セロトニン性の神経回路による運動学習の機構
川島 尚之
(米国Howard Hughes Medical Institute,Janelia Research Campus)
email:川島尚之
DOI: 10.7875/first.author.2016.118
The serotonergic system tracks the outcomes of actions to mediate short-term motor learning.
Takashi Kawashima, Maarten F. Zwart, Chao-Tsung Yang, Brett D. Mensh, Misha B. Ahrens
Cell, 167, 933-946.e20 (2016)
外部の環境がたえず変動するなかでもつねに意図したとおりの動作ができるよう,動物の運動は学習の機構によりたえず制御される.これまで,運動の際にセロトニン性の神経回路が活性化することは知られていたが,具体的にどのように運動の制御に関与するのかは不明であった.筆者らは,この研究において,ゼブラフィッシュの全脳Ca2+イメージング法により縫線核のセロトニン系の神経回路が短期の運動学習において中心的な役割を担うことを発見した.さらに詳細な解析により,縫線核のニューロンには運動指令と同期した視覚の刺激への選択的な反応,および,くり返しの学習による持続的なニューロンの発火の蓄積,という2種類の計算原理の存在することが見い出された.この研究による運動学習のシンプルな理解から,セロトニン系の神経回路の多様な機能に共通する原理の解明につながることが期待される.
縫線核に代表されるセロトニン系の神経回路は報酬学習や恐怖記憶といった幅広い情動行動にかかわることが知られ,うつ病に代表される精神疾患においても注目されている.これまで,セロトニンの伝達を制御する治療薬を中心に薬理学的な行動解析は幅広く行われてきたものの,セロトニン系の神経回路における神経機構に着目した研究は少なく,その幅広い役割に共通する原理は未解明なままである1).その理由のひとつとして,情動行動や精神疾患にかかわる行動の要素は複雑であり,単純な要素に分解した解析が困難なことがあげられる.
運動学習は動物の行動においてもっとも基本的な学習形態のひとつであり,脳からの運動指令の出力ののち,1)動作の結果を感知し,2)その情報を保存し,3)つぎの運動指令を修正する,という過程のくり返しにより担われる.これまでの長期の運動学習の研究により,この過程が小脳や大脳皮質の運動野のシナプス可塑性により担われることは解明されたものの,数分以内に起こるような短期の運動学習の機構についてはあまり知られていない.また,動物の運動の際に縫線核のニューロンが活性化することは知られていたものの2),その運動制御における具体的な役割は不明であった.
この研究においては,短期の運動学習の機構の解明をめざし,仮想環境においたゼブラフィッシュに対し全脳Ca2+イメージング法を適用した.
短期の運動学習における責任神経回路を同定するため,ゼブラフィッシュの稚魚に対し仮想環境を用いて運動学習のパラダイムを構築した.この仮想環境において,ゼブラフィッシュの動きを固定するため筋肉を麻痺させ,筋肉へと伸びる脊髄運動ニューロンの軸索から電気生理学的に運動シグナルを記録した.ゼブラフィッシュはその下に投影された仮想環境を見ることができる.この仮想環境を前方に動かすと,ゼブラフィッシュはこれを本能的に追従しようとして運動シグナルを出力する.運動シグナルが記録されると仮想環境は後方に動き,ゼブラフィッシュはこれを見て自分が前進したと錯覚する3,4).この仮想環境においてゼブラフィッシュは運動学習を行う.水中においては水の温度や粘性の変化により意図した泳ぎの強さと実際の移動距離とにずれが生じる.このとき,泳ぎの強さに対して得られた移動距離をゲインといい,これに応じてゼブラフィッシュは運動の強さを制御する.仮想環境においては,記録された運動シグナルと仮想環境の動きとのあいだのゲインの値を操作することにより,ゼブラフィッシュの運動出力の強度を変化させることができる.ゲインが低いとゼブラフィッシュは前進するため強い運動シグナルを出力する必要がある.逆に,ゲインが高いとゼブラフィッシュは弱い運動シグナルでも十分に前進することができる.ゲインを継続的に高くする訓練を行えば,ゼブラフィッシュは運動学習により弱い運動シグナルを継続して出力するようになり,この弱い運動シグナルの出力は,たとえば,10秒間にわたり運動を停止させたのち,もういちど仮想環境を提示しても継続される.これが,この研究における“短期の運動学習”である.
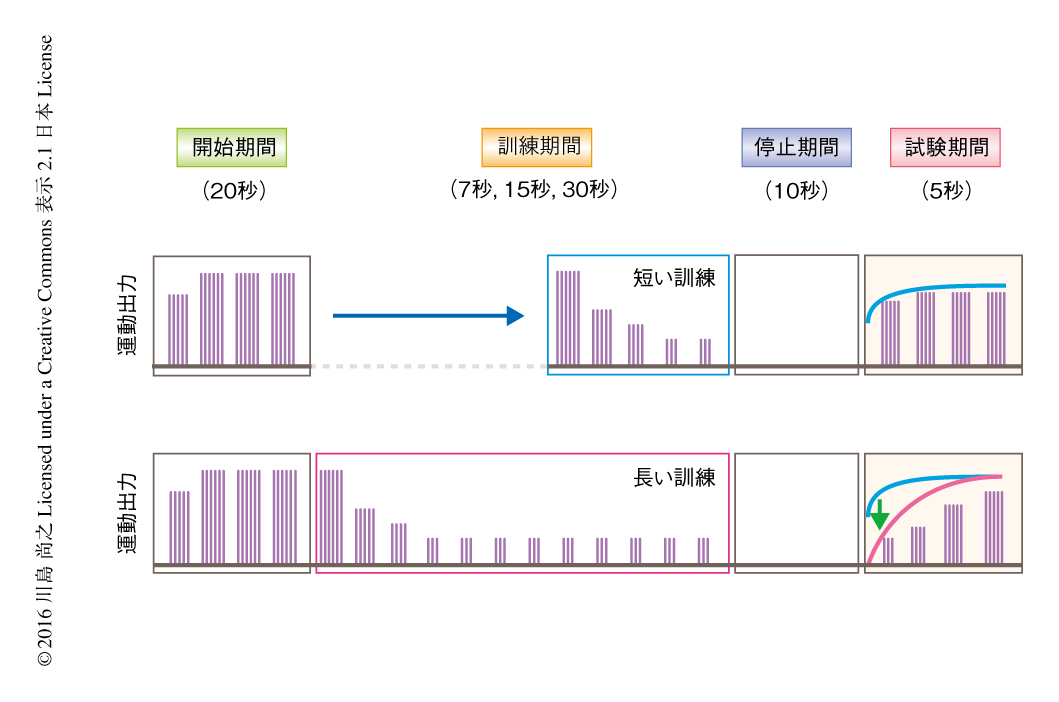
学習の程度が訓練する時間の長さに比例することに注目して行動のパラダイムを開発した.このパラダイムは“開始期間”“訓練期間”“停止期間”“試験期間”の4つの期間から構成される(図1).開始期間において仮想環境のゲインは低く設定され,ゼブラフィッシュは強い運動シグナルを継続的に出力する.訓練期間において仮想環境のゲインは高く設定され,ゼブラフィッシュは短い時間(7秒)あるいは長い時間(30秒)にわたり弱い運動シグナルを継続的に出力する.停止期間において仮想環境は停止し,ゼブラフィッシュの運動も停止する.試験期間において仮想環境はもういちど前方に動き,それに応答したゼブラフィッシュの運動シグナルの強度を測定する.その結果,長い訓練を行った場合には,短い訓練の場合と比較して試験期間においてより弱い運動シグナルが記録された(図1).訓練中の運動シグナルの強度は時間の長さに関係なく一定であるため,脳にはくり返しの回数に依存して学習の効果が蓄積するような過程の存在することが示唆された.さらには,試験期間におけるゼブラフィッシュの反応時間や反応率は訓練の長さに対し不変であったことから,この学習はゼブラフィッシュの感覚応答性や覚醒状態には影響せず運動出力に特異的に作用することが示された.
この行動の責任領域を探索するため,運動学習中の脳における神経活動をライトシート顕微鏡により記録した.すべてのニューロンにCa2+指示タンパク質であるGCaMP6 5) を発現させた遺伝子改変ゼブラフィッシュに対し,2方向から励起光を照射することにより仮想環境において行動しているあいだ,全脳の8割以上,約10万個ものニューロンの神経活動が1~2 Hzで同時記録された6).
訓練中に蓄積された学習の情報を停止期間をこえて保持するニューロンを探索したところ,脳幹の前部に強い集積がみられた.過去の文献および免疫染色法により,これはセロトニン性の背側縫線核であると同定された.縫線核のセロトニン性のニューロンは訓練期間において時間に依存的に神経活動を蓄積し,停止期間において蓄積された神経活動が緩徐に減衰し,蓄積された学習の情報は試験期間まで保持された.停止期間における縫線核の神経活動が高いほど,つぎの試験期間における運動出力の強度は抑制された.
背側縫線核の運動学習への関与についてさらに調べるため,その神経活動を操作した.薬理学的な手法によりセロトニンの伝達を遮断する,あるいは,化学遺伝学的な手法によりセロトニン性のニューロンを選択的に細胞死させることにより,ゼブラフィッシュの短期の学習行動は完全に阻害された.また,光遺伝学的な手法により停止期間において背側縫線核の神経活動を上昇させると短期の学習行動は強化された.これらの結果から,背側縫線核の神経活動が短期の運動学習において中心的な役割をはたすことが示された.
縫線核のニューロンは運動学習中にどのような計算を行うのであろうか? その詳細な神経活動について明らかにするため,運動学習中のゼブラフィッシュの縫線核のニューロンから2光子顕微鏡による観察のもと細胞外記録を行った.ゼブラフィッシュの小さな脳に細胞外電極を挿入すると大半は行動をとめてしまうためこの実験は困難をきわめたが,少数ながら学習行動を示すゼブラフィッシュから記録を行った.その結果,縫線核のニューロンは訓練期間において個別の泳ぎの直後にバースト様の発火を示すこと,そして,くり返しの訓練が進むにつれ泳ぎに同期しない持続的な発火が蓄積することがわかった.また,このニューロンの持続的な発火は停止期間においても持続した.このニューロンの持続的な発火の蓄積は報酬学習の際にも起こることが報告されており7),セロトニン系の幅広い機能に共通することが示唆される.
では,個別の泳ぎの直後のニューロンのバースト様の発火はどのような情報をコードするのであろうか? コードされる情報は運動の強度であろうか,それとも,運動の直後の仮想環境の動きであろうか? このニューロンのバースト様の発火は高速のCa2+イメージング法によっても個別の運動出力の直後に観察されたため,細胞の大規模な解析が可能なCa2+イメージング法により解析した.仮想環境の条件をくふうすることにより,運動出力の強度を同じにしたまま,仮想環境の動きがゲインに比例して速くなる条件を構築した.この場合,縫線核のニューロンのバースト様の発火は仮想環境の動きの速さに比例して増大した.逆に,異なる運動出力の強度に対し仮想環境の動く速度が変わらない条件を設定すると,縫線核のニューロンのバースト様の発火は変化しなかった.このことから,縫線核のセロトニン性のニューロンのバースト様の発火は運動出力に付随して起こる仮想環境の動きをコードすることがわかった.
しかしながら,ここでひとつの疑問が浮上した.ゼブラフィッシュは自らの動きにより起こった周囲の環境の動きと,外部の力により動いた周囲の環境の動きの両方を見ることができる.縫線核において正確な運動学習を成立させるためには,自らの動きに付随する視覚入力を選択的に学習する必要がある.この疑問に答えるため,仮想環境の動きが運動出力と非同期になる状態をつくり,仮想環境の動きがゼブラフィッシュの泳ぎと同時に起こった場合と時間的にずれた場合との縫線核のニューロンのバースト様の発火を比較した.その結果,縫線核は周囲の環境の動きがゼブラフィッシュの運動出力と同期した場合にのみ反応することが示された(図2).
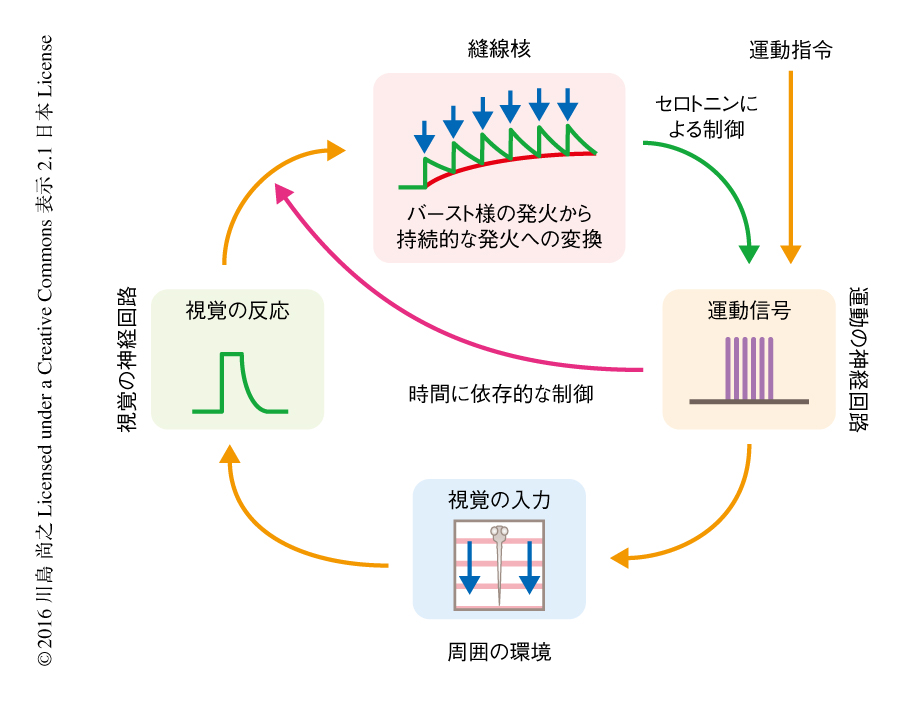
これらの結果から,縫線核が運動学習をつかさどる機構の全容がみえてきた.周囲の環境に対する運動のゲインが高くなり弱い運動出力で長距離を移動できる条件におかれた場合,自らの泳ぎに付随する周囲の環境の動きをニューロンのバースト様の発火によりコードする.そして,くり返しの学習により一連のニューロンのバースト様の発火を持続的な発火に変換して蓄積し,泳ぎが停止した期間もニューロンの持続的な発火を維持することにより直前の学習の情報を保持する.縫線核の神経活動が高まるとセロトニン性の伝達により泳ぎの強度が抑制されるため,弱い運動出力の記憶が保持される(図2).
運動学習において全脳Ca2+イメージング法による解析をするたびに,いつも特異な神経活動を示すニューロンの名前がわからず首をかしげていたときからこの研究はスタートした.責任領域が縫線核であると同定されたのちにも,この情動をつかさどるセロトニン系のニューロンにほんとうに運動学習の機構があるのか懐疑的な考え方はしばらくはなれなかったが,電気生理学的な実験により縫線核のニューロンに速い計算原理のあることが確認されたあたりから,ようやくこれが新規性のある研究であるとの認識が深まったことを覚えている.
これまで,セロトニン系の神経回路は情動行動や精神疾患において研究の対象になってきたが,今回の研究は,運動学習というまったく別の視点を提供した.当然,従来の研究との関連性を説明する必要が生じるが,セロトニン性の神経回路に関して提唱されている機能はあまりに多く,そのすべてをこの研究の結果と関連づけて説明することはできないし,矛盾するようにみえる研究も多い.ゼブラフィッシュは情動行動や精神疾患のモデル動物としても研究が進んでいることから,ゼブラフィッシュの系において縫線核の多様な側面を統合するモデルを構築していきたいと考えている.
略歴:2013年 東京大学大学院医学系研究科博士課程 修了,同年より米国Howard Hughes Medical Institute研究員.
研究テーマ:運動学習にかかわる神経回路の大規模イメージング法による解析.
関心事:音楽,テニス,ジョギング.留学中に料理の腕もあがりました.
© 2016 川島 尚之 Licensed under CC 表示 2.1 日本
(米国Howard Hughes Medical Institute,Janelia Research Campus)
email:川島尚之
DOI: 10.7875/first.author.2016.118
The serotonergic system tracks the outcomes of actions to mediate short-term motor learning.
Takashi Kawashima, Maarten F. Zwart, Chao-Tsung Yang, Brett D. Mensh, Misha B. Ahrens
Cell, 167, 933-946.e20 (2016)
要 約
外部の環境がたえず変動するなかでもつねに意図したとおりの動作ができるよう,動物の運動は学習の機構によりたえず制御される.これまで,運動の際にセロトニン性の神経回路が活性化することは知られていたが,具体的にどのように運動の制御に関与するのかは不明であった.筆者らは,この研究において,ゼブラフィッシュの全脳Ca2+イメージング法により縫線核のセロトニン系の神経回路が短期の運動学習において中心的な役割を担うことを発見した.さらに詳細な解析により,縫線核のニューロンには運動指令と同期した視覚の刺激への選択的な反応,および,くり返しの学習による持続的なニューロンの発火の蓄積,という2種類の計算原理の存在することが見い出された.この研究による運動学習のシンプルな理解から,セロトニン系の神経回路の多様な機能に共通する原理の解明につながることが期待される.
はじめに
縫線核に代表されるセロトニン系の神経回路は報酬学習や恐怖記憶といった幅広い情動行動にかかわることが知られ,うつ病に代表される精神疾患においても注目されている.これまで,セロトニンの伝達を制御する治療薬を中心に薬理学的な行動解析は幅広く行われてきたものの,セロトニン系の神経回路における神経機構に着目した研究は少なく,その幅広い役割に共通する原理は未解明なままである1).その理由のひとつとして,情動行動や精神疾患にかかわる行動の要素は複雑であり,単純な要素に分解した解析が困難なことがあげられる.
運動学習は動物の行動においてもっとも基本的な学習形態のひとつであり,脳からの運動指令の出力ののち,1)動作の結果を感知し,2)その情報を保存し,3)つぎの運動指令を修正する,という過程のくり返しにより担われる.これまでの長期の運動学習の研究により,この過程が小脳や大脳皮質の運動野のシナプス可塑性により担われることは解明されたものの,数分以内に起こるような短期の運動学習の機構についてはあまり知られていない.また,動物の運動の際に縫線核のニューロンが活性化することは知られていたものの2),その運動制御における具体的な役割は不明であった.
この研究においては,短期の運動学習の機構の解明をめざし,仮想環境においたゼブラフィッシュに対し全脳Ca2+イメージング法を適用した.
1.短期の運動学習におけるパラダイムの確立
短期の運動学習における責任神経回路を同定するため,ゼブラフィッシュの稚魚に対し仮想環境を用いて運動学習のパラダイムを構築した.この仮想環境において,ゼブラフィッシュの動きを固定するため筋肉を麻痺させ,筋肉へと伸びる脊髄運動ニューロンの軸索から電気生理学的に運動シグナルを記録した.ゼブラフィッシュはその下に投影された仮想環境を見ることができる.この仮想環境を前方に動かすと,ゼブラフィッシュはこれを本能的に追従しようとして運動シグナルを出力する.運動シグナルが記録されると仮想環境は後方に動き,ゼブラフィッシュはこれを見て自分が前進したと錯覚する3,4).この仮想環境においてゼブラフィッシュは運動学習を行う.水中においては水の温度や粘性の変化により意図した泳ぎの強さと実際の移動距離とにずれが生じる.このとき,泳ぎの強さに対して得られた移動距離をゲインといい,これに応じてゼブラフィッシュは運動の強さを制御する.仮想環境においては,記録された運動シグナルと仮想環境の動きとのあいだのゲインの値を操作することにより,ゼブラフィッシュの運動出力の強度を変化させることができる.ゲインが低いとゼブラフィッシュは前進するため強い運動シグナルを出力する必要がある.逆に,ゲインが高いとゼブラフィッシュは弱い運動シグナルでも十分に前進することができる.ゲインを継続的に高くする訓練を行えば,ゼブラフィッシュは運動学習により弱い運動シグナルを継続して出力するようになり,この弱い運動シグナルの出力は,たとえば,10秒間にわたり運動を停止させたのち,もういちど仮想環境を提示しても継続される.これが,この研究における“短期の運動学習”である.
学習の程度が訓練する時間の長さに比例することに注目して行動のパラダイムを開発した.このパラダイムは“開始期間”“訓練期間”“停止期間”“試験期間”の4つの期間から構成される(図1).開始期間において仮想環境のゲインは低く設定され,ゼブラフィッシュは強い運動シグナルを継続的に出力する.訓練期間において仮想環境のゲインは高く設定され,ゼブラフィッシュは短い時間(7秒)あるいは長い時間(30秒)にわたり弱い運動シグナルを継続的に出力する.停止期間において仮想環境は停止し,ゼブラフィッシュの運動も停止する.試験期間において仮想環境はもういちど前方に動き,それに応答したゼブラフィッシュの運動シグナルの強度を測定する.その結果,長い訓練を行った場合には,短い訓練の場合と比較して試験期間においてより弱い運動シグナルが記録された(図1).訓練中の運動シグナルの強度は時間の長さに関係なく一定であるため,脳にはくり返しの回数に依存して学習の効果が蓄積するような過程の存在することが示唆された.さらには,試験期間におけるゼブラフィッシュの反応時間や反応率は訓練の長さに対し不変であったことから,この学習はゼブラフィッシュの感覚応答性や覚醒状態には影響せず運動出力に特異的に作用することが示された.
2.全脳Ca2+イメージング法による責任領域の探索
この行動の責任領域を探索するため,運動学習中の脳における神経活動をライトシート顕微鏡により記録した.すべてのニューロンにCa2+指示タンパク質であるGCaMP6 5) を発現させた遺伝子改変ゼブラフィッシュに対し,2方向から励起光を照射することにより仮想環境において行動しているあいだ,全脳の8割以上,約10万個ものニューロンの神経活動が1~2 Hzで同時記録された6).
訓練中に蓄積された学習の情報を停止期間をこえて保持するニューロンを探索したところ,脳幹の前部に強い集積がみられた.過去の文献および免疫染色法により,これはセロトニン性の背側縫線核であると同定された.縫線核のセロトニン性のニューロンは訓練期間において時間に依存的に神経活動を蓄積し,停止期間において蓄積された神経活動が緩徐に減衰し,蓄積された学習の情報は試験期間まで保持された.停止期間における縫線核の神経活動が高いほど,つぎの試験期間における運動出力の強度は抑制された.
背側縫線核の運動学習への関与についてさらに調べるため,その神経活動を操作した.薬理学的な手法によりセロトニンの伝達を遮断する,あるいは,化学遺伝学的な手法によりセロトニン性のニューロンを選択的に細胞死させることにより,ゼブラフィッシュの短期の学習行動は完全に阻害された.また,光遺伝学的な手法により停止期間において背側縫線核の神経活動を上昇させると短期の学習行動は強化された.これらの結果から,背側縫線核の神経活動が短期の運動学習において中心的な役割をはたすことが示された.
3.電気生理学的な手法および高速イメージング法による縫線核のニューロンの計算原理の解明
縫線核のニューロンは運動学習中にどのような計算を行うのであろうか? その詳細な神経活動について明らかにするため,運動学習中のゼブラフィッシュの縫線核のニューロンから2光子顕微鏡による観察のもと細胞外記録を行った.ゼブラフィッシュの小さな脳に細胞外電極を挿入すると大半は行動をとめてしまうためこの実験は困難をきわめたが,少数ながら学習行動を示すゼブラフィッシュから記録を行った.その結果,縫線核のニューロンは訓練期間において個別の泳ぎの直後にバースト様の発火を示すこと,そして,くり返しの訓練が進むにつれ泳ぎに同期しない持続的な発火が蓄積することがわかった.また,このニューロンの持続的な発火は停止期間においても持続した.このニューロンの持続的な発火の蓄積は報酬学習の際にも起こることが報告されており7),セロトニン系の幅広い機能に共通することが示唆される.
では,個別の泳ぎの直後のニューロンのバースト様の発火はどのような情報をコードするのであろうか? コードされる情報は運動の強度であろうか,それとも,運動の直後の仮想環境の動きであろうか? このニューロンのバースト様の発火は高速のCa2+イメージング法によっても個別の運動出力の直後に観察されたため,細胞の大規模な解析が可能なCa2+イメージング法により解析した.仮想環境の条件をくふうすることにより,運動出力の強度を同じにしたまま,仮想環境の動きがゲインに比例して速くなる条件を構築した.この場合,縫線核のニューロンのバースト様の発火は仮想環境の動きの速さに比例して増大した.逆に,異なる運動出力の強度に対し仮想環境の動く速度が変わらない条件を設定すると,縫線核のニューロンのバースト様の発火は変化しなかった.このことから,縫線核のセロトニン性のニューロンのバースト様の発火は運動出力に付随して起こる仮想環境の動きをコードすることがわかった.
しかしながら,ここでひとつの疑問が浮上した.ゼブラフィッシュは自らの動きにより起こった周囲の環境の動きと,外部の力により動いた周囲の環境の動きの両方を見ることができる.縫線核において正確な運動学習を成立させるためには,自らの動きに付随する視覚入力を選択的に学習する必要がある.この疑問に答えるため,仮想環境の動きが運動出力と非同期になる状態をつくり,仮想環境の動きがゼブラフィッシュの泳ぎと同時に起こった場合と時間的にずれた場合との縫線核のニューロンのバースト様の発火を比較した.その結果,縫線核は周囲の環境の動きがゼブラフィッシュの運動出力と同期した場合にのみ反応することが示された(図2).
4.縫線核における運動学習モデルの構築
これらの結果から,縫線核が運動学習をつかさどる機構の全容がみえてきた.周囲の環境に対する運動のゲインが高くなり弱い運動出力で長距離を移動できる条件におかれた場合,自らの泳ぎに付随する周囲の環境の動きをニューロンのバースト様の発火によりコードする.そして,くり返しの学習により一連のニューロンのバースト様の発火を持続的な発火に変換して蓄積し,泳ぎが停止した期間もニューロンの持続的な発火を維持することにより直前の学習の情報を保持する.縫線核の神経活動が高まるとセロトニン性の伝達により泳ぎの強度が抑制されるため,弱い運動出力の記憶が保持される(図2).
おわりに
運動学習において全脳Ca2+イメージング法による解析をするたびに,いつも特異な神経活動を示すニューロンの名前がわからず首をかしげていたときからこの研究はスタートした.責任領域が縫線核であると同定されたのちにも,この情動をつかさどるセロトニン系のニューロンにほんとうに運動学習の機構があるのか懐疑的な考え方はしばらくはなれなかったが,電気生理学的な実験により縫線核のニューロンに速い計算原理のあることが確認されたあたりから,ようやくこれが新規性のある研究であるとの認識が深まったことを覚えている.
これまで,セロトニン系の神経回路は情動行動や精神疾患において研究の対象になってきたが,今回の研究は,運動学習というまったく別の視点を提供した.当然,従来の研究との関連性を説明する必要が生じるが,セロトニン性の神経回路に関して提唱されている機能はあまりに多く,そのすべてをこの研究の結果と関連づけて説明することはできないし,矛盾するようにみえる研究も多い.ゼブラフィッシュは情動行動や精神疾患のモデル動物としても研究が進んでいることから,ゼブラフィッシュの系において縫線核の多様な側面を統合するモデルを構築していきたいと考えている.
文 献
- Dayan, P. & Huys, Q.: Serotonin’s many meanings elude simple theories. Elife, 4, e07390 (2015)[PubMed]
- Jacobs, B. L. & Fornal, C. A.: Serotonin and motor activity. Curr. Opin. Neurobiol., 7, 820-825 (1997)[PubMed]
- Ahrens, M. B., Li, J. M., Orger, M. B. et al.: Brain-wide neuronal dynamics during motor adaptation in zebrafish. Nature, 485, 471-477 (2012)[PubMed]
- Vladimirov, N., Mu, Y., Kawashima, T. et al.: Light-sheet functional imaging in fictively behaving zebrafish. Nat. Methods, 11, 883-884 (2014)[PubMed]
- Chen, T. W., Wardill, T. J., Sun, Y. et al.: Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499, 295-300 (2013)[PubMed]
- Ahrens, M. B., Orger, M. B., Robson, D. N. et al.: Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat. Methods, 10, 413-420 (2013)[PubMed]
- Cohen, J. Y., Amoroso, M. W. & Uchida, N.: Serotonergic neurons signal reward and punishment on multiple timescales. Elife, 4, e06346 (2015)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2013年 東京大学大学院医学系研究科博士課程 修了,同年より米国Howard Hughes Medical Institute研究員.
研究テーマ:運動学習にかかわる神経回路の大規模イメージング法による解析.
関心事:音楽,テニス,ジョギング.留学中に料理の腕もあがりました.
© 2016 川島 尚之 Licensed under CC 表示 2.1 日本
