霊長類の側頭葉において記憶を想起の対象へと変換する第5層から第6層への層構造のモジュール
小谷野賢治・竹田真己・宮下保司
(東京大学大学院医学系研究科 統合生理学)
email:小谷野賢治,竹田真己,宮下保司
DOI: 10.7875/first.author.2016.106
Laminar module cascade from layer 5 to 6 implementing cue-to-target conversion for object memory retrieval in the primate temporal cortex.
Kenji W. Koyano, Masaki Takeda, Teppei Matsui, Toshiyuki Hirabayashi, Yohei Ohashi, Yasushi Miyashita
Neuron, 92, 518-529 (2016)
大脳皮質は6つの層からなる層構造により構成され,それぞれの層は異なる機能を分担しながら局所の神経回路により結合し情報を処理すると考えられている.しかしながら,従来の実験手法には方法論的な限界があったため,これまで,高次連合野において層構造にそって情報が処理されているかどうかはわかっていなかった.この研究においては,高磁場MRI法を用いることにより神経活動を記録したニューロンの層構造における位置を同定する手法を開発し,記憶想起課題を遂行しているサルの側頭葉において適用した.その結果,手がかりとなる刺激から想起の対象への変換は大脳皮質の深層において起こることが明らかにされた.想起の対象への変換は第5層においてはじまり,つづいて,第6層において観察された.第5層に存在するニューロンが手がかりとなる刺激と想起の対象とのあいだの連合記憶を符号化する一方,第6層のニューロンは想起された情報を出力しており,第5層から第6層へと情報が受けわたされるなかで想起の対象へと表象が変換される情報処理の機構が示された.
大脳皮質は6つの重なりあった層構造から構成され,それぞれの層は独自の細胞種の構成,層の内外との解剖学的な結合,脳の離れた領域との入出力のパターンをもつ.大脳皮質の層構造における情報処理のモデルとしては,大脳皮質に入力された情報は最初に第4層へ到達し,ついで浅層(第2層~第3層),最後に深層(第5層~第6層)において処理されるというカノニカル回路モデルが提唱されてきた1).初期の感覚野においては大脳皮質の層構造を単位とする感覚情報の符号化についてよく研究されており,カノニカル回路にそって感覚情報が漸進的に処理され,しだいに複雑な表現で符号化されるといわれている2).しかしながら,記憶や注意などの高次の認知機能を担う高次連合野においても,大脳皮質のおのおのの層が異なる機能を担いながら情報を処理するのかどうかはわかっていなかった.これまでのヒトおよび霊長類における一連の研究により,視覚性の長期記憶の想起には側頭部の連合野が深くかかわることが示されており3,4),なかでも,手がかりとなる刺激をもとに対象を想起する際には36野が重要な役割をはたすことがわかっている3).これまでに,36野においては層構造をまたいだ信号の流れがあることは調べられていたものの5),おのおのの層が想起のときにどのような機能をはたすのかについてはわかっていなかった.
このような問題を調べるには,神経活動を記録したニューロンがどの層に存在するのかを信頼できる手法により確かめることが必須である.従来,麻酔下の動物を用いた初期の感覚野の急性実験においては,微小電極を用いた記録ののち局所的に焼灼しその位置を標識する手法が用いられてきた.しかしながら,このような標識は短期間しか残存せず標識が可能な数もかぎられるため,課題を訓練した霊長類を慢性的に用いる高次の認知機能の研究には不向きであった.そこで,この研究においては,MRI(magnetic resonance imaging,核磁気共鳴画像)法を用いて生体において微小電極の先端の位置を正確に同定する手法6,7) を開発することにより,記憶の想起における大脳皮質の層構造の機能的な分化について調べた.
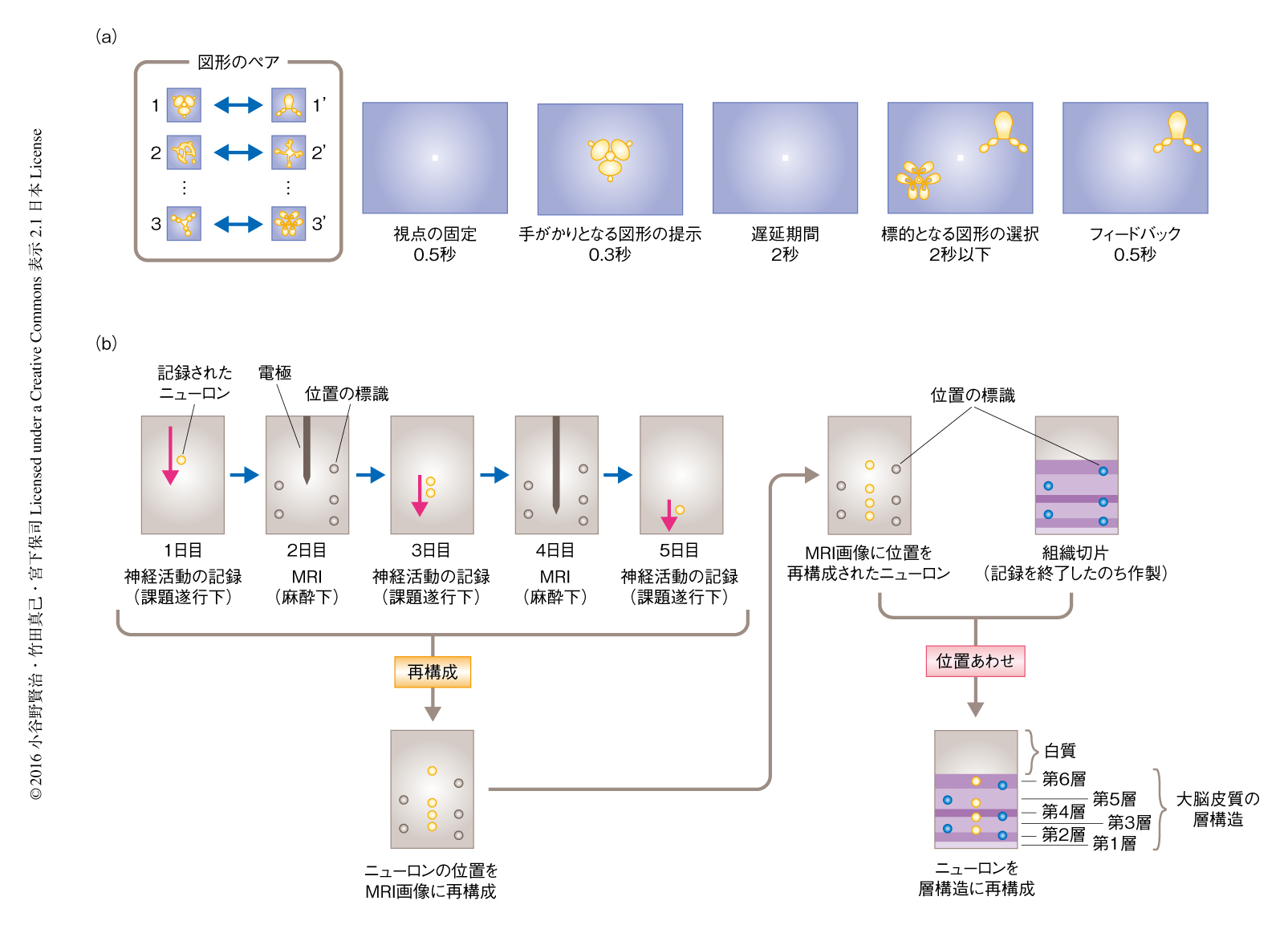
2頭のマカクザルに記憶想起課題を訓練した.この課題は標準的な記憶障害検査において使用されているものと同様である.複数の図形のペアを記憶するよう要求され,おのおのの試行において手がかりとなる図形からそれとペアを組むもう一方の図形を選択する(図1a).この課題を遂行しているサルから微小電極により単一ユニットの神経活動を記録した.微小電極の詳細な位置はその記録の最中に外部から直接に見ることはできない.そこで,ニューロンの位置を大脳皮質の層構造の解像度で同定するため,高磁場MRI法および組織切片法を併用する新たな手法を開発した(図1b).毎回の刺入において微小電極は半慢性的に留置され,刺入の途中にてMRI法によりその位置を同定した.微小電極の先端はMRI画像のボクセルの大きさよりもかなり小さいため,MRI画像の撮像に際しては微小電極の先端の信号を強調してその位置を検出できるよう調整した撮像パラメーター6) を用いた.おのおのの刺入において記録されたニューロンは,撮像されたMRI画像をもとにその位置を再構成した.しかしながら,MRI画像のみでは大脳皮質の層構造を判別するだけのコントラストを達成できない.そのため,層構造の同定には細胞種の構成の差の検出力にすぐれた組織切片法を用いた.MRI法を併用した一連のニューロンの記録をおえたのち組織切片を作製し,ニッスル染色法により記録された部位の周辺の層構造を同定した.MRI画像に再構成したニューロンの位置を,MRI法および組織切片法により描出の可能な位置の標識7) をもとに,この組織切片へと位置あわせすることにより大脳皮質の層構造における位置を同定した.この手法による再構成の誤差平均は層構造の大きさに比べ十分に小さく,シミュレーションによる試行においては,記録されたニューロンのうち86.1%は層への同定において再構成の誤差による影響はなかった.
第5層に位置するいくつかのニューロンにおいては,もっとも大きな応答をひき起こす“最適な”図形にくわえ,それとペアを組む図形に対しても持続的な応答を示すような反応が観察される傾向にあり,これらは学習した図形どうしの連合記憶を表象していると考えられた.一方,第6層に位置するニューロンにおいては,最適な図形とペアを組む図形を提示した際に,手がかりとなる図形の提示につづく遅延期間のなかほどから応答が上昇する傾向のみられるものがあり,これらは標的として想起された“最適な”図形の情報を表象していると考えられた.これら深層に位置するニューロンは想起に関連した神経活動を示したが,それに対し,浅層である第3層に位置するニューロンにおいては提示された手がかりとなる図形の情報を維持つづける神経活動がみられた.これらの反応パターンを定量化するため,提示された図形に対する表象,および,想起の対象となる図形に対する表象の度合いを定量化する指標を定義して,ニューロンごとにその時間的な変遷について解析した.
その結果,遅延期間において提示された図形に対する表象の指標はすべての層において有意に0よりも大きかったが,想起の対象となる図形に対する表象の指標は第5層および第6層において有意に0よりも大きく,かつ,ほかの層よりも高い値を示し,これら深層では想起された対象に対する表象が進んでいた.提示された図形に対する表象と想起の対象となる図形に対する表象との相対的な関係をこの2つの指標の差分により定量したところ,第2層~第4層においては提示された手がかりとなる図形の情報を表象し,第6層においては想起の対象となる図形の情報を表象していた.第5層においてはバランスがとれており,これは,提示された手がかりとなる図形および想起の対象となる図形を表象しているためと考えられた.
以上の結果から,深層を構成する第5層および第6層は遅延期間において想起の対象に対する情報を符号化することがわかったが,第5層および第6層は異なる細胞種から構築されていることから機能的にも差異があると思われた.実際に,遅延期間にさきだつ手がかりとなる図形の提示期間において,第5層においてはすでに想起の対象となる図形の情報を表象しはじめていたものの,第6層においては想起の対象となる図形に対する表象は観察されず,想起の対象となる図形の情報処理に関する時間的な差のあることが示唆された.そこで,想起の対象となる図形の情報を表象するようになるまでの潜時を定量し比較したところ,第5層においては第6層よりも早く想起の対象となる図形の情報を表象するようになることが確認された.第5層においては想起の対象に対し神経活動を示すニューロンの1/3が150 msよりも短い潜時をもち,これらはフィードフォワード処理8,9) により生成されることが示唆された.一方,提示された手がかりとなる図形の情報の表象に関しては,第6層においてはより短い期間しか保持されていなかった.
これら想起の対象となる図形および手がかりとなる図形に対する時間の経過をもとにクラスター解析により第5層および第6層のニューロンを分類したところ,第6層においては2つのクラスターに分割された.1つ目のクラスターである早期グループは,想起の対象となる図形の情報を早い段階から表象し,また,手がかりとなる図形の情報も相対的に長く保持し,結果として,想起の対象となる図形の情報および手がかりとなる図形の情報を表象するタイミングが重なりあう傾向にあった.2つ目のクラスターである後期グループは,想起の対象となる図形の情報をより遅いタイミングで表象しはじめ,手がかりとなる図形の情報を早い段階で失うため,想起の対象となる図形の情報および手がかりとなる図形の情報を表象するタイミングは重なりあわない傾向にあった.そのため,このグループのニューロンは遅延期間の終盤においては想起の対象となる図形の情報のみを表象していた.これら2つのグループは同じ第6層に存在するにもかかわらず異なる性質を示したことから,第6層における別々の神経基盤にもとづきはたらく可能性が考えられた.そこで,局所の神経活動の総体が反映される局所フィールド電位とおのおののニューロンの発火とのあいだのコヒーレンスを計算してその同期的な神経活動について調べた.その結果,後期グループにおいては遅延期間においてθ帯域における局所フィールド電位と同期した神経活動が上昇した.このことから,最終的に想起の対象となる図形の情報のみを表象するようになる後期グループはθ帯域において協調的に活動し,想起した情報を脳のほかの領域へと出力することが示唆された.
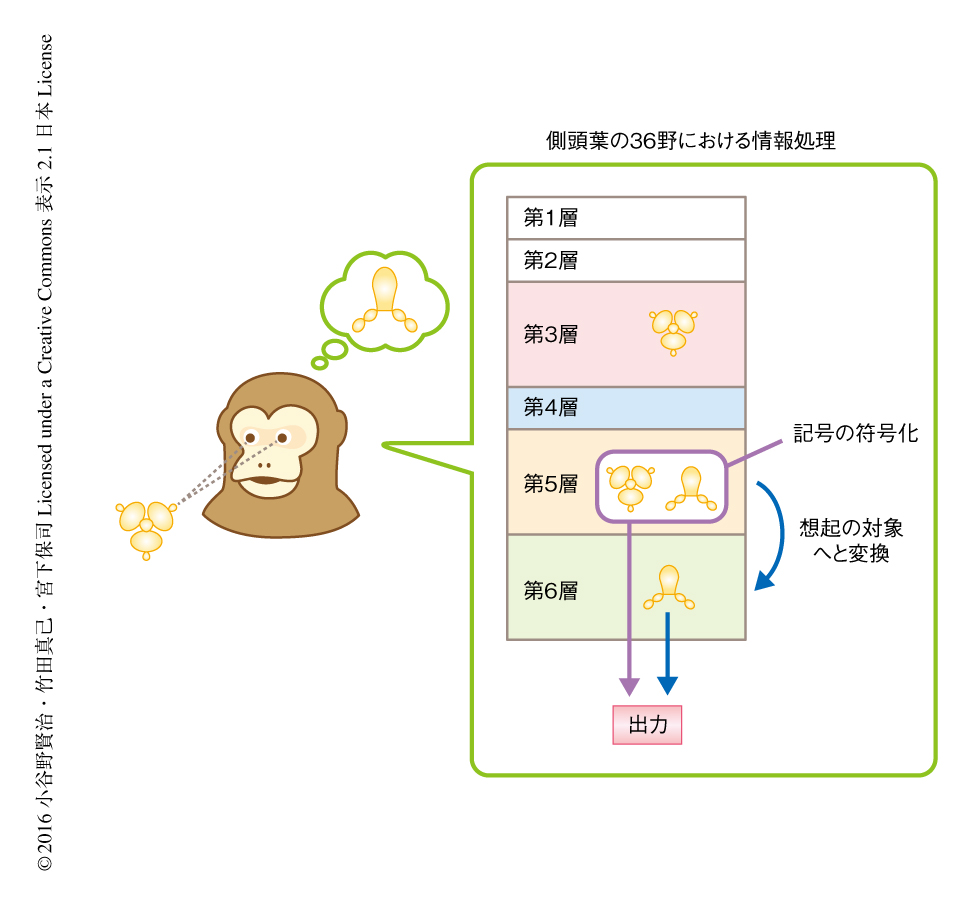
この研究により,霊長類では側頭葉の連合野である36野において,大脳皮質の第5層は記憶情報の連合に関与し,そこから想起の対象へと変換された情報を第6層が処理すること,また,想起の対象を処理する第6層の一部は互いに同期して活動しながら情報を脳のほかの領域に出力することが示唆された(図2).これらは,図形の複雑な情報が漸進的に処理されるという初期の視覚野における神経回路モデル2) とは異なる動作であり,側頭葉における記憶の神経回路にはそのための独自の機構が備わることが示された.初期の感覚野との神経回路の違いがどのように生じているか,また,ほかの高次の認知機能を担う連合野においてはどうなのか,今後の研究が待たれる.また,この研究において開発されたMRI法を併用したニューロンの位置の同定はほかの連合野における研究においても適用が可能であり,ほかのさまざまな高次の認知機能の情報処理においても大脳皮質の層構造にそった神経回路がはたす役割が明らかにされることが期待される.
略歴:2008年 東京大学大学院医学系研究科研究科博士課程 修了,同 研究員,同 助教を経て,2014年より米国NIH National Institute of Mental Health博士研究員.
研究テーマ:視覚の情報処理にかかわる高次連合野の神経回路.
抱負:知覚の心的な表象の機構を,錯視,注意,想起などの観点から明らかにしていきたい.
竹田 真己(Masaki Takeda)
順天堂大学大学院医学研究科 特任准教授.
宮下 保司(Yasushi Miyashita)
順天堂大学大学院医学研究科 特任教授.
研究室URL:http://www.physiol.m.u-tokyo.ac.jp/
© 2016 小谷野賢治・竹田真己・宮下保司 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 統合生理学)
email:小谷野賢治,竹田真己,宮下保司
DOI: 10.7875/first.author.2016.106
Laminar module cascade from layer 5 to 6 implementing cue-to-target conversion for object memory retrieval in the primate temporal cortex.
Kenji W. Koyano, Masaki Takeda, Teppei Matsui, Toshiyuki Hirabayashi, Yohei Ohashi, Yasushi Miyashita
Neuron, 92, 518-529 (2016)
要 約
大脳皮質は6つの層からなる層構造により構成され,それぞれの層は異なる機能を分担しながら局所の神経回路により結合し情報を処理すると考えられている.しかしながら,従来の実験手法には方法論的な限界があったため,これまで,高次連合野において層構造にそって情報が処理されているかどうかはわかっていなかった.この研究においては,高磁場MRI法を用いることにより神経活動を記録したニューロンの層構造における位置を同定する手法を開発し,記憶想起課題を遂行しているサルの側頭葉において適用した.その結果,手がかりとなる刺激から想起の対象への変換は大脳皮質の深層において起こることが明らかにされた.想起の対象への変換は第5層においてはじまり,つづいて,第6層において観察された.第5層に存在するニューロンが手がかりとなる刺激と想起の対象とのあいだの連合記憶を符号化する一方,第6層のニューロンは想起された情報を出力しており,第5層から第6層へと情報が受けわたされるなかで想起の対象へと表象が変換される情報処理の機構が示された.
はじめに
大脳皮質は6つの重なりあった層構造から構成され,それぞれの層は独自の細胞種の構成,層の内外との解剖学的な結合,脳の離れた領域との入出力のパターンをもつ.大脳皮質の層構造における情報処理のモデルとしては,大脳皮質に入力された情報は最初に第4層へ到達し,ついで浅層(第2層~第3層),最後に深層(第5層~第6層)において処理されるというカノニカル回路モデルが提唱されてきた1).初期の感覚野においては大脳皮質の層構造を単位とする感覚情報の符号化についてよく研究されており,カノニカル回路にそって感覚情報が漸進的に処理され,しだいに複雑な表現で符号化されるといわれている2).しかしながら,記憶や注意などの高次の認知機能を担う高次連合野においても,大脳皮質のおのおのの層が異なる機能を担いながら情報を処理するのかどうかはわかっていなかった.これまでのヒトおよび霊長類における一連の研究により,視覚性の長期記憶の想起には側頭部の連合野が深くかかわることが示されており3,4),なかでも,手がかりとなる刺激をもとに対象を想起する際には36野が重要な役割をはたすことがわかっている3).これまでに,36野においては層構造をまたいだ信号の流れがあることは調べられていたものの5),おのおのの層が想起のときにどのような機能をはたすのかについてはわかっていなかった.
このような問題を調べるには,神経活動を記録したニューロンがどの層に存在するのかを信頼できる手法により確かめることが必須である.従来,麻酔下の動物を用いた初期の感覚野の急性実験においては,微小電極を用いた記録ののち局所的に焼灼しその位置を標識する手法が用いられてきた.しかしながら,このような標識は短期間しか残存せず標識が可能な数もかぎられるため,課題を訓練した霊長類を慢性的に用いる高次の認知機能の研究には不向きであった.そこで,この研究においては,MRI(magnetic resonance imaging,核磁気共鳴画像)法を用いて生体において微小電極の先端の位置を正確に同定する手法6,7) を開発することにより,記憶の想起における大脳皮質の層構造の機能的な分化について調べた.
1.MRI法による神経活動を記録したニューロンの位置の同定
2頭のマカクザルに記憶想起課題を訓練した.この課題は標準的な記憶障害検査において使用されているものと同様である.複数の図形のペアを記憶するよう要求され,おのおのの試行において手がかりとなる図形からそれとペアを組むもう一方の図形を選択する(図1a).この課題を遂行しているサルから微小電極により単一ユニットの神経活動を記録した.微小電極の詳細な位置はその記録の最中に外部から直接に見ることはできない.そこで,ニューロンの位置を大脳皮質の層構造の解像度で同定するため,高磁場MRI法および組織切片法を併用する新たな手法を開発した(図1b).毎回の刺入において微小電極は半慢性的に留置され,刺入の途中にてMRI法によりその位置を同定した.微小電極の先端はMRI画像のボクセルの大きさよりもかなり小さいため,MRI画像の撮像に際しては微小電極の先端の信号を強調してその位置を検出できるよう調整した撮像パラメーター6) を用いた.おのおのの刺入において記録されたニューロンは,撮像されたMRI画像をもとにその位置を再構成した.しかしながら,MRI画像のみでは大脳皮質の層構造を判別するだけのコントラストを達成できない.そのため,層構造の同定には細胞種の構成の差の検出力にすぐれた組織切片法を用いた.MRI法を併用した一連のニューロンの記録をおえたのち組織切片を作製し,ニッスル染色法により記録された部位の周辺の層構造を同定した.MRI画像に再構成したニューロンの位置を,MRI法および組織切片法により描出の可能な位置の標識7) をもとに,この組織切片へと位置あわせすることにより大脳皮質の層構造における位置を同定した.この手法による再構成の誤差平均は層構造の大きさに比べ十分に小さく,シミュレーションによる試行においては,記録されたニューロンのうち86.1%は層への同定において再構成の誤差による影響はなかった.
2.おのおのの層のニューロンによる物体の符号化のダイナミクス
第5層に位置するいくつかのニューロンにおいては,もっとも大きな応答をひき起こす“最適な”図形にくわえ,それとペアを組む図形に対しても持続的な応答を示すような反応が観察される傾向にあり,これらは学習した図形どうしの連合記憶を表象していると考えられた.一方,第6層に位置するニューロンにおいては,最適な図形とペアを組む図形を提示した際に,手がかりとなる図形の提示につづく遅延期間のなかほどから応答が上昇する傾向のみられるものがあり,これらは標的として想起された“最適な”図形の情報を表象していると考えられた.これら深層に位置するニューロンは想起に関連した神経活動を示したが,それに対し,浅層である第3層に位置するニューロンにおいては提示された手がかりとなる図形の情報を維持つづける神経活動がみられた.これらの反応パターンを定量化するため,提示された図形に対する表象,および,想起の対象となる図形に対する表象の度合いを定量化する指標を定義して,ニューロンごとにその時間的な変遷について解析した.
その結果,遅延期間において提示された図形に対する表象の指標はすべての層において有意に0よりも大きかったが,想起の対象となる図形に対する表象の指標は第5層および第6層において有意に0よりも大きく,かつ,ほかの層よりも高い値を示し,これら深層では想起された対象に対する表象が進んでいた.提示された図形に対する表象と想起の対象となる図形に対する表象との相対的な関係をこの2つの指標の差分により定量したところ,第2層~第4層においては提示された手がかりとなる図形の情報を表象し,第6層においては想起の対象となる図形の情報を表象していた.第5層においてはバランスがとれており,これは,提示された手がかりとなる図形および想起の対象となる図形を表象しているためと考えられた.
3.第5層および第6層における想起の情報のダイナミクス
以上の結果から,深層を構成する第5層および第6層は遅延期間において想起の対象に対する情報を符号化することがわかったが,第5層および第6層は異なる細胞種から構築されていることから機能的にも差異があると思われた.実際に,遅延期間にさきだつ手がかりとなる図形の提示期間において,第5層においてはすでに想起の対象となる図形の情報を表象しはじめていたものの,第6層においては想起の対象となる図形に対する表象は観察されず,想起の対象となる図形の情報処理に関する時間的な差のあることが示唆された.そこで,想起の対象となる図形の情報を表象するようになるまでの潜時を定量し比較したところ,第5層においては第6層よりも早く想起の対象となる図形の情報を表象するようになることが確認された.第5層においては想起の対象に対し神経活動を示すニューロンの1/3が150 msよりも短い潜時をもち,これらはフィードフォワード処理8,9) により生成されることが示唆された.一方,提示された手がかりとなる図形の情報の表象に関しては,第6層においてはより短い期間しか保持されていなかった.
これら想起の対象となる図形および手がかりとなる図形に対する時間の経過をもとにクラスター解析により第5層および第6層のニューロンを分類したところ,第6層においては2つのクラスターに分割された.1つ目のクラスターである早期グループは,想起の対象となる図形の情報を早い段階から表象し,また,手がかりとなる図形の情報も相対的に長く保持し,結果として,想起の対象となる図形の情報および手がかりとなる図形の情報を表象するタイミングが重なりあう傾向にあった.2つ目のクラスターである後期グループは,想起の対象となる図形の情報をより遅いタイミングで表象しはじめ,手がかりとなる図形の情報を早い段階で失うため,想起の対象となる図形の情報および手がかりとなる図形の情報を表象するタイミングは重なりあわない傾向にあった.そのため,このグループのニューロンは遅延期間の終盤においては想起の対象となる図形の情報のみを表象していた.これら2つのグループは同じ第6層に存在するにもかかわらず異なる性質を示したことから,第6層における別々の神経基盤にもとづきはたらく可能性が考えられた.そこで,局所の神経活動の総体が反映される局所フィールド電位とおのおののニューロンの発火とのあいだのコヒーレンスを計算してその同期的な神経活動について調べた.その結果,後期グループにおいては遅延期間においてθ帯域における局所フィールド電位と同期した神経活動が上昇した.このことから,最終的に想起の対象となる図形の情報のみを表象するようになる後期グループはθ帯域において協調的に活動し,想起した情報を脳のほかの領域へと出力することが示唆された.
おわりに
この研究により,霊長類では側頭葉の連合野である36野において,大脳皮質の第5層は記憶情報の連合に関与し,そこから想起の対象へと変換された情報を第6層が処理すること,また,想起の対象を処理する第6層の一部は互いに同期して活動しながら情報を脳のほかの領域に出力することが示唆された(図2).これらは,図形の複雑な情報が漸進的に処理されるという初期の視覚野における神経回路モデル2) とは異なる動作であり,側頭葉における記憶の神経回路にはそのための独自の機構が備わることが示された.初期の感覚野との神経回路の違いがどのように生じているか,また,ほかの高次の認知機能を担う連合野においてはどうなのか,今後の研究が待たれる.また,この研究において開発されたMRI法を併用したニューロンの位置の同定はほかの連合野における研究においても適用が可能であり,ほかのさまざまな高次の認知機能の情報処理においても大脳皮質の層構造にそった神経回路がはたす役割が明らかにされることが期待される.
文 献
- Douglas, R. J. & Martin, K. A.: Neuronal circuits of the neocortex. Annu. Rev. Neurosci., 27, 419-451 (2004)[PubMed]
- Reid, R. C. & Alonso, J. M.: The processing and encoding of information in the visual cortex. Curr. Opin. Neurobiol., 6, 475-480 (1996)[PubMed]
- Miyashita, Y.: Cognitive memory: cellular and network machineries and their top-down control. Science, 306, 435-440 (2004)[PubMed]
- Squire, L. R., Stark, C. E. & Clark, R. E.: The medial temporal lobe. Annu. Rev. Neurosci., 27, 279-306 (2004)[PubMed]
- Takeuchi, D., Hirabayashi, T., Tamura, K. et al.: Reversal of interlaminar signal between sensory and memory processing in monkey temporal cortex. Science, 331, 1443-1447 (2011)[PubMed]
- Matsui, T., Koyano, K. W., Koyama, M. et al.: MRI-based localization of electrophysiological recording sites within the cerebral cortex at single-voxel accuracy. Nat. Methods, 4, 161-168 (2007)[PubMed]
- Koyano, K. W., Machino, A., Takeda, M. et al.: In vivo visualization of single-unit recording sites using MRI-detectable elgiloy deposit marking. J. Neurophysiol., 105, 1380-1392 (2011)[PubMed]
- Naya, Y., Yoshida, M. & Miyashita, Y.: Forward processing of long-term associative memory in monkey inferotemporal cortex. J. Neurosci., 23, 2861-2871 (2003)[PubMed]
- Hirabayashi, T., Takeuchi, D., Tamura, K. et al.: Microcircuits for hierarchical elaboration of object coding across primate temporal areas. Science, 341, 191-195 (2013)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2008年 東京大学大学院医学系研究科研究科博士課程 修了,同 研究員,同 助教を経て,2014年より米国NIH National Institute of Mental Health博士研究員.
研究テーマ:視覚の情報処理にかかわる高次連合野の神経回路.
抱負:知覚の心的な表象の機構を,錯視,注意,想起などの観点から明らかにしていきたい.
竹田 真己(Masaki Takeda)
順天堂大学大学院医学研究科 特任准教授.
宮下 保司(Yasushi Miyashita)
順天堂大学大学院医学研究科 特任教授.
研究室URL:http://www.physiol.m.u-tokyo.ac.jp/
© 2016 小谷野賢治・竹田真己・宮下保司 Licensed under CC 表示 2.1 日本
