到達運動の学習を推進する運動野からもたらされる誤差の信号
北澤 茂・井上雅仁
(大阪大学脳情報通信融合研究センター)
email:北澤 茂,井上雅仁
DOI: 10.7875/first.author.2016.057
Error signals in motor cortices drive adaptation in reaching.
Masato Inoue, Motoaki Uchimura, Shigeru Kitazawa
Neuron, 90, 1114-1126 (2016)
目標に手を伸ばす運動は,プリズムにより視野をずらしたり外乱をあたえたりすると誤差を減らすように順応する.大脳皮質の運動野が誤差の信号を提供するという仮説が提唱された一方で,ヒトの脳機能イメージング法においては頭頂葉の連合野だけが誤差を表現すると報告されてきた.運動野は本当に誤差の信号を提供して学習を推進するのだろうか.筆者らは,サルを用いて1次運動野および運動前野が到達運動の終点の誤差の情報を表現することを確かめた.さらに,運動の直後に微小な電気刺激をあたえることにより,誤差は試行を重ねるごとに徐々に蓄積し,その残効は徐々に減少することが明らかにされた.これらの結果から,運動野から発せられる到達運動の誤差の信号は順応を推進することが明確に示された.
運動がうまくなるには練習が欠かせない.はじめは下手だった運動が,こつこつと練習をくり返すことによりうまくなる.うまくなるのは,うまくなった望ましい状態との差,すなわち,運動の誤差を減らすよう脳が学習するからにほかならない.1970年,このような運動の学習の過程において小脳が重要な役割をはたすことが予想され,延髄の下オリーブ核から小脳に投射する登上線維が学習における教師の役割をはたすと想定された1).実際に,眼球の運動の制御に関しては,網膜像のブレを抑えるための反射から,すばやいサッケード運動,動く物体を目で追いかける滑動性の追跡運動にいたるまで,この枠組みの成立することが確かめられた2).手の随意運動においても基本的には同様である.1977年には,外乱に抵抗してハンドルを保持する課題において,外乱に対し登上線維が応答することが報告された3).物をつかむとき(最近では,スマートフォンの画面をタッチするとき)に日々行う到達運動については,筆者らにより,手と対象のあいだの誤差の情報が登上線維により伝達されることが明らかにされた4).しかし,少し視野を広げて,登上線維に届く誤差の信号の由来を考えてみると,これがじつのところ,まったくわかっていなかった(図1).
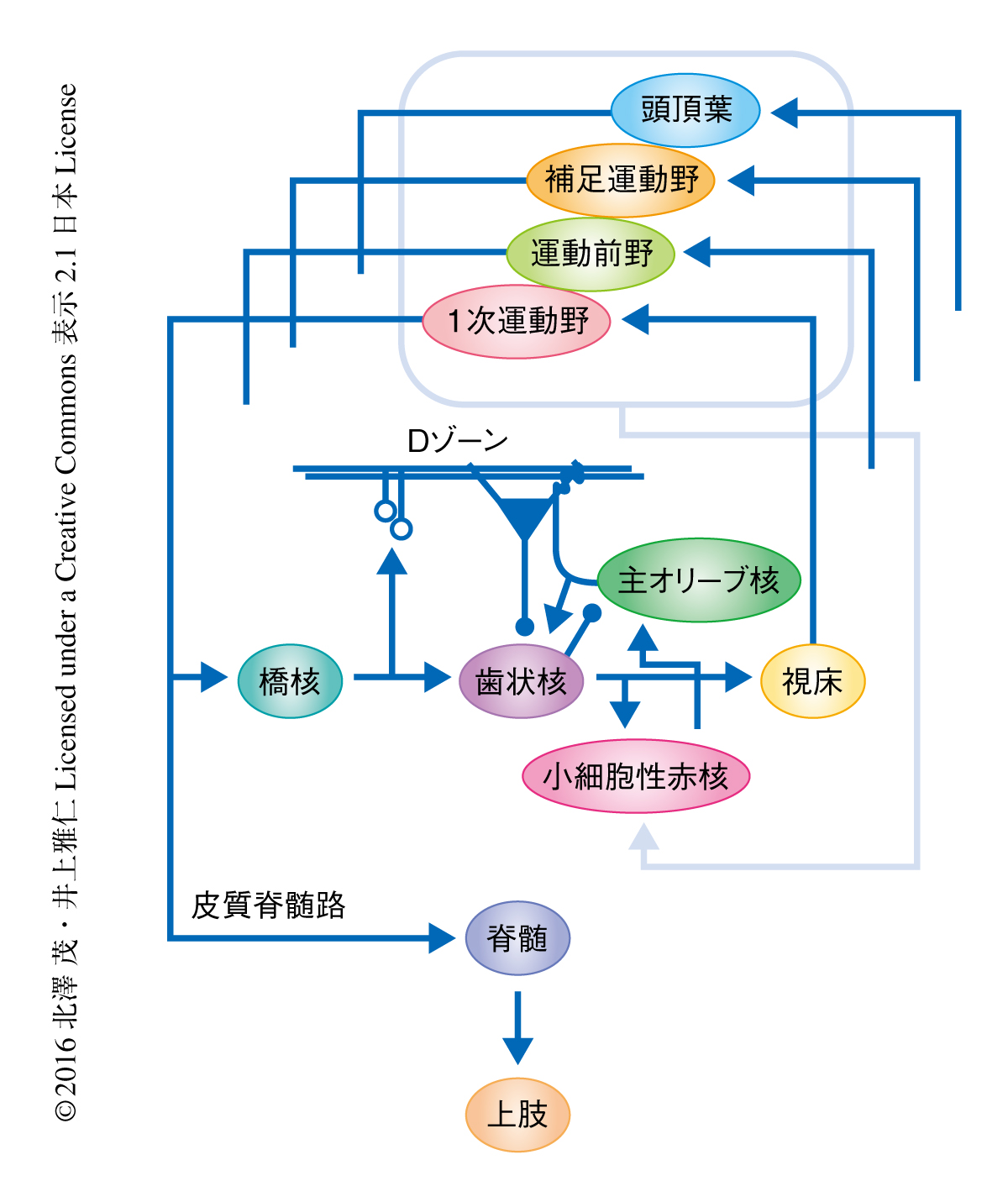
到達運動の制御に大脳皮質の1次運動野が必要であることには異論はないだろう.到達運動のたくみな制御に必要な時系列信号は,1次運動野-橋核-歯状核-視床-1次運動野という興奮性のループと,このループに適切なタイミングでブレーキをかける抑制性のプルキンエ細胞により生成される(図1).これらのループにおいて生成された信号は,そのまま皮質脊髄路により脊髄前核の運動ニューロンに伝達され,20個をこえる筋肉をたくみに収縮あるいは弛緩させて腕を動かす.この小脳の外側の半球部を含むループに登上線維を伸ばす主オリーブ核は,小細胞性赤核から入力をうける2).その小細胞性赤核は前頭葉の運動野や頭頂葉などから幅広い投射をうける5).ということは,腕の運動を改善するための誤差の信号はこれらの領域のどこからきてもよいことになる.ところが,ヒトを対象とした非侵襲脳機能画像法を用いた研究においては,到達運動の誤差と相関して血流が増加したのは頭頂葉だけであった6,7).これが正しければ,到達運動の誤差と運動野とは無関係ということになる.一方,サルを使った研究においては,運動前野の活動をGABAA受容体の作動薬であるムシモールにより抑制すると到達運動の誤差が修正されなくなると報告された8).また,誤差を挽回するために生じる運動野のフィードバック運動指令が分岐して,そのまま登上線維への信号になるというフィードバック誤差学習説も提唱された9).これらの研究にしたがえば,運動前野や1次運動野にも誤差を表現して学習を推進する十二分な資格があると思われる.
そこで,この研究においては,到達運動の学習を推進する誤差の信号が大脳皮質の運動前野あるいは1次運動野からもたらされるのかどうかを明らかにすることを目的とした.具体的には,運動野あるいは1次運動野が到達運動の終点の誤差の情報を伝達するのかどうか,伝達しているとすれば,その活動が本当に学習をひき起こすのかどうかを調べた.
目のまえの画面に現われる十字の目標にむかって手を伸ばし,触れたら手の位置を動かさずに1秒間保持するようにサルを訓練した.運動中は液晶シャッターを使って視覚の情報を遮断し,画面に触れた瞬間にもういちど液晶シャッターを開いて0.3秒だけ運動の結果を見せた.しかし,ただ運動させるとほとんど誤差を生じることなく到達運動をしてしまう.これは誤差の研究には具合が悪い.そこで,コンピューターにより制御したプリズムを使って視野をランダムな方向にずらした.こうすることにより,目標が見える場所に手を伸ばしても手が右や左に少しずれて到着するので,誤差と神経の活動との関係を調べることが可能になる.すると,たとえば,あるひとつの運動前野のニューロンは,運動の直後に右下の誤差に対しよく応答することがわかった.このようなニューロンの応答が誤差に関してもつ情報量を計算したところ,運動前野においては調べたニューロンのおよそ3割が有意な情報をもっていた.情報量のピーク潜時は誤差の入力から90ミリ秒だった.1次運動野も同様で,調べたニューロンの約3割が誤差の情報をコードしていた.ただ,ピーク潜時はやや遅れて230ミリ秒だった.いずれにしても,到達運動の誤差は運動前野も1次運動野も表現していた.また,ニューロンごとによく応答する誤差の方向はまちまちで,全体としてあらゆる方向に分布していた.
誤差の信号が到達運動の学習にかかわることを証明するため,運動の直後に微小な電気刺激をあたえた.たとえば,“右上の誤差”を選好するニューロンが学習にかかわっているなら,このニューロンが活動したつぎの試行においては,到着点が少しだけ“左下”に修正されるはずである.結果は予想どおりで,右上の誤差に応答する領域を運動の直後に刺激すると,誤差は徐々に累積して左下へとずれていった.刺激をやめると,誤差は試行ごとに少しずつ減少し,約30回かけてもとにもどった.運動前野および1次運動野の計31か所に刺激をあたえたところ,15か所で有意な効果が認められた.つまり,1次運動野あるいは運動前野のどちらも,選好する誤差の方向の反対への運動の学習を推進する力があった.
刺激のタイミングを0.1秒遅らせるだけで電気刺激の効果は消失した.つまり,誤差の信号は0.1秒遅れるだけで“後の祭り”になってしまうのだ.運動の直後の0.1秒のあいだに運動野を通過する誤差の信号だけが順応を推進する力をもっていた.
さきの実験では運動閾値より小さな電気刺激をあたえたので,実際には運動は生じなかった.しかし,運動閾値をこえる電気刺激をあたえると,活動する筋肉が選好する誤差の方向は,ニューロンの選好する誤差の方向とほぼ一致した.わかりやすくいうと,運動野あるいは1次運動野の活動には,誤差を減少する方向に現在の手の位置を動かすという作用もあることがわかった.これはまさに,誤差を減少させるためのフィードバック信号が学習用の信号として小脳に伝達される,というフィードバック誤差学習説9) を支持する結果であった.
今回の研究により,運動前野および1次運動野に到達運動の誤差が表現され,その誤差を減少するような運動の学習の原因になることが明らかにされた.その作用の部位としては小脳を想定していたが,今回の結果には,作用の部位が小脳であるかどうかを決める情報はない.この点を明らかにするには,たとえば,大脳皮質から小細胞性赤核への投射を適切な手法により可逆的にブロックし,電気刺激による人工的な順応の効果が消失するかどうかを調べるような実験が必要だろう.小細胞性赤核は大脳皮質の誤差の信号を小脳に伝達する重要な中継核であるにもかかわらず,ほとんど研究されていない.今後の研究の展開が待たれるところである.
さらに付言すれば,筆者の最大の関心事は誤差を減少させる神経機構の解明ではない.到達運動は開始から終了までを制御する信号のパターンが全体として,躍度の時間積分を最小化する10),トルクの変化を最小化する11),終点の誤差の分散を最小化する12),ように最適化されているという.この時系列パターンの最適化の機構こそ,最終的に明らかにしたい標的である13).じつは,この研究において開発した電気刺激により人工的に誤差の信号を注入する実験系を使えば,この目標に近づけるのではないかと考えている.
略歴:1991年 東京大学大学院医学系研究科 修了,1995年 工業技術院電子技術総合研究所 主任研究官,2003年 順天堂大学医学部 教授を経て,2011年より大阪大学大学院生命機能研究科 教授.
研究テーマ:到達運動の最適化の神経機構,時間および空間の認識機構.
関心事:新学術領域研究“こころの時間学”を発展させること.目を動かしても世界が動かない理由の追求が目下のマイブームで,Whatの認識に成功した人工知能をWhereあるいはWhenの認識にも拡張してみたい.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/181/
井上 雅仁(Masato Inoue)
脳情報通信融合研究センター 研究員.
© 2016 北澤 茂・井上雅仁 Licensed under CC 表示 2.1 日本
(大阪大学脳情報通信融合研究センター)
email:北澤 茂,井上雅仁
DOI: 10.7875/first.author.2016.057
Error signals in motor cortices drive adaptation in reaching.
Masato Inoue, Motoaki Uchimura, Shigeru Kitazawa
Neuron, 90, 1114-1126 (2016)
要 約
目標に手を伸ばす運動は,プリズムにより視野をずらしたり外乱をあたえたりすると誤差を減らすように順応する.大脳皮質の運動野が誤差の信号を提供するという仮説が提唱された一方で,ヒトの脳機能イメージング法においては頭頂葉の連合野だけが誤差を表現すると報告されてきた.運動野は本当に誤差の信号を提供して学習を推進するのだろうか.筆者らは,サルを用いて1次運動野および運動前野が到達運動の終点の誤差の情報を表現することを確かめた.さらに,運動の直後に微小な電気刺激をあたえることにより,誤差は試行を重ねるごとに徐々に蓄積し,その残効は徐々に減少することが明らかにされた.これらの結果から,運動野から発せられる到達運動の誤差の信号は順応を推進することが明確に示された.
はじめに
運動がうまくなるには練習が欠かせない.はじめは下手だった運動が,こつこつと練習をくり返すことによりうまくなる.うまくなるのは,うまくなった望ましい状態との差,すなわち,運動の誤差を減らすよう脳が学習するからにほかならない.1970年,このような運動の学習の過程において小脳が重要な役割をはたすことが予想され,延髄の下オリーブ核から小脳に投射する登上線維が学習における教師の役割をはたすと想定された1).実際に,眼球の運動の制御に関しては,網膜像のブレを抑えるための反射から,すばやいサッケード運動,動く物体を目で追いかける滑動性の追跡運動にいたるまで,この枠組みの成立することが確かめられた2).手の随意運動においても基本的には同様である.1977年には,外乱に抵抗してハンドルを保持する課題において,外乱に対し登上線維が応答することが報告された3).物をつかむとき(最近では,スマートフォンの画面をタッチするとき)に日々行う到達運動については,筆者らにより,手と対象のあいだの誤差の情報が登上線維により伝達されることが明らかにされた4).しかし,少し視野を広げて,登上線維に届く誤差の信号の由来を考えてみると,これがじつのところ,まったくわかっていなかった(図1).
到達運動の制御に大脳皮質の1次運動野が必要であることには異論はないだろう.到達運動のたくみな制御に必要な時系列信号は,1次運動野-橋核-歯状核-視床-1次運動野という興奮性のループと,このループに適切なタイミングでブレーキをかける抑制性のプルキンエ細胞により生成される(図1).これらのループにおいて生成された信号は,そのまま皮質脊髄路により脊髄前核の運動ニューロンに伝達され,20個をこえる筋肉をたくみに収縮あるいは弛緩させて腕を動かす.この小脳の外側の半球部を含むループに登上線維を伸ばす主オリーブ核は,小細胞性赤核から入力をうける2).その小細胞性赤核は前頭葉の運動野や頭頂葉などから幅広い投射をうける5).ということは,腕の運動を改善するための誤差の信号はこれらの領域のどこからきてもよいことになる.ところが,ヒトを対象とした非侵襲脳機能画像法を用いた研究においては,到達運動の誤差と相関して血流が増加したのは頭頂葉だけであった6,7).これが正しければ,到達運動の誤差と運動野とは無関係ということになる.一方,サルを使った研究においては,運動前野の活動をGABAA受容体の作動薬であるムシモールにより抑制すると到達運動の誤差が修正されなくなると報告された8).また,誤差を挽回するために生じる運動野のフィードバック運動指令が分岐して,そのまま登上線維への信号になるというフィードバック誤差学習説も提唱された9).これらの研究にしたがえば,運動前野や1次運動野にも誤差を表現して学習を推進する十二分な資格があると思われる.
そこで,この研究においては,到達運動の学習を推進する誤差の信号が大脳皮質の運動前野あるいは1次運動野からもたらされるのかどうかを明らかにすることを目的とした.具体的には,運動野あるいは1次運動野が到達運動の終点の誤差の情報を伝達するのかどうか,伝達しているとすれば,その活動が本当に学習をひき起こすのかどうかを調べた.
1.到達運動の誤差は運動前野も1次運動野も表現していた
目のまえの画面に現われる十字の目標にむかって手を伸ばし,触れたら手の位置を動かさずに1秒間保持するようにサルを訓練した.運動中は液晶シャッターを使って視覚の情報を遮断し,画面に触れた瞬間にもういちど液晶シャッターを開いて0.3秒だけ運動の結果を見せた.しかし,ただ運動させるとほとんど誤差を生じることなく到達運動をしてしまう.これは誤差の研究には具合が悪い.そこで,コンピューターにより制御したプリズムを使って視野をランダムな方向にずらした.こうすることにより,目標が見える場所に手を伸ばしても手が右や左に少しずれて到着するので,誤差と神経の活動との関係を調べることが可能になる.すると,たとえば,あるひとつの運動前野のニューロンは,運動の直後に右下の誤差に対しよく応答することがわかった.このようなニューロンの応答が誤差に関してもつ情報量を計算したところ,運動前野においては調べたニューロンのおよそ3割が有意な情報をもっていた.情報量のピーク潜時は誤差の入力から90ミリ秒だった.1次運動野も同様で,調べたニューロンの約3割が誤差の情報をコードしていた.ただ,ピーク潜時はやや遅れて230ミリ秒だった.いずれにしても,到達運動の誤差は運動前野も1次運動野も表現していた.また,ニューロンごとによく応答する誤差の方向はまちまちで,全体としてあらゆる方向に分布していた.
2.運動の直後の電気刺激により人工的な順応が生じた
誤差の信号が到達運動の学習にかかわることを証明するため,運動の直後に微小な電気刺激をあたえた.たとえば,“右上の誤差”を選好するニューロンが学習にかかわっているなら,このニューロンが活動したつぎの試行においては,到着点が少しだけ“左下”に修正されるはずである.結果は予想どおりで,右上の誤差に応答する領域を運動の直後に刺激すると,誤差は徐々に累積して左下へとずれていった.刺激をやめると,誤差は試行ごとに少しずつ減少し,約30回かけてもとにもどった.運動前野および1次運動野の計31か所に刺激をあたえたところ,15か所で有意な効果が認められた.つまり,1次運動野あるいは運動前野のどちらも,選好する誤差の方向の反対への運動の学習を推進する力があった.
3.電気刺激の効果は運動の直後の0.1秒に限られた
刺激のタイミングを0.1秒遅らせるだけで電気刺激の効果は消失した.つまり,誤差の信号は0.1秒遅れるだけで“後の祭り”になってしまうのだ.運動の直後の0.1秒のあいだに運動野を通過する誤差の信号だけが順応を推進する力をもっていた.
4.電気刺激を強くした際の筋肉の活動は選好する方向の誤差を減少させた
さきの実験では運動閾値より小さな電気刺激をあたえたので,実際には運動は生じなかった.しかし,運動閾値をこえる電気刺激をあたえると,活動する筋肉が選好する誤差の方向は,ニューロンの選好する誤差の方向とほぼ一致した.わかりやすくいうと,運動野あるいは1次運動野の活動には,誤差を減少する方向に現在の手の位置を動かすという作用もあることがわかった.これはまさに,誤差を減少させるためのフィードバック信号が学習用の信号として小脳に伝達される,というフィードバック誤差学習説9) を支持する結果であった.
おわりに
今回の研究により,運動前野および1次運動野に到達運動の誤差が表現され,その誤差を減少するような運動の学習の原因になることが明らかにされた.その作用の部位としては小脳を想定していたが,今回の結果には,作用の部位が小脳であるかどうかを決める情報はない.この点を明らかにするには,たとえば,大脳皮質から小細胞性赤核への投射を適切な手法により可逆的にブロックし,電気刺激による人工的な順応の効果が消失するかどうかを調べるような実験が必要だろう.小細胞性赤核は大脳皮質の誤差の信号を小脳に伝達する重要な中継核であるにもかかわらず,ほとんど研究されていない.今後の研究の展開が待たれるところである.
さらに付言すれば,筆者の最大の関心事は誤差を減少させる神経機構の解明ではない.到達運動は開始から終了までを制御する信号のパターンが全体として,躍度の時間積分を最小化する10),トルクの変化を最小化する11),終点の誤差の分散を最小化する12),ように最適化されているという.この時系列パターンの最適化の機構こそ,最終的に明らかにしたい標的である13).じつは,この研究において開発した電気刺激により人工的に誤差の信号を注入する実験系を使えば,この目標に近づけるのではないかと考えている.
文 献
- Ito, M.: Neurophysiological aspects of the cerebellar motor control system. Int. J. Neurol., 7, 162-176 (1970)[PubMed]
- Ito, M.: Error detection and representation in the olivo-cerebellar system. Front. Neural Circuits, 7, 1 (2013)[PubMed]
- Gilbert, P. F. & Thach, W. T.: Purkinje cell activity during motor learning. Brain Res., 128, 309-328 (1977)[PubMed]
- Kitazawa, S., Kimura, T. & Yin, P. -B.: Cerebellar complex spikes encode both destinations and errors in arm movements. Nature, 392, 494-497 (1998)[PubMed]
- Burman, K., Darian-Smith, C. & Darian-Smith, I.: Macaque red nucleus: origin of spinal and olivary projections and terminations of cortical inputs. J. Comp. Neurol., 423, 179-196 (2000)[PubMed]
- Diedrichsen, J., Hashambhoy, Y., Rane, T. et al.: Neural correlates of reach errors. J. Neurosci., 25, 9919-9931 (2005)[PubMed]
- Luaute, J., Schwartz, S., Rossetti, Y. et al.: Dynamic changes in brain activity during prism adaptation. J. Neurosci., 29, 169-178 (2009)[PubMed]
- Kurata, K. & Hoshi, E.: Reacquisition deficits in prism adaptation after muscimol microinjection into the ventral premotor cortex of monkeys. J. Neurophysiol., 81, 1927-1938 (1999)[PubMed]
- Kawato, M. & Gomi, H.: A computational model of four regions of the cerebellum based on feedback-error learning. Biol. Cybern., 68, 95-103 (1992)[PubMed]
- Flash, T. & Hogan, N.: The coordination of arm movements: an experimentally confirmed mathematical model. J. Neurosci., 5, 1688-1703 (1985)[PubMed]
- Uno, Y., Kawato, M. & Suzuki, R.: Formation and control of optimal trajectory in human multijoint arm movement. Minimum torque-change model. Biol. Cybern., 61, 89-101 (1989)[PubMed]
- Harris, C. M. & Wolpert, D. M.: Signal-dependent noise determines motor planning. Nature, 394, 780-784 (1998)[PubMed]
- Kitazawa, S.: Optimization of goal-directed movements in the cerebellum: a random walk hypothesis. Neurosci. Res., 43, 289-294 (2002)[PubMed]
著者プロフィール
略歴:1991年 東京大学大学院医学系研究科 修了,1995年 工業技術院電子技術総合研究所 主任研究官,2003年 順天堂大学医学部 教授を経て,2011年より大阪大学大学院生命機能研究科 教授.
研究テーマ:到達運動の最適化の神経機構,時間および空間の認識機構.
関心事:新学術領域研究“こころの時間学”を発展させること.目を動かしても世界が動かない理由の追求が目下のマイブームで,Whatの認識に成功した人工知能をWhereあるいはWhenの認識にも拡張してみたい.
研究室URL:http://www.fbs.osaka-u.ac.jp/jpn/general/lab/181/
井上 雅仁(Masato Inoue)
脳情報通信融合研究センター 研究員.
© 2016 北澤 茂・井上雅仁 Licensed under CC 表示 2.1 日本
