中心子の構造の決定機構におけるカートホイールと微小管との動的な相互作用
苗加 彰・廣野雅文
(法政大学生命科学部 生命機能学科)
email:廣野雅文
DOI: 10.7875/first.author.2016.030
SAS-6 engineering reveals interdependence between cartwheel and microtubules in determining centriole architecture.
Manuel Hilbert, Akira Noga, Daniel Frey, Virginie Hamel, Paul Guichard, Sebastian H. W. Kraatz, Moritz Pfreundschuh, Sarah Hosner, Isabelle Flückiger, Rolf Jaussi, Mara M. Wieser, Katherine M. Thieltges, Xavier Deupi, Daniel J. Müller, Richard A. Kammerer, Pierre Gönczy, Masafumi Hirono, Michel O. Steinmetz
Nature Cell Biology, 18, 393-403 (2016)
中心子は真核生物に存在する中心体の中核となり,繊毛および鞭毛の形成の基部としても機能するオルガネラである.9本の三連微小管が回転対称に配置した特徴的な骨格の構造をもつが,この構造の形成にはカートホイールという9回対称性の車輪状の構造に依存する機構,および,カートホイールには依存しない機構の2つがはたらく.しかし,両者の関係,および,カートホイールに非依存性の機構の実体は未解明であった.この研究においては,カートホイールの主要な構成タンパク質であるSAS-6に変異を導入することにより,形成されるカートホイールの回転対称性を変化させ,その際の中心子の構造の変化について検討した.6回対称性のカートホイールを形成するSAS-6を作製しクラミドモナスのSAS-6欠失株に発現させたところ,細胞において形成された中心子の多くは9回対称性であり,そこに含まれるカートホイールも多くが9回対称性であった.しかし,カートホイールと周囲の三連微小管とのあいだに介在するタンパク質の機能を低下させたところ,6回対称性のカートホイールが形成された.これらの結果から,三連微小管それ自体が9回対称性に集合する固有の性質をもち,それがカートホイールの対称性にも影響することが示唆された.したがって,カートホイールと中心子微小管はそれぞれ独立に集合し,それらのあいだのダイナミックな相互作用により中心子の9回対称性が確立される可能性が考えられた.
中心子(centriole)は真核生物において微小管からなる細胞内の構造の形成にまさに中心的な役割を担うオルガネラであり,中心体の中核として細胞質微小管の重合の中心となり,鞭毛および繊毛の形成の基部としてその内部の構造の9 + 2パターンを規定する1,2).中心子の形成の異常や構成タンパク質の変異は,がん化や,小脳症,水頭症,内臓逆位など繊毛病とよばれる一連の疾患の原因となる3).中心子は9本の三連微小管が回転対称に配置した特徴的な円筒状の構造をもつ(図1a).この9回対称性の構造は高等植物や酵母を除くほとんどすべての真核生物に共通する.なぜ“9”なのか,どのようにして厳密に“9”という数に決定されるのかについては長いあいだ謎であったが,最近になり,緑藻クラミドモナスの変異株を用いた解析から,カートホイールという構造が重要な役割をはたすことが明らかにされてきた4-6).

カートホイールは中心子の内腔の底部にある車輪状の構造で,中心のリング(ハブ)とそこから放射状に伸びる9本の繊維(スポーク)からなる1)(図1b).クラミドモナスにおいてこの構造を欠失するbld12変異株が単離され,その表現型および変異遺伝子産物であるSAS-6の解析により,以下のことが明らかにされた.1)bld12変異株においては中心子を構成する三連微小管の数が9本に固定されず7~11本の中心子が形成される5).2)しかし,bld12変異株においてももっとも多く形成されるのは三連微小管が9本の中心子である.したがって,中心子の9回対称性の確立にはカートホイールに依存しない機構もはたらくと考えられる5).3)SAS-6は2つの球状の頭部と1本の棒状の尾部からなる二量体を形成し,9個の二量体が回転対称に会合することによりカートホイールの中心の部分と相同の構造を形成する7,8)(図1b).これらの知見は,中心子の9回対称性の謎を解くものとして注目され,SAS-6二量体の自己集合によりカートホイールが形成されたのち,それが足場となり中心子の9回対称性が確立される,という構築モデルが広く受け入れられてきた2).しかし,カートホイールに依存しない機構も同様に重要であるという事実には注意がはらわれなかった.
この研究においては,カートホイールに依存しない機構についての手がかりを得るため,SAS-6に変異を導入し,その会合性の変化が中心子の構造に及ぼす影響について検討した.
組換えSAS-6はin vitroにおいて会合しカートホイールと相同の放射状の構造を形成する.X線結晶構造の情報をもとに,SAS-6二量体のあいだの結合面に結合能を強化するような複数の変異を導入し,それらを組み合わせて28種類の変異SAS-6を作製した.それらのin vitroにおける会合体を原子間力顕微鏡により観察したところ,多くが二量体の数が9個よりも少ない会合体を形成することがわかった.なかでも,NN24と名づけた変異SAS-6はとくに少数で会合し,おもに6回対称性のカートホイール様の構造を形成した.
SAS-6の会合性の変化が中心子の構造に及ぼす影響について調べるため,NN24をクラミドモナスのbld12変異株に発現させ,中心子の構造を透過型電子顕微鏡により観察した.野生型のSAS-6を発現させた対照実験ではすべての中心子が9回対称性であったが(図2a),NN24を発現させた場合は8回対称性の中心子が約33%をしめ,残りは9回対称性であった(図2b).
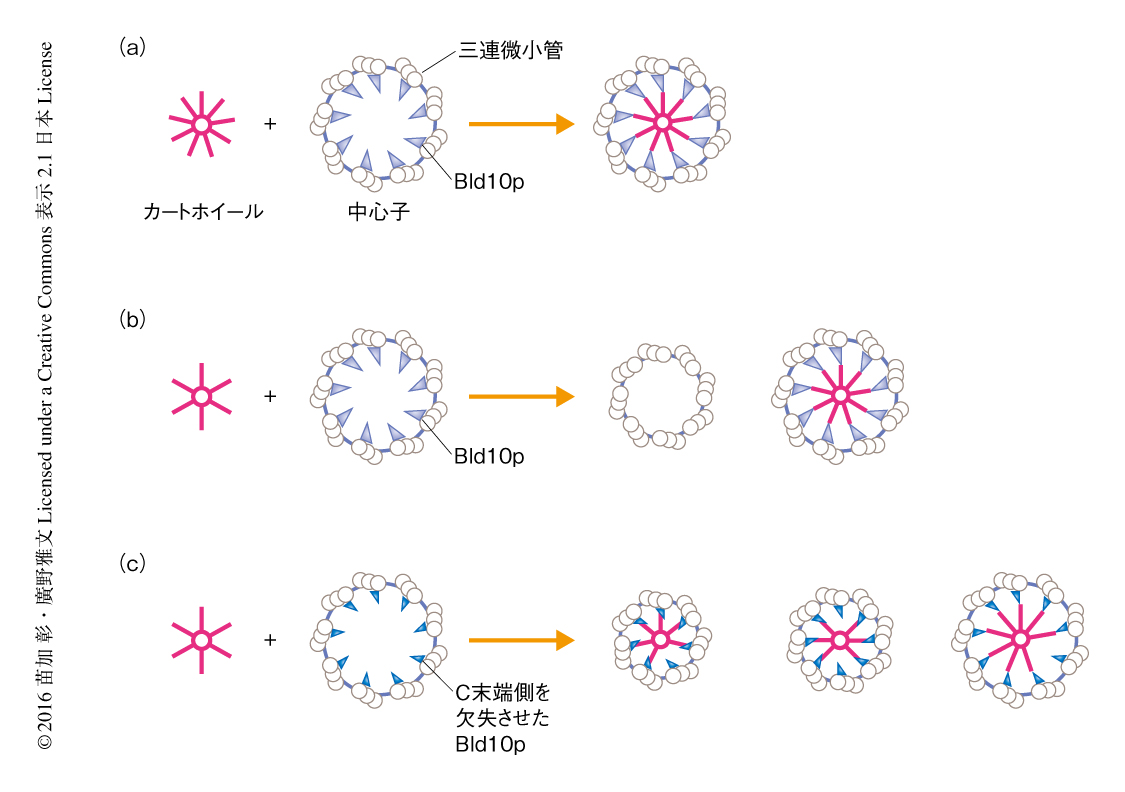
NN24を発現させた際に形成された9回対称性の中心子が内包するカートホイールを観察したところ,6回対称性のカートホイールはまったく観察されず,9回対称性のカートホイールが高頻度で観察された(図2b).したがって,中心子の9回対称性の決定にはたらく未知の要因は,三連微小管の集合状態だけではなく,カートホイールの構造にも影響を及ぼすと考えられた.
カートホイールを欠失するbld12変異株においても,多くの場合,中心子の三連微小管は9回対称性に集合するという事実から,この未知の要因はカートホイールの周囲にリング状に配置した三連微小管にあると推察した.この推察が正しければ,微小管とカートホイールとのあいだの相互作用を弱めるとカートホイールの構造になんらかの変化が起こると期待された.
以前に,筆者らは,クラミドモナスのbld10変異株の解析から,カートホイールと微小管とが結合する部分に局在するタンパク質Bld10pを同定した4,6).そして,Bld10pのN末端側の54%あるいはC末端側の35%の配列を欠失させた断片をbld10変異株に発現させると,カートホイールと微小管との結合が不安定になった6).そこで,このBld10pの断片をNN24と同時にbld10bld12二重変異株に発現させてカートホイールの構造を観察したところ,6~8回対称性のカートホイール,つまり,変異SAS-6のin vitroにおける会合性がより忠実に反映されたカートホイールが観察された(図2c).したがって,未知の機構はカートホイールの周囲に存在する三連微小管のリング状の集合体にあり,中心子の9回対称性の確立には微小管とカートホイールとの相互作用が重要であると結論された.
カートホイールのかたちがSAS-6の会合性とは異なる要因の影響をうけるという今回の発見は,SAS-6の会合により形成されたカートホイールが鋳型となるという,現在,優勢になりつつある中心子の構築のモデルに異をとなえるものであった.この研究の結果から,微小管とカートホイールは独立に形成され,それらのあいだの相互作用の結果,9回対称性の中心子だけが安定化されて残る,という機構が考えられた.いい換えると,中心子の普遍的なかたちは独立したタンパク質の集合の過程が順に起こって決定されるのではなく,カートホイールと微小管との相互作用というダイナミックな過程をへて決定されると考えられた.
今回の研究により,カートホイールに依存しない未知の機構は,環状に配置した微小管にあることが明らかにされた.この分子機構を明らかにするのが今後の課題である.筆者らは,微小管のあいだの架橋の構造は,それらを一定の角度に保ち正九角形の配置をとりやすくする機能があると推察している.この架橋を担う実体を明らかにすることは,普遍的な中心子の9回対称性の決定機構を解明するうえで重要な課題である.
略歴:東京大学大学院理学系研究科博士課程 在学中.
廣野 雅文(Masafumi Hirono)
法政大学生命科学部 教授.
© 2016 苗加 彰・廣野雅文 Licensed under CC 表示 2.1 日本
(法政大学生命科学部 生命機能学科)
email:廣野雅文
DOI: 10.7875/first.author.2016.030
SAS-6 engineering reveals interdependence between cartwheel and microtubules in determining centriole architecture.
Manuel Hilbert, Akira Noga, Daniel Frey, Virginie Hamel, Paul Guichard, Sebastian H. W. Kraatz, Moritz Pfreundschuh, Sarah Hosner, Isabelle Flückiger, Rolf Jaussi, Mara M. Wieser, Katherine M. Thieltges, Xavier Deupi, Daniel J. Müller, Richard A. Kammerer, Pierre Gönczy, Masafumi Hirono, Michel O. Steinmetz
Nature Cell Biology, 18, 393-403 (2016)
要 約
中心子は真核生物に存在する中心体の中核となり,繊毛および鞭毛の形成の基部としても機能するオルガネラである.9本の三連微小管が回転対称に配置した特徴的な骨格の構造をもつが,この構造の形成にはカートホイールという9回対称性の車輪状の構造に依存する機構,および,カートホイールには依存しない機構の2つがはたらく.しかし,両者の関係,および,カートホイールに非依存性の機構の実体は未解明であった.この研究においては,カートホイールの主要な構成タンパク質であるSAS-6に変異を導入することにより,形成されるカートホイールの回転対称性を変化させ,その際の中心子の構造の変化について検討した.6回対称性のカートホイールを形成するSAS-6を作製しクラミドモナスのSAS-6欠失株に発現させたところ,細胞において形成された中心子の多くは9回対称性であり,そこに含まれるカートホイールも多くが9回対称性であった.しかし,カートホイールと周囲の三連微小管とのあいだに介在するタンパク質の機能を低下させたところ,6回対称性のカートホイールが形成された.これらの結果から,三連微小管それ自体が9回対称性に集合する固有の性質をもち,それがカートホイールの対称性にも影響することが示唆された.したがって,カートホイールと中心子微小管はそれぞれ独立に集合し,それらのあいだのダイナミックな相互作用により中心子の9回対称性が確立される可能性が考えられた.
はじめに
中心子(centriole)は真核生物において微小管からなる細胞内の構造の形成にまさに中心的な役割を担うオルガネラであり,中心体の中核として細胞質微小管の重合の中心となり,鞭毛および繊毛の形成の基部としてその内部の構造の9 + 2パターンを規定する1,2).中心子の形成の異常や構成タンパク質の変異は,がん化や,小脳症,水頭症,内臓逆位など繊毛病とよばれる一連の疾患の原因となる3).中心子は9本の三連微小管が回転対称に配置した特徴的な円筒状の構造をもつ(図1a).この9回対称性の構造は高等植物や酵母を除くほとんどすべての真核生物に共通する.なぜ“9”なのか,どのようにして厳密に“9”という数に決定されるのかについては長いあいだ謎であったが,最近になり,緑藻クラミドモナスの変異株を用いた解析から,カートホイールという構造が重要な役割をはたすことが明らかにされてきた4-6).
カートホイールは中心子の内腔の底部にある車輪状の構造で,中心のリング(ハブ)とそこから放射状に伸びる9本の繊維(スポーク)からなる1)(図1b).クラミドモナスにおいてこの構造を欠失するbld12変異株が単離され,その表現型および変異遺伝子産物であるSAS-6の解析により,以下のことが明らかにされた.1)bld12変異株においては中心子を構成する三連微小管の数が9本に固定されず7~11本の中心子が形成される5).2)しかし,bld12変異株においてももっとも多く形成されるのは三連微小管が9本の中心子である.したがって,中心子の9回対称性の確立にはカートホイールに依存しない機構もはたらくと考えられる5).3)SAS-6は2つの球状の頭部と1本の棒状の尾部からなる二量体を形成し,9個の二量体が回転対称に会合することによりカートホイールの中心の部分と相同の構造を形成する7,8)(図1b).これらの知見は,中心子の9回対称性の謎を解くものとして注目され,SAS-6二量体の自己集合によりカートホイールが形成されたのち,それが足場となり中心子の9回対称性が確立される,という構築モデルが広く受け入れられてきた2).しかし,カートホイールに依存しない機構も同様に重要であるという事実には注意がはらわれなかった.
この研究においては,カートホイールに依存しない機構についての手がかりを得るため,SAS-6に変異を導入し,その会合性の変化が中心子の構造に及ぼす影響について検討した.
1.SAS-6への変異の導入による会合性の改変
組換えSAS-6はin vitroにおいて会合しカートホイールと相同の放射状の構造を形成する.X線結晶構造の情報をもとに,SAS-6二量体のあいだの結合面に結合能を強化するような複数の変異を導入し,それらを組み合わせて28種類の変異SAS-6を作製した.それらのin vitroにおける会合体を原子間力顕微鏡により観察したところ,多くが二量体の数が9個よりも少ない会合体を形成することがわかった.なかでも,NN24と名づけた変異SAS-6はとくに少数で会合し,おもに6回対称性のカートホイール様の構造を形成した.
2.変異SAS-6の中心子の構造への影響
SAS-6の会合性の変化が中心子の構造に及ぼす影響について調べるため,NN24をクラミドモナスのbld12変異株に発現させ,中心子の構造を透過型電子顕微鏡により観察した.野生型のSAS-6を発現させた対照実験ではすべての中心子が9回対称性であったが(図2a),NN24を発現させた場合は8回対称性の中心子が約33%をしめ,残りは9回対称性であった(図2b).
3.中心子の構築における三連微小管の集合体の重要性
NN24を発現させた際に形成された9回対称性の中心子が内包するカートホイールを観察したところ,6回対称性のカートホイールはまったく観察されず,9回対称性のカートホイールが高頻度で観察された(図2b).したがって,中心子の9回対称性の決定にはたらく未知の要因は,三連微小管の集合状態だけではなく,カートホイールの構造にも影響を及ぼすと考えられた.
カートホイールを欠失するbld12変異株においても,多くの場合,中心子の三連微小管は9回対称性に集合するという事実から,この未知の要因はカートホイールの周囲にリング状に配置した三連微小管にあると推察した.この推察が正しければ,微小管とカートホイールとのあいだの相互作用を弱めるとカートホイールの構造になんらかの変化が起こると期待された.
以前に,筆者らは,クラミドモナスのbld10変異株の解析から,カートホイールと微小管とが結合する部分に局在するタンパク質Bld10pを同定した4,6).そして,Bld10pのN末端側の54%あるいはC末端側の35%の配列を欠失させた断片をbld10変異株に発現させると,カートホイールと微小管との結合が不安定になった6).そこで,このBld10pの断片をNN24と同時にbld10bld12二重変異株に発現させてカートホイールの構造を観察したところ,6~8回対称性のカートホイール,つまり,変異SAS-6のin vitroにおける会合性がより忠実に反映されたカートホイールが観察された(図2c).したがって,未知の機構はカートホイールの周囲に存在する三連微小管のリング状の集合体にあり,中心子の9回対称性の確立には微小管とカートホイールとの相互作用が重要であると結論された.
4.中心子の構築の機構の新しいモデル
カートホイールのかたちがSAS-6の会合性とは異なる要因の影響をうけるという今回の発見は,SAS-6の会合により形成されたカートホイールが鋳型となるという,現在,優勢になりつつある中心子の構築のモデルに異をとなえるものであった.この研究の結果から,微小管とカートホイールは独立に形成され,それらのあいだの相互作用の結果,9回対称性の中心子だけが安定化されて残る,という機構が考えられた.いい換えると,中心子の普遍的なかたちは独立したタンパク質の集合の過程が順に起こって決定されるのではなく,カートホイールと微小管との相互作用というダイナミックな過程をへて決定されると考えられた.
おわりに
今回の研究により,カートホイールに依存しない未知の機構は,環状に配置した微小管にあることが明らかにされた.この分子機構を明らかにするのが今後の課題である.筆者らは,微小管のあいだの架橋の構造は,それらを一定の角度に保ち正九角形の配置をとりやすくする機能があると推察している.この架橋を担う実体を明らかにすることは,普遍的な中心子の9回対称性の決定機構を解明するうえで重要な課題である.
文 献
- Hirono, M.: Cartwheel assembly. Philos. Trans. R. Soc. Lond. B. Biol. Sci., 369, 20130458 (2014)[PubMed]
- Nakazawa, Y., Ariyoshi, T., Noga, A. et al.: Space-dependent formation of central pair microtubules and their interactions with radial spokes. PLoS One 9, e110513 (2014)[PubMed]
- Nigg, E. A. & Raff, J. W.: Centrioles, centrosomes, and cilia in health and disease. Cell, 139, 663-678 (2009)[PubMed]
- Matsuura, K., Lefebvre, P. A., Kamiya, R. et al.: Bld10p, a novel protein essential for basal body assembly in Chlamydomonas: localization to the cartwheel, the first ninefold symmetrical structure appearing during assembly. J. Cell Biol., 165, 663-671 (2004)[PubMed]
- Nakazawa, Y., Hiraki, M., Kamiya, R. et al.: SAS-6 is a cartwheel protein that establishes the 9-fold symmetry of the centriole. Curr. Biol., 17, 2169-2174 (2007)[PubMed]
- Hiraki, M., Nakazawa, Y., Kamiya, R. et al.: Bld10p constitutes the cartwheel-spoke tip and stabilizes the 9-fold symmetry of the centriole. Curr. Biol., 17, 1778-1783 (2007)[PubMed]
- van Breugel, M., Hirono, M., Andreeva, A. et al.: Structures of SAS-6 suggest its organization in centrioles. Science, 331, 1196-1199 (2011)[PubMed]
- Kitagawa, D., Vakonakis, I., Olieric, N. et al.: Structural basis of the 9-fold symmetry of centrioles. Cell, 144, 364-375 (2011)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:東京大学大学院理学系研究科博士課程 在学中.
廣野 雅文(Masafumi Hirono)
法政大学生命科学部 教授.
© 2016 苗加 彰・廣野雅文 Licensed under CC 表示 2.1 日本
