C9orf72遺伝子のもつ異常なくり返し配列から産生されるPRポリペプチドはLCドメインから形成されるポリマーと結合する
森 英一朗・加藤昌人
(米国Texas大学Southwestern Medical Center,Department of Biochemistry)
email:森 英一朗,加藤昌人
DOI: 10.7875/first.author.2016.112
Toxic PR poly-dipeptides encoded by the C9orf72 repeat expansion target LC domain polymers.
Yi Lin, Eiichiro Mori, Masato Kato, Siheng Xiang, Leeju Wu, Ilmin Kwon, Steven L. McKnight
Cell, 167, 789-802.e12 (2016)
筋萎縮性側索硬化症の主要な原因遺伝子のひとつであるC9orf72遺伝子のもつ異常なくり返し配列から産生される,細胞毒性のあるPRポリペプチドの細胞における標的タンパク質を2つの網羅的な手法により探索した.2つの方法に共通して検出されたPRポリペプチドとの結合タンパク質は多岐にわたったが,その多くは膜をもたないオルガネラに存在するタンパク質,および,中間径フィラメントを形成するタンパク質であった.PRポリペプチドの結合タンパク質の多くはLCドメインをもち,PRポリペプチドはポリマーの状態にあるLCドメインと結合していた.これらのことから,PRポリペプチドによる細胞毒性は,細胞構造の動態や遺伝子の転写あるいは翻訳の経路など,多岐にわたる機能の障害によりもたらされることが示唆された.
家族性の筋萎縮性側索硬化症の原因のうちもっとも頻度の高いものとして,C9orf72遺伝子の最初のイントロンにおけるGGGGCCからなるくり返し配列の異常な増幅が知られている1,2).健常者におけるくり返しの回数はごくわずかだが,患者においてはくり返しの回数が非常に多くなっている.このくり返し配列の増幅により筋萎縮性側索硬化症が発症する機序については,遺伝子の機能の抑制,転写産物による毒性,開始コドンに非依存的な翻訳による毒性ポリペプチドの産生,などさまざまなモデルが提唱されている.このくり返し配列からは合計で5種類のポリペプチドが産生されるが,筆者らは,なかでもGlyとArgからなるGRポリペプチドおよびProとArgからなるPRポリペプチドが強い細胞毒性をもつこと,そして,GRポリペプチドおよびPRポリペプチドはRNAの代謝を傷害することを明らかにしてきた3).しかしながら,GRポリペプチドあるいはPRポリペプチドの細胞における標的タンパク質は同定されていなかった.そこで,PRポリペプチドの細胞における標的タンパク質を2つの網羅的な手法により探索した.
PRポリペプチドの結合タンパク質の探索に2つの網羅的な手法を用いた.ひとつは,紫外線の照射により架橋を起こすようにした20回のくり返しをもつPRペプチドを培地にくわえて結合タンパク質を検出した.もうひとつは,タグを付加した100回のくり返しをもつPRペプチドを発現する細胞において結合タンパク質を検出した.これら2つの手法により検出されたタンパク質は高頻度で重複した.また,PRポリペプチドとの結合タンパク質の多くは膜をもたないオルガネラに存在するタンパク質,および,中間径フィラメントを形成するタンパク質であった.RNAの代謝にかかわるタンパク質が標的であることは,すでに筆者らの過去の報告3) からも想定されていたが,中間径フィラメントを形成するタンパク質が標的であることは予想外であった.これらのほかにも,核膜孔,核小体,核ラミナなどに存在するタンパク質が標的として検出された.同様の結果は,ほかの研究グループからもCell誌の同じ号に報告された4).
PRポリペプチドの結合タンパク質の多くはLC(Low-complexity)ドメインをもっていた.ヒトのプロテオームの全体にしめるLCドメインをもつタンパク質の比率は約10%であるのに対し,2つの手法において共通して検出されたタンパク質のLCドメインをもつ比率は約22%であった.このことから,PRポリペプチドはLCドメインをもつタンパク質と選択的に結合することが示唆された.実際に,LCドメインをもつhnRNPA1の組換え体を用いることにより,PRポリペプチドとの結合にはLCドメインが必須であることが確認された.
RNA結合タンパク質のもつLCドメインは生理的な条件においてアミロイド様のポリマーを形成する.LCドメインからなるポリマーは,アミロイド線維と同様にクロスβ構造をもつが,不可逆性のアミロイド線維とは異なり不安定で解離しやすい.筆者らは,これらのLCドメインからなる不安定なポリマーが膜をもたないオルガネラを構築するうえで機能する可能性を報告してきた5,6)(文献5) は新着論文レビュー でも掲載).
脂肪族アルコールである1,6-ヘキサンジオールは核膜孔やRNA顆粒などを溶解することが以前より知られている.これらにはLCドメインをもつタンパク質が多く存在する.実際に,さまざまな脂肪族アルコールを用いて比較したところ,FUSのLCドメインからなるポリマーのヒドロゲルは1,6-ヘキサンジオールにより溶解したが2,5-ヘキサンジオールでは溶解しなかった.しかしながら,病原性アミロイドであるアミロイドβタンパク質からなるポリマーはどの脂肪族アルコールによっても溶解しなかった.また,hnRNPA2やFUSのLCドメインからなる液体様の小滴も同様に,1,6-ヘキサンジオールにより選択的に溶解した.
細胞においてオルガネラが1,6-ヘキサンジオールにより溶解するか調べたところ,ストレス顆粒,Cajal小体,核顆粒は1,6-ヘキサンジオールにより溶解したが2,5-ヘキサンジオールでは溶解しなかった.さらに,アクチンフィラメントおよび微小管はどの脂肪族アルコールによっても溶解しなかったが,ケラチンあるいはビメンチンから形成された中間径フィラメントは1,6-ヘキサンジオールにより選択的に溶解した.1,6-ヘキサンジオールによる選択的な溶解から,これらの構造物はヒドロゲルと同様にクロスβポリマーの相互作用を介して形成されることが強く示唆された.
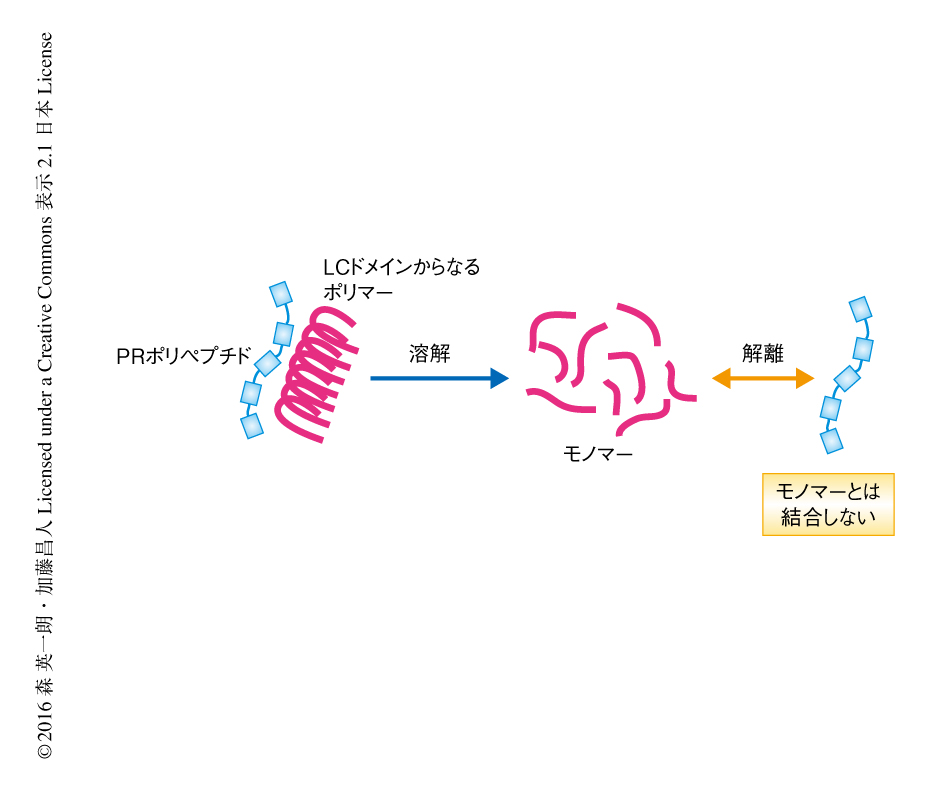
以前に筆者らは,PRポリペプチドはLCドメインからなるポリマーと結合することを報告したが3),PRポリペプチドがLCドメインからなるポリマーとのみ結合しているか,あるいは,モノマーとも結合するかは明らかではなかった.hnRNPA2のLCドメインからなるポリマーとPRポリペプチドとを結合させ1,6-ヘキサンジオールにより処理するとポリマーは溶解し,同時に,PRポリペプチドとの結合が消失した(図1).同様に,100回のくり返しをもつPRポリペプチドを発現させた細胞を1,6-ヘキサンジオールにより処理すると,PRポリペプチドと結合して精製されるタンパク質の多くが消失した.このことから,PRポリペプチドはLCドメインがポリマーの状態である場合にのみ結合すると考えられた.
hnRNPA2のLCドメインにおいて筋萎縮性側索硬化症などの神経変性疾患の患者からみつかったAsp290がValに置換した変異体は,ポリマーを形成しやすいことが知られている.一方,筆者らは,Phe291がSerに置換した変異体はポリマーの形成能がいちじるしく低いことを明らかにした7).これら2つのLCドメインの変異体を野生型のドメインと比較したところ,Asp290がValに置換した変異体は野生型と同じ程度にPRポリペプチドと結合したが,Phe291がSerに置換した変異体はPRポリペプチドと結合しなかった.このことからも,LCドメインのポリマーの形成がPRポリペプチドとの結合に必要であることが示された.
PRポリペプチドの標的として同定された中間径フィラメントを形成するタンパク質は,中央部にコイルドコイル二量体の形成に重要であるロッドドメインを,N末端側の領域およびC末端側の領域にLCドメインであるヘッドドメインおよびテイルドメインをもつ.これまで,中間径フィラメントの形成にはヘッドドメインが不可欠であることがわかっていたが,どのような機構によりはたらくのかはまったくわかっていなかった.中間径フィラメントを形成するいくつかのタンパク質のLCドメインを精製して調べたところ,調べたすべてのヘッドドメインはクロスβポリマーを形成することがわかった.ヘッドドメインからなるポリマーは,ほかのLCドメインからなるポリマーと同様に1,6-ヘキサンジオールにより選択的に溶解した.1,6-ヘキサンジオールにより処理することにより細胞においてビメンチンからなる中間径フィラメントが溶解したが,ビメンチンの組換え体から形成された中間径フィラメントも同様に1,6-ヘキサンジオールにより選択的に溶解した.これらのことより,ヘッドドメインにおけるクロスβポリマーの形成が中間径フィラメントの形成の過程において重要なステップである可能性が示唆された.PRポリペプチドはヘッドドメインからなるポリマーのヒドロゲルと結合した.さらに,PRポリペプチドをビメンチンの組換え体から形成された中間径フィラメントと混ぜると,中間径フィラメントにしずくのように結合するようすが観察された.しずくのあいだの間隔は中間径フィラメントの基本単位の長さと同じ47 nmであった.中間径フィラメントには47 nmごとにヘッドドメインとテイルドメインとのジャンクションがあることから,PRポリペプチドはジャンクションにあるヘッドドメインからなるポリマーを認識して結合することが示唆された.
この研究により,PRポリペプチドの細胞における標的はポリマーの状態のLCドメインであることが明らかにされた.この結果は,筆者らが以前より提唱している,RNA結合タンパク質などのもつLCドメインは細胞においてふだんからポリマーの状態として機能するという考えと合致した.PRポリペプチドの細胞毒性は,PRポリペプチドとの結合によりLCドメインからなるポリマーの安定性に変化が生じ通常の機能がさまたげられることによりひき起こされると考えられた.今回の研究の成果が,さまざまな神経変性疾患において発症の機序の理解につながることが期待される.
略歴:米国Texas大学Southwestern Medical Center研究員.
研究テーマ:LCドメインの構造および機能.
加藤 昌人(Masato Kato)
米国Texas大学Southwestern Medical CenterにてAssociate Professor.
© 2016 森 英一朗・加藤昌人 Licensed under CC 表示 2.1 日本
(米国Texas大学Southwestern Medical Center,Department of Biochemistry)
email:森 英一朗,加藤昌人
DOI: 10.7875/first.author.2016.112
Toxic PR poly-dipeptides encoded by the C9orf72 repeat expansion target LC domain polymers.
Yi Lin, Eiichiro Mori, Masato Kato, Siheng Xiang, Leeju Wu, Ilmin Kwon, Steven L. McKnight
Cell, 167, 789-802.e12 (2016)
要 約
筋萎縮性側索硬化症の主要な原因遺伝子のひとつであるC9orf72遺伝子のもつ異常なくり返し配列から産生される,細胞毒性のあるPRポリペプチドの細胞における標的タンパク質を2つの網羅的な手法により探索した.2つの方法に共通して検出されたPRポリペプチドとの結合タンパク質は多岐にわたったが,その多くは膜をもたないオルガネラに存在するタンパク質,および,中間径フィラメントを形成するタンパク質であった.PRポリペプチドの結合タンパク質の多くはLCドメインをもち,PRポリペプチドはポリマーの状態にあるLCドメインと結合していた.これらのことから,PRポリペプチドによる細胞毒性は,細胞構造の動態や遺伝子の転写あるいは翻訳の経路など,多岐にわたる機能の障害によりもたらされることが示唆された.
はじめに
家族性の筋萎縮性側索硬化症の原因のうちもっとも頻度の高いものとして,C9orf72遺伝子の最初のイントロンにおけるGGGGCCからなるくり返し配列の異常な増幅が知られている1,2).健常者におけるくり返しの回数はごくわずかだが,患者においてはくり返しの回数が非常に多くなっている.このくり返し配列の増幅により筋萎縮性側索硬化症が発症する機序については,遺伝子の機能の抑制,転写産物による毒性,開始コドンに非依存的な翻訳による毒性ポリペプチドの産生,などさまざまなモデルが提唱されている.このくり返し配列からは合計で5種類のポリペプチドが産生されるが,筆者らは,なかでもGlyとArgからなるGRポリペプチドおよびProとArgからなるPRポリペプチドが強い細胞毒性をもつこと,そして,GRポリペプチドおよびPRポリペプチドはRNAの代謝を傷害することを明らかにしてきた3).しかしながら,GRポリペプチドあるいはPRポリペプチドの細胞における標的タンパク質は同定されていなかった.そこで,PRポリペプチドの細胞における標的タンパク質を2つの網羅的な手法により探索した.
1.PRポリペプチドはLCドメインと結合する
PRポリペプチドの結合タンパク質の探索に2つの網羅的な手法を用いた.ひとつは,紫外線の照射により架橋を起こすようにした20回のくり返しをもつPRペプチドを培地にくわえて結合タンパク質を検出した.もうひとつは,タグを付加した100回のくり返しをもつPRペプチドを発現する細胞において結合タンパク質を検出した.これら2つの手法により検出されたタンパク質は高頻度で重複した.また,PRポリペプチドとの結合タンパク質の多くは膜をもたないオルガネラに存在するタンパク質,および,中間径フィラメントを形成するタンパク質であった.RNAの代謝にかかわるタンパク質が標的であることは,すでに筆者らの過去の報告3) からも想定されていたが,中間径フィラメントを形成するタンパク質が標的であることは予想外であった.これらのほかにも,核膜孔,核小体,核ラミナなどに存在するタンパク質が標的として検出された.同様の結果は,ほかの研究グループからもCell誌の同じ号に報告された4).
PRポリペプチドの結合タンパク質の多くはLC(Low-complexity)ドメインをもっていた.ヒトのプロテオームの全体にしめるLCドメインをもつタンパク質の比率は約10%であるのに対し,2つの手法において共通して検出されたタンパク質のLCドメインをもつ比率は約22%であった.このことから,PRポリペプチドはLCドメインをもつタンパク質と選択的に結合することが示唆された.実際に,LCドメインをもつhnRNPA1の組換え体を用いることにより,PRポリペプチドとの結合にはLCドメインが必須であることが確認された.
2.脂肪族アルコールによる細胞内の構造物の溶解
RNA結合タンパク質のもつLCドメインは生理的な条件においてアミロイド様のポリマーを形成する.LCドメインからなるポリマーは,アミロイド線維と同様にクロスβ構造をもつが,不可逆性のアミロイド線維とは異なり不安定で解離しやすい.筆者らは,これらのLCドメインからなる不安定なポリマーが膜をもたないオルガネラを構築するうえで機能する可能性を報告してきた5,6)(文献5) は新着論文レビュー でも掲載).
脂肪族アルコールである1,6-ヘキサンジオールは核膜孔やRNA顆粒などを溶解することが以前より知られている.これらにはLCドメインをもつタンパク質が多く存在する.実際に,さまざまな脂肪族アルコールを用いて比較したところ,FUSのLCドメインからなるポリマーのヒドロゲルは1,6-ヘキサンジオールにより溶解したが2,5-ヘキサンジオールでは溶解しなかった.しかしながら,病原性アミロイドであるアミロイドβタンパク質からなるポリマーはどの脂肪族アルコールによっても溶解しなかった.また,hnRNPA2やFUSのLCドメインからなる液体様の小滴も同様に,1,6-ヘキサンジオールにより選択的に溶解した.
細胞においてオルガネラが1,6-ヘキサンジオールにより溶解するか調べたところ,ストレス顆粒,Cajal小体,核顆粒は1,6-ヘキサンジオールにより溶解したが2,5-ヘキサンジオールでは溶解しなかった.さらに,アクチンフィラメントおよび微小管はどの脂肪族アルコールによっても溶解しなかったが,ケラチンあるいはビメンチンから形成された中間径フィラメントは1,6-ヘキサンジオールにより選択的に溶解した.1,6-ヘキサンジオールによる選択的な溶解から,これらの構造物はヒドロゲルと同様にクロスβポリマーの相互作用を介して形成されることが強く示唆された.
3.PRポリペプチドはhnRNPA2のLCドメインからなるポリマーと結合する
以前に筆者らは,PRポリペプチドはLCドメインからなるポリマーと結合することを報告したが3),PRポリペプチドがLCドメインからなるポリマーとのみ結合しているか,あるいは,モノマーとも結合するかは明らかではなかった.hnRNPA2のLCドメインからなるポリマーとPRポリペプチドとを結合させ1,6-ヘキサンジオールにより処理するとポリマーは溶解し,同時に,PRポリペプチドとの結合が消失した(図1).同様に,100回のくり返しをもつPRポリペプチドを発現させた細胞を1,6-ヘキサンジオールにより処理すると,PRポリペプチドと結合して精製されるタンパク質の多くが消失した.このことから,PRポリペプチドはLCドメインがポリマーの状態である場合にのみ結合すると考えられた.
hnRNPA2のLCドメインにおいて筋萎縮性側索硬化症などの神経変性疾患の患者からみつかったAsp290がValに置換した変異体は,ポリマーを形成しやすいことが知られている.一方,筆者らは,Phe291がSerに置換した変異体はポリマーの形成能がいちじるしく低いことを明らかにした7).これら2つのLCドメインの変異体を野生型のドメインと比較したところ,Asp290がValに置換した変異体は野生型と同じ程度にPRポリペプチドと結合したが,Phe291がSerに置換した変異体はPRポリペプチドと結合しなかった.このことからも,LCドメインのポリマーの形成がPRポリペプチドとの結合に必要であることが示された.
4.PRペプチドは中間径フィラメントを形成するタンパク質のN末端の領域に結合する
PRポリペプチドの標的として同定された中間径フィラメントを形成するタンパク質は,中央部にコイルドコイル二量体の形成に重要であるロッドドメインを,N末端側の領域およびC末端側の領域にLCドメインであるヘッドドメインおよびテイルドメインをもつ.これまで,中間径フィラメントの形成にはヘッドドメインが不可欠であることがわかっていたが,どのような機構によりはたらくのかはまったくわかっていなかった.中間径フィラメントを形成するいくつかのタンパク質のLCドメインを精製して調べたところ,調べたすべてのヘッドドメインはクロスβポリマーを形成することがわかった.ヘッドドメインからなるポリマーは,ほかのLCドメインからなるポリマーと同様に1,6-ヘキサンジオールにより選択的に溶解した.1,6-ヘキサンジオールにより処理することにより細胞においてビメンチンからなる中間径フィラメントが溶解したが,ビメンチンの組換え体から形成された中間径フィラメントも同様に1,6-ヘキサンジオールにより選択的に溶解した.これらのことより,ヘッドドメインにおけるクロスβポリマーの形成が中間径フィラメントの形成の過程において重要なステップである可能性が示唆された.PRポリペプチドはヘッドドメインからなるポリマーのヒドロゲルと結合した.さらに,PRポリペプチドをビメンチンの組換え体から形成された中間径フィラメントと混ぜると,中間径フィラメントにしずくのように結合するようすが観察された.しずくのあいだの間隔は中間径フィラメントの基本単位の長さと同じ47 nmであった.中間径フィラメントには47 nmごとにヘッドドメインとテイルドメインとのジャンクションがあることから,PRポリペプチドはジャンクションにあるヘッドドメインからなるポリマーを認識して結合することが示唆された.
おわりに
この研究により,PRポリペプチドの細胞における標的はポリマーの状態のLCドメインであることが明らかにされた.この結果は,筆者らが以前より提唱している,RNA結合タンパク質などのもつLCドメインは細胞においてふだんからポリマーの状態として機能するという考えと合致した.PRポリペプチドの細胞毒性は,PRポリペプチドとの結合によりLCドメインからなるポリマーの安定性に変化が生じ通常の機能がさまたげられることによりひき起こされると考えられた.今回の研究の成果が,さまざまな神経変性疾患において発症の機序の理解につながることが期待される.
文 献
- DeJesus-Hernandez, M., Mackenzie, I. R., Boeve, B. F. et al.: Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron, 72, 245-256 (2011)[PubMed]
- Renton, A. E., Majounie, E., Waite, A. et al.: A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron, 72, 257-268 (2011)[PubMed]
- Kwon, I., Xiang, S., Kato, M. et al.: Poly-dipeptides encoded by the C9orf72 repeats bind nucleoli, impede RNA biogenesis, and kill cells. Science, 345, 1139-1145 (2014)[PubMed]
- Lee, K. H., Zhang, P., Kim, H. J. et al.: C9orf72 dipeptide repeats impair the assembly, dynamics, and function of membrane-less organelles. Cell, 167, 774-788 (2016)[PubMed]
- Kato, M., Han, T. W., Xie, S. et al.: Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell, 149, 753-767 (2012)[PubMed] [新着論文レビュー]
- Han, T. W., Kato, M., Xie, S. et al.: Cell-free formation of RNA granules: bound RNAs identify features and components of cellular assemblies. Cell, 149, 768-779 (2012)[PubMed]
- Xiang, S., Kato, M., Wu, L. C. et al.: The LC domain of hnRNPA2 adopts similar conformations in hydrogel polymers, liquid-like droplets, and nuclei. Cell, 163, 829-839 (2015)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:米国Texas大学Southwestern Medical Center研究員.
研究テーマ:LCドメインの構造および機能.
加藤 昌人(Masato Kato)
米国Texas大学Southwestern Medical CenterにてAssociate Professor.
© 2016 森 英一朗・加藤昌人 Licensed under CC 表示 2.1 日本
