいてその増加はみられないはずである.ところが,LXRを欠損したマクロファージにおいても野生型のマクロファージと同様に,炎症応答の後期において不飽和脂肪酸の合成に関連する遺伝子の発現が上昇した.そこで,炎症応答の初期に発現が低下し後期に上昇するような遺伝子のエンハンサー領域についてモチーフ解析をした.その結果,マクロファージに特異的な転写制御に重要な転写因子としてPU.1,C/EBP,AP-1にひきつづき,転写因子SREBPの結合するシス配列SREが上位にランクインした.
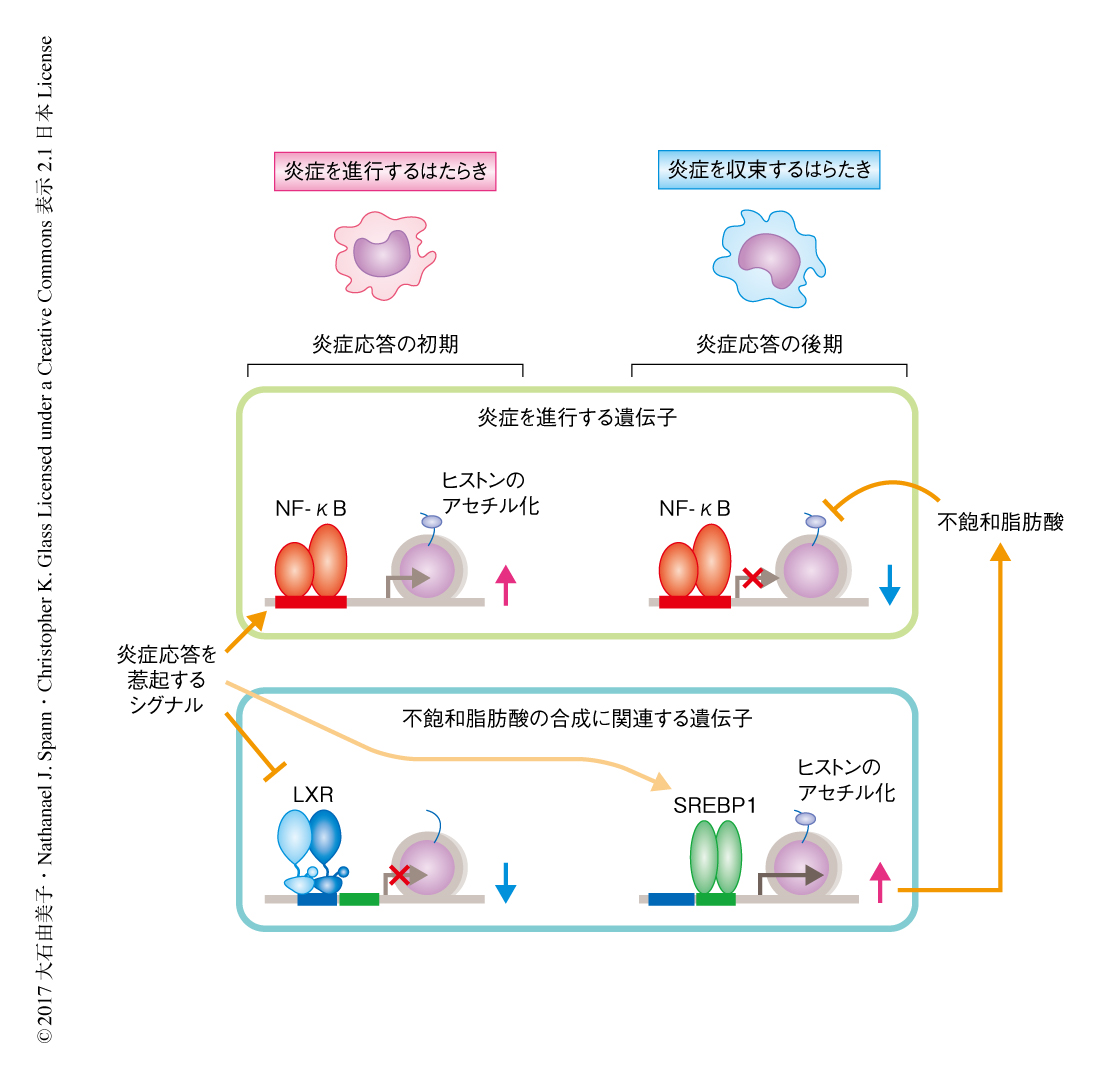
これまで,マクロファージにおいてSREBP1はLXRと協調してコレステロールおよび脂肪酸の代謝を制御することが報告されている8).SREBP1は定常状態においては小胞体膜タンパク質として存在するが,活性化の刺激がくわわるとゴルジ体へと移行して切断をうけ,活性化されたSREBP1は核へと移行して脂質の合成に関連する遺伝子の発現を上昇させることが知られている.そこで,核に存在する活性化型のSREBP1をウェスタンブロッティング法により検出したところ,炎症の刺激をくわえたマクロファージにおいては炎症応答の後期に小胞体膜からのプロセシングをへてSREBP1が活性化されることが明らかにされた.さらに,ChIP-seq法により転写およびエピゲノムの動態を解析した結果,活性化されたSREBP1はScd2遺伝子やFadsなど脂肪酸の不飽和化に関連する遺伝子の転写制御領域にリクルートされ,転写の活性化と相関するヒストンのアセチル化を増加させて抗炎症性の脂肪酸の合成を増加させることが明らかにされた(図2).
SREBP1の重要性についてさらに明らかにするため,Srebp1ノックアウトマウス9) を用いて検討した.Srebp1を欠損するマクロファージは炎症応答の後期において不飽和脂肪酸の合成が減少していただけでなく炎症応答が遷延していた.Srebp1ノックアウトマウスにおいても敗血症モデルによる全身の炎症応答が遷延したが,抗炎症性の不飽和脂肪酸であるエイコサヘキサエン酸を前投与すると炎症応答の遷延は回避された.このことから,マクロファージにおいて炎症応答の後期にて合成される不飽和脂肪酸は炎症応答の適切な制御に必須であることが明らかにされた.
この研究の成果は,マクロファージにおける脂質の代謝と機能との連携が生活習慣病に対する新しい治療の標的として有効である可能性を示し,細胞における代謝を標的とした治療法の開発の基盤になるものである.
略歴:2006年 東京大学大学院医学系研究科博士課程 修了.同 特任助教,米国California大学San Diego校School of Medicine研究員を経て,2013年より東京医科歯科大学難治疾患研究所 准教授.
研究テーマ:代謝,炎症,再生のあいだの連携による生活習慣病における分子機構.
関心事:いっしょに研究を進めてくれる大学院生・研究員を募集中.
研究室URL:http://www.tmd.ac.jp/dcmm/index.html
Nathanael J. Spann
米国California大学San Diego校School of MedicineにてProject Scientist.
Christopher K. Glass
米国California大学San Diego校School of MedicineにてProfessor.
研究室URL:http://cmm.ucsd.edu/glass/glasslab/index.html
© 2017 大石由美子・Nathanael J. Spann・Christopher K. Glass Licensed under CC 表示 2.1 日本
(1東京医科歯科大学難治疾患研究所 細胞分子医学分野,2米国California大学San Diego校School of Medicine,Department of Cellular and Molecular Medicine)
email:大石由美子
DOI: 10.7875/first.author.2017.008
SREBP1 contributes to resolution of pro-inflammatory TLR4 signaling by reprogramming fatty acid metabolism.
Yumiko Oishi, Nathanael J. Spann, Verena M. Link, Evan D. Muse, Tobias Strid, Chantle Edillor, Matthew J. Kolar, Takashi Matsuzaka, Sumio Hayakawa, Jenhan Tao, Minna U. Kaikkonen, Aaron F. Carlin, Michael T. Lam, Ichiro Manabe, Hitoshi Shimano, Alan Saghatelian, Christopher K. Glass
Cell Metabolism, 25, 412-427 (2017)
この論文に出現する遺伝子・タンパク質のUniprot ID
SREBP1(Q9WTN3), TLR4(Q9QUK6), インスリン, HIF-1α(Q61221), NF-κB, Toll様受容体, Toll-like receptor, TLR, Scd2(P13011), Fads, LXR, PU.1(P17433), C/EBP, AP-1, SREBP, ヒストン, Srebp1(Q9WTN3)
要 約
マクロファージは病原体の感染に対する防御だけでなく,肥満や糖尿病など生活習慣病の発症につながる慢性炎症の制御においても重要である.この研究において,マクロファージは炎症応答の後期において炎症を抑制するはたらきをもつ不飽和脂肪酸を活発に合成すること,合成された不飽和脂肪酸は身体や組織に生じた炎症応答を抑制し収束させるのに重要であることが明らかにされた.このように,マクロファージの細胞機能としての炎症応答は細胞における脂肪酸の代謝と密接に関連していた.この研究の成果は,マクロファージにおける脂質の合成や代謝を標的とする新しい生活習慣病の治療法および予防法の開発への道を拓くものと期待される.
はじめに
肥満,糖尿病,動脈硬化症などの生活習慣病や発がんに共通した病態として,慢性炎症が注目されている.慢性炎症は内外のストレスにより生じた炎症応答が適切に収束されずにくすぶった状態と考えられており,組織に障害をひき起こす要因になる.ところが,炎症の慢性化の分子機構は明らかにされていない.慢性炎症においては,多彩な機能をもつマクロファージが炎症の惹起および収束の両面において重要な役割をはたす1).
生体の恒常性をつかさどる免疫系と代謝系は,個体および組織のレベルにおいて密接に連携する.たとえば,肥満においては脂肪組織,肝臓,骨格筋に浸潤する炎症性マクロファージの数が増加し,インスリンシグナルを抑制してインスリン抵抗性をひき起こす2,3).さらに,細胞のレベルにおいても炎症は代謝と密接に関連する.マクロファージは炎症をひき起こす刺激をうけ活性化するとすみやかにHIF-1αやNF-κBを介して解糖系を活性化し,酸素の濃度の低い炎症組織においても活動し殺菌作用を示すようになる.一方,組織の修復やリモデリングをつかさどるマクロファージは細胞における代謝を酸化的リン酸化にシフトし炎症を抑制する表現型を呈する4).
また,マクロファージは種々の脂質を合成することが明らかにされている5).炎症の刺激をうけ活性化されたマクロファージは一過性にエイコサノイドの合成を増加させるが,炎症応答の後期にはスフィンゴ脂質やステロールを合成するようになる.また,エイコサヘキサエン酸やドコサヘキサエン酸などの不飽和脂肪酸はマクロファージにおいて抗炎症性を示すことが知られている6).この研究においては,マクロファージの機能としての炎症応答が代謝と協調して制御され炎症の慢性化にも影響をおよぼすとの仮説を検証した.
1.炎症応答においてマクロファージの代謝はダイナミックに変動する
初代培養マクロファージにおいてToll様受容体(Toll-like receptor:TLR)のひとつTLR4を活性化し炎症応答を惹起した際の代謝を観察した.その結果,炎症応答の初期においては解糖系を亢進させ炎症を促進する形質を示したが,炎症応答の後期においては脂肪酸の合成を増加させ炎症を収束する形質に変化した(図1).炎症応答の過程において脂質の合成がダイナミックに変化したことから,炎症応答においてマクロファージにおける種々の脂肪酸の量の変化をリピドミクス法を用いて網羅的に定量した.炎症の刺激をうけたマクロファージにおいては脂肪酸の量は一過性に減少したが,炎症応答の後期になると不飽和脂肪酸の合成が増加に転じた.さらに,RNA-seq法による解析の結果,Scd2遺伝子やFadsなど脂肪酸の不飽和化に関連する遺伝子の発現についても,炎症応答の初期に低下し後期に上昇した.
2.炎症応答の初期には定常状態において脂肪酸の合成を担う核内受容体LXRの機能が一時的に抑制される
炎症応答における脂肪酸の合成の制御の分子機構を明らかにするため,ChIP-seq法およびRNA-seq法を用いて炎症応答における全体的な転写およびエピゲノムの動態について解析した.定常状態においてはScd2遺伝子やFads遺伝子など脂肪酸の不飽和化に関連する遺伝子の発現制御領域には核内受容体LXRが結合し発現を維持していた.ところが,炎症応答の初期においてはLXRの機能はNF-κBに依存してすみやかに抑制され,Scd2遺伝子やFadsの発現は低下し抗炎症性の脂肪酸の合成は一過性に抑制された.このことから,炎症応答の初期における脂肪酸の合成の減少は,定常状態において脂肪酸の合成を担うLXRの機能が一時的に抑制されるため生じることが明らかにされた.
3.炎症応答の後期には転写因子SREBP1が活性化され不飽和脂肪酸の合成が増加する
炎症応答の後期において不飽和脂肪酸の合成が増加に転じる分子機構を明らかにするため,LXRを欠損したマクロファージ7) において炎症応答における遺伝子の発現の変化を観察した.炎症応答の後期における脂肪酸の合成の増加もLXRに依存的であるなら,LXRを欠損したマクロファージにおいてその増加はみられないはずである.ところが,LXRを欠損したマクロファージにおいても野生型のマクロファージと同様に,炎症応答の後期において不飽和脂肪酸の合成に関連する遺伝子の発現が上昇した.そこで,炎症応答の初期に発現が低下し後期に上昇するような遺伝子のエンハンサー領域についてモチーフ解析をした.その結果,マクロファージに特異的な転写制御に重要な転写因子としてPU.1,C/EBP,AP-1にひきつづき,転写因子SREBPの結合するシス配列SREが上位にランクインした.
これまで,マクロファージにおいてSREBP1はLXRと協調してコレステロールおよび脂肪酸の代謝を制御することが報告されている8).SREBP1は定常状態においては小胞体膜タンパク質として存在するが,活性化の刺激がくわわるとゴルジ体へと移行して切断をうけ,活性化されたSREBP1は核へと移行して脂質の合成に関連する遺伝子の発現を上昇させることが知られている.そこで,核に存在する活性化型のSREBP1をウェスタンブロッティング法により検出したところ,炎症の刺激をくわえたマクロファージにおいては炎症応答の後期に小胞体膜からのプロセシングをへてSREBP1が活性化されることが明らかにされた.さらに,ChIP-seq法により転写およびエピゲノムの動態を解析した結果,活性化されたSREBP1はScd2遺伝子やFadsなど脂肪酸の不飽和化に関連する遺伝子の転写制御領域にリクルートされ,転写の活性化と相関するヒストンのアセチル化を増加させて抗炎症性の脂肪酸の合成を増加させることが明らかにされた(図2).
4.マクロファージによる不飽和脂肪酸の合成は炎症応答の適切な収束に必須である
SREBP1の重要性についてさらに明らかにするため,Srebp1ノックアウトマウス9) を用いて検討した.Srebp1を欠損するマクロファージは炎症応答の後期において不飽和脂肪酸の合成が減少していただけでなく炎症応答が遷延していた.Srebp1ノックアウトマウスにおいても敗血症モデルによる全身の炎症応答が遷延したが,抗炎症性の不飽和脂肪酸であるエイコサヘキサエン酸を前投与すると炎症応答の遷延は回避された.このことから,マクロファージにおいて炎症応答の後期にて合成される不飽和脂肪酸は炎症応答の適切な制御に必須であることが明らかにされた.
おわりに
この研究の成果は,マクロファージにおける脂質の代謝と機能との連携が生活習慣病に対する新しい治療の標的として有効である可能性を示し,細胞における代謝を標的とした治療法の開発の基盤になるものである.
文 献
- Tabas, I. & Glass, C. K.: Anti-inflammatory therapy in chronic disease: challenges and opportunities. Science, 339, 166-172 (2013)[PubMed]
- Lumeng, C. N., DelProposto, J. B., Westcott, D. J. et al.: Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes, 57, 3239-3246 (2008)[PubMed]
- Osborn, O. & Olefsky, J. M.: The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med., 18, 363-374 (2012)[PubMed]
- Mantovani, A., Sozzani, S., Locati, M. et al.: Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol., 23, 549-555 (2002)[PubMed]
- Dennis, E. A., Deems, R. A., Harkewicz, R. et al.: A mouse macrophage lipidome. J. Biol. Chem., 285, 39976-39985 (2010)[PubMed]
- Oh, D. Y., Talukdar, S., Bae, E. J. et al.: GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell, 142, 687-698 (2010)[PubMed]
- Repa, J. J., Turley, S. D., Lobaccaro, J. A. et al.: Regulation of absorption and ABC1-mediated efflux of cholesterol by RXR heterodimers. Science, 289, 1524-1529 (2000)[PubMed]
- Spann, N. J., Garmire, L. X., McDonald, J. G. et al.: Regulated accumulation of desmosterol integrates macrophage lipid metabolism and inflammatory responses. Cell, 151, 138-152 (2012)[PubMed]
- Shimano, H., Shimomura, I., Hammer, R. E. et al.: Elevated levels of SREBP-2 and cholesterol synthesis in livers of mice homozygous for a targeted disruption of the SREBP-1 gene. J. Clin. Invest., 100, 2115-2124 (1997)[PubMed]
著者プロフィール
略歴:2006年 東京大学大学院医学系研究科博士課程 修了.同 特任助教,米国California大学San Diego校School of Medicine研究員を経て,2013年より東京医科歯科大学難治疾患研究所 准教授.
研究テーマ:代謝,炎症,再生のあいだの連携による生活習慣病における分子機構.
関心事:いっしょに研究を進めてくれる大学院生・研究員を募集中.
研究室URL:http://www.tmd.ac.jp/dcmm/index.html
Nathanael J. Spann
米国California大学San Diego校School of MedicineにてProject Scientist.
Christopher K. Glass
米国California大学San Diego校School of MedicineにてProfessor.
研究室URL:http://cmm.ucsd.edu/glass/glasslab/index.html
© 2017 大石由美子・Nathanael J. Spann・Christopher K. Glass Licensed under CC 表示 2.1 日本
