シロイヌナズナにおいて花粉管の先端に局在する受容体が花粉管の伸長および花粉管誘引ペプチドの感知を制御する
武内秀憲・東山哲也
(科学技術振興機構ERATO東山ライブホロニクスプロジェクト)
email:武内秀憲
DOI: 10.7875/first.author.2016.026
Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis.
Hidenori Takeuchi, Tetsuya Higashiyama
Nature, 531, 245-248 (2016)
多細胞生物において,細胞の成長方向の制御は正常な組織の形成や細胞間コミュニケーションに必須である.被子植物の生殖の過程においては,先端成長する花粉管が雌しべのなかを進み,卵細胞のとなりにある助細胞からの誘引シグナルを感知することで受精の場まで到達する.今回,筆者らは,シロイヌナズナの花粉管誘引ペプチドAtLURE1の感知に必須な花粉管の受容体としてPRK6を同定した.PRK6はほかのPRKファミリーと協調してはたらき,種に特異的なAtLURE1を認識すると考えられた.PRK6は,細胞内シグナル伝達スイッチであるROPの活性化タンパク質であり花粉管の正常な伸長に重要なROPGEFと相互作用した.花粉管がAtLURE1にむかい方向を転換する際にPRK6の局在を観察したところ,伸長の方向を変えるまえに,細胞膜に局在するPRK6はAtLURE1をくわえた側に非対称に蓄積した.これらの結果から,PRK6を中心としたPRKファミリーがAtLURE1を認識し,花粉管の伸長に重要なタンパク質をAtLURE1の方向に集積することにより,正確かつ迅速な花粉管の伸長方向の変化が達成されることが明らかにされた.
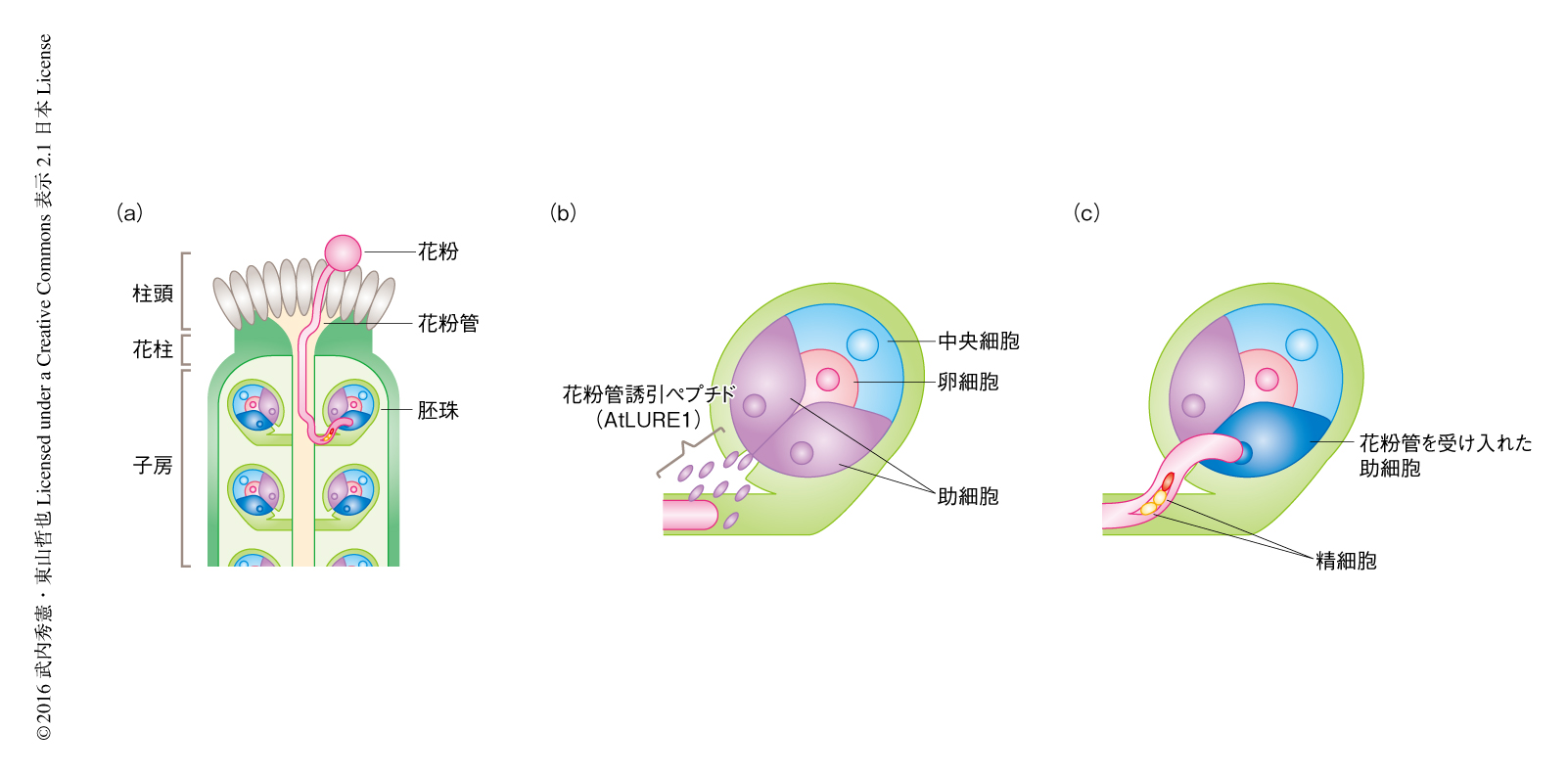
被子植物は同種どうしでの受精を効率よく達成するため,花粉管が雌しべの奥深くにある卵細胞まで精細胞を送り届けるしくみを発達させた1).花粉管は,花粉が柱頭に受粉し発芽することで発達するひとつの細胞からなる雄性配偶体であり,内部に精細胞を含む.花粉管は厳密に制御された先端成長のしくみをもち,雌しべの柱頭,花柱を通過し,胚珠を含む子房の内部へと進入する(図1).胚珠は受精ののち種子へと発達する組織で,内部に卵細胞を含む.卵細胞のとなりにある助細胞からの誘引シグナルにより花粉管は胚珠へと誘引され,精細胞が卵細胞へと受け渡されることで受精が行われる.助細胞から分泌される誘引物質として,双子葉植物のトレニアおよびシロイヌナズナにおいてLUREが2,3),単子葉植物のトウモロコシにおいてZmEA1が同定されていた4).LUREはCRPファミリーに属する約70アミノ酸残基からなる分泌性のペプチドである.トレニアとシロイヌナズナのLUREのあいだにはシステイン残基の共通性はあるものの,全長配列の同一性はほとんどみられない.さらに,近縁な種のあいだでも配列に違いがあり,花粉管の誘引活性も同種に対してより効率的にはたらく3).ひとつの胚珠に1本の花粉管だけを誘引するために重要な花粉管誘引ペプチドの分泌停止のしくみ5)(新着論文レビュー でも掲載)を含め,花粉管誘引ペプチドを分泌する雌しべの側のしくみについては比較的理解が進んでいる.しかしながら,花粉管誘引ペプチドを含む雌しべの組織からのシグナルを花粉管がどのように感知しているのかはほとんどわかっていなかった.
なお,植物における受精については,東山 哲也, 領域融合レビュー, 1, e007, 2012 も参照されたい.
シロイヌナズナAtLURE1の認識および応答にかかわる受容体様キナーゼを探索した.このとき,筆者らによる先行研究から,AtLURE1のほかにも誘引物質が存在し,雌しべの組織からのリガンドと花粉管の受容体との多対多のペアの存在が想像された.そこで,細胞外領域をもつ受容体様キナーゼのなかから,複数のホモログの遺伝子が花粉管において特異的に発現するものに着目した.培地に伸長させた花粉管に精製AtLURE1を含むゼラチンビーズを作用させて応答を調べるsemi-in vivo花粉管誘引アッセイにより,受容体様キナーゼの変異体の花粉管をスクリーニングした.その結果,解析した23の受容体様キナーゼの変異体のうち,prk6変異体のみがAtLURE1にまったく反応しなかった.ほかの7つのprk変異体は正常に反応した.蛍光タンパク質との融合により局在を観察したところ,膜貫通型の受容体様キナーゼであるPRK6はおもに伸長する花粉管の先端の細胞膜に局在した.このことから,PRK6がAtLURE1を感知し,花粉管の先端成長の方向を制御するタンパク質であると考えられた.
花粉管の受容体と外部からのリガンドとの多対多のペアの存在を予想していたことにくわえ,トマトのPRKファミリーにおける先行研究においてLePRK1とLePRK2の相互作用の状態の変化が花粉管の発芽および伸長を制御することが示唆されていた6).そこで,複数のPRKファミリーが花粉管の伸長およびAtLURE1の感知を制御していると考え,prk多重変異体を作製した.多数の変異体を同時に解析するにあたり,AtLURE1をくわえた培地に花粉管を伸長させることにより花粉管の伸長およびAtLURE1への反応性を同時にみるという,簡便で用量に依存的なsemi-in vivoアッセイを新たに用いた.このアッセイでは,野生型の花粉管はうねうねと波うった伸長パターンを示したが,prk6変異体の花粉管はAtLURE1を含まない培地と同様に真っすぐ伸長した.PRK6と系統的にもっとも近いPRK3に対する変異を含めたprk3 prk6二重変異体,さらに,同じサブクレードに属するPRK8に対する変異を含めたprk3 prk6 prk8三重変異体は,prk6変異によりAtLURE1を感知できないことにくわえ花粉管の伸長の異常を示した.PRK6とは異なるサブクレードに属するPRK1およびPRK3に対するprk1 prk3二重変異体の花粉管は正常に伸長したが,AtLURE1への応答性が低下していた.prk1 prk3 prk6三重変異体はprk3 prk6二重変異体より重篤な花粉管の伸長の異常を示した.このsemi-in vivoアッセイにより,AtLURE1の感知および花粉管の伸長は複数のPRKファミリーとPRK6とが協調的にはたらくことにより制御されることが明らかにされた.
雌しべにおけるPRKファミリーの機能について調べた.野生型と比べ,prk6変異体の花粉管は胚珠への誘引に若干の乱れが観察された.この弱い表現型は,AtLURE1をノックダウンした胚珠に対し低頻度で花粉管の誘引の異常がみられたことと矛盾しなかった3).prk3 prk6二重変異体では花粉管の伸長率は低下し胚珠への誘引が遅れた.prk3 prk6 prk8三重変異体およびprk1 prk3 prk6三重変異体においては,花粉管の伸長のより顕著な異常,および,胚珠の近傍まで伸長してきた花粉管に対する誘引頻度の低下が観察され,種子の発達率にも影響がみられた.これらの結果は,雌しべにおいてもPRKファミリーが花粉管の伸長および誘引物質の感知に重要な役割をはたすることを示した.トマトにおける研究から,LePRK2は花粉それ自体や花柱組織から分泌される複数のCRPファミリーのリガンドを受容することにより花粉管の発芽および伸長を制御することが示唆されている7,8).これらのことから,助細胞から分泌されるAtLURE1にくわえ,未同定の花粉管誘引ペプチドや花粉管の伸長の促進にかかわるペプチドなど,多数のリガンドを複数のPRKファミリーが感知することが予想された.
トマトのLePRK1とLePRK2およびシロイヌナズナPRK2についての先行研究により,PRKファミリーは細胞内シグナル伝達スイッチとしてはたらくROPの活性化タンパク質ROPGEF(GEF:guanine nucleotide-exchange factor,グアニンヌクレオチド交換因子)と相互作用することが考えられた.実際に,BiFC(bimolecular fluorescence complementation,2分子蛍光相補性)法により,PRK6は花粉管において発現するROPGEFと相互作用することが示された.PRK6と花粉管で発現するROPGEFのひとつであるROPGEF12との結合領域をさらに探索するため,細胞内領域を段階的に欠失させたPRK6を用いて共免疫沈降実験を行った.タバコの葉における一過的な発現系を用いた実験により,キナーゼ領域を欠失させたPRK6でもROPGEF12との相互作用が確認された一方,膜貫通領域とキナーゼ領域のあいだの膜近傍領域も欠失させたPRK6では相互作用はみられなかった.この結果は,PRK6と花粉管の伸長に中心的な役割をはたすROPGEFとの相互作用にPRK6のキナーゼ領域は必要ないことを示唆した.
同様に,花粉管の伸長およびAtLURE1の感知に対するPRK6の細胞内領域の重要性について,変異体の相補実験により調べた.prk6変異体のAtLURE1非感受性の表現型は,キナーゼ領域を欠失させたPRK6の発現により相補されたが,膜近傍領域も欠失させたPRK6の発現では相補されなかった.prk3 prk6二重変異体における花粉管の伸長の異常の表現型は,全長のPRK6の発現では相補されたが,キナーゼ領域を欠失させたPRK6の発現では相補されなかった.
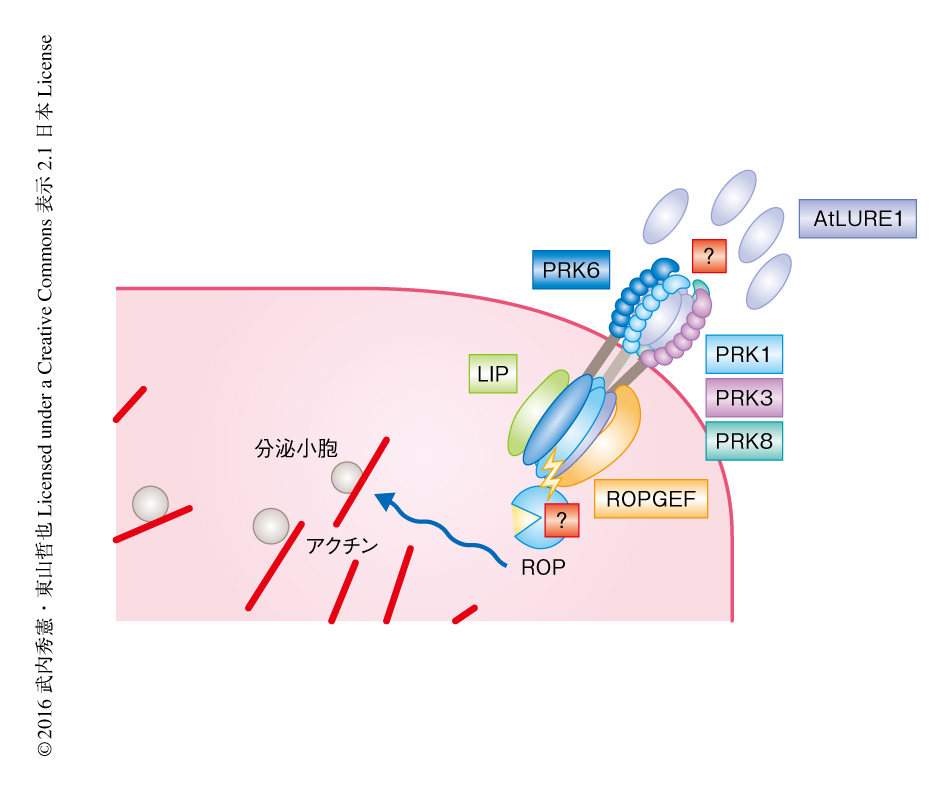
また,BiFC法により,PRK6はPRK6それ自体やPRK3,花粉管の伸長および胚珠への誘引にかかわる花粉管のタンパク質である受容体様細胞質キナーゼLIP1およびLIP2と相互作用しうることが示された.以上の結果より,PRK6はほかのPRKファミリーや細胞内のタンパク質と複合体を形成し,花粉管の伸長に中心的な役割をはたすROPGEFを介することにより,花粉管の伸長およびAtLURE1の感知を細胞内に伝達していると考えられた(図2).
AtLURE1は同じ種であるシロイヌナズナに対し効率的にはたらく3).そこで,同じアブラナ科の近縁種でありゲノム塩基配列が解読されているルベラナズナ(Capsella rubella)を用いて,PRK6がAtLURE1の認識の機能をもつかどうか調べた.野生型のルベラナズナの花粉管はシロイヌナズナのAtLURE1に反応しなかった一方,シロイヌナズナのPRK6を発現させたルベラナズナの花粉管はシロイヌナズナのAtLURE1に高効率で反応した.この結果は,種に特異的にはたらくAtLURE1の認識をPRK6が担うことを強く示唆した.
さらに,ルベラナズナのPRK6をシロイヌナズナのprk変異体に導入し表現型を相補するかどうか調べた.意外なことに,ルベラナズナPRK6の発現はprk6変異体のAtLURE1非感受性の表現型を相補した.しかしながら,prk3 prk6二重変異体などの多重変異体にルベラナズナPRK6を発現させたところ,花粉管の伸長の異常はシロイヌナズナのPRK6と同様に相補したものの,AtLURE1非感受性の表現型は部分的に相補しただけであった.さきに述べたprk1 prk3二重変異体の花粉管においてAtLURE1の感受性が低下していたことを含め,ここまでの結果から,PRK6はほかのPRKファミリーと協調して種に特異的なAtLURE1を認識することが示された(図2).
PRK6は花粉管の先端の細胞膜に集中して局在していた.花粉管の先端部では活発なエクソサイトーシスやエンドサイトーシスにより先端成長が制御されている.そこで,外部からのAtLURE1のシグナルにより花粉管が伸長方向を変化させる際,花粉管の先端のPRK6がどのように挙動するかを,semi-in vivo花粉管誘引アッセイとタイムラプス観察を組み合わせて解析した.その結果,花粉管が真っすぐに伸長している際には蛍光タンパク質で標識したPRK6は花粉管の先端の細胞膜に左右均等に局在した.AtLURE1を含むゼラチンビーズを花粉管の片側に作用させたところ,花粉管がビーズをおいた位置にむかい伸長方向を変化させるまえに,PRK6がビーズをおいた側に非対称に蓄積するようすが観察された.花粉管の先端部においてPRK6の局在がかたよったのち,花粉管はその方向にむかいふくらんでいき,先端の伸長方向が変化した.このことから,外部からのAtLURE1のシグナルにより花粉管の伸長に重要なタンパク質と相互作用するPRK6の局在が制御されることで,花粉管の伸長方向の変化がひき起こされると考えられた(図2).
この研究により,PRK6を中心とした複数のPRKファミリーがシロイヌナズナの花粉管誘引ペプチドAtLURE1の認識に必須であることが明らかにされた.中国の研究グループは,この論文と連報で,AtLURE1へのノックアウト変異体の応答欠損の表現型は弱いものの,AtLURE1と結合するMDIS1およびMIK1とMIK2と名づけられた受容体の同定を報告した9).MDIS1は花粉管において高発現しており,筆者らの解析の候補にもあった.MIK1およびMIK2はそれぞれLRR XIおよびLRR XIIの異なるサブファミリーに属する受容体であるが,二重変異体の解析,および,AtLURE1に依存的にそれぞれMDIS1とヘテロ二量体を形成するという結果が報告された.筆者らは,AtLURE1は誘引活性に重要な塩基性の領域をもつことを示しており,その領域を介した粘着的な相互作用がさまたげとなりAtLURE1とPRKファミリーとの特異的な結合を示すにはいたっていない.今後,AtLURE1とPRKファミリーとの相互作用のさらなる解析や,PRKファミリーとMDIS1およびMIKとの関係を調べることにより,AtLURE1の受容機構が明らかにされると考えられる.また,AtLURE1のほかにも存在するであろう誘引物質を同定し,これら受容体との関係を調べる必要もある.
今回,ROPGEFなど花粉管の伸長に重要なタンパク質と相互作用するPRKファミリーがAtLURE1の感知に必須であることが示された.AtLURE1の受容によりROPシグナル伝達系が活性化されるかどうかは明らかにされなかったが,伸長方向の変化のまえにPRK6が非対称に局在することは興味深い.AtLURE1による花粉管の内部におけるタンパク質の活性化だけでなく,PRK6の局在の制御が細胞外からの誘引シグナルの微小な変化を感知するため重要なしくみなのかもしれない.AtLURE1と花粉管の先端における受容体の挙動や細胞内シグナル伝達タンパク質の活性化をリアルタイムで解析することにより,活発に先端成長する花粉管における方向の制御の分子機構が明らかにされると考えられる.
略歴:2013年 名古屋大学大学院理学研究科 修了,同年 科学技術振興機構ERATO東山ライブホロニクスプロジェクト 研究員を経て,2016年よりオーストリアGregor Mendel Institute研究員.
東山 哲也(Tetsuya Higashiyama)
名古屋大学トランスフォーマティブ生命分子研究所 教授.
研究室URL:http://www.higashiyama-lab.com
© 2016 武内秀憲・東山哲也 Licensed under CC 表示 2.1 日本
(科学技術振興機構ERATO東山ライブホロニクスプロジェクト)
email:武内秀憲
DOI: 10.7875/first.author.2016.026
Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis.
Hidenori Takeuchi, Tetsuya Higashiyama
Nature, 531, 245-248 (2016)
要 約
多細胞生物において,細胞の成長方向の制御は正常な組織の形成や細胞間コミュニケーションに必須である.被子植物の生殖の過程においては,先端成長する花粉管が雌しべのなかを進み,卵細胞のとなりにある助細胞からの誘引シグナルを感知することで受精の場まで到達する.今回,筆者らは,シロイヌナズナの花粉管誘引ペプチドAtLURE1の感知に必須な花粉管の受容体としてPRK6を同定した.PRK6はほかのPRKファミリーと協調してはたらき,種に特異的なAtLURE1を認識すると考えられた.PRK6は,細胞内シグナル伝達スイッチであるROPの活性化タンパク質であり花粉管の正常な伸長に重要なROPGEFと相互作用した.花粉管がAtLURE1にむかい方向を転換する際にPRK6の局在を観察したところ,伸長の方向を変えるまえに,細胞膜に局在するPRK6はAtLURE1をくわえた側に非対称に蓄積した.これらの結果から,PRK6を中心としたPRKファミリーがAtLURE1を認識し,花粉管の伸長に重要なタンパク質をAtLURE1の方向に集積することにより,正確かつ迅速な花粉管の伸長方向の変化が達成されることが明らかにされた.
はじめに
被子植物は同種どうしでの受精を効率よく達成するため,花粉管が雌しべの奥深くにある卵細胞まで精細胞を送り届けるしくみを発達させた1).花粉管は,花粉が柱頭に受粉し発芽することで発達するひとつの細胞からなる雄性配偶体であり,内部に精細胞を含む.花粉管は厳密に制御された先端成長のしくみをもち,雌しべの柱頭,花柱を通過し,胚珠を含む子房の内部へと進入する(図1).胚珠は受精ののち種子へと発達する組織で,内部に卵細胞を含む.卵細胞のとなりにある助細胞からの誘引シグナルにより花粉管は胚珠へと誘引され,精細胞が卵細胞へと受け渡されることで受精が行われる.助細胞から分泌される誘引物質として,双子葉植物のトレニアおよびシロイヌナズナにおいてLUREが2,3),単子葉植物のトウモロコシにおいてZmEA1が同定されていた4).LUREはCRPファミリーに属する約70アミノ酸残基からなる分泌性のペプチドである.トレニアとシロイヌナズナのLUREのあいだにはシステイン残基の共通性はあるものの,全長配列の同一性はほとんどみられない.さらに,近縁な種のあいだでも配列に違いがあり,花粉管の誘引活性も同種に対してより効率的にはたらく3).ひとつの胚珠に1本の花粉管だけを誘引するために重要な花粉管誘引ペプチドの分泌停止のしくみ5)(新着論文レビュー でも掲載)を含め,花粉管誘引ペプチドを分泌する雌しべの側のしくみについては比較的理解が進んでいる.しかしながら,花粉管誘引ペプチドを含む雌しべの組織からのシグナルを花粉管がどのように感知しているのかはほとんどわかっていなかった.
なお,植物における受精については,東山 哲也, 領域融合レビュー, 1, e007, 2012 も参照されたい.
1.花粉管の先端に局在する受容体様キナーゼPRK6はAtLURE1の感知に必須である
シロイヌナズナAtLURE1の認識および応答にかかわる受容体様キナーゼを探索した.このとき,筆者らによる先行研究から,AtLURE1のほかにも誘引物質が存在し,雌しべの組織からのリガンドと花粉管の受容体との多対多のペアの存在が想像された.そこで,細胞外領域をもつ受容体様キナーゼのなかから,複数のホモログの遺伝子が花粉管において特異的に発現するものに着目した.培地に伸長させた花粉管に精製AtLURE1を含むゼラチンビーズを作用させて応答を調べるsemi-in vivo花粉管誘引アッセイにより,受容体様キナーゼの変異体の花粉管をスクリーニングした.その結果,解析した23の受容体様キナーゼの変異体のうち,prk6変異体のみがAtLURE1にまったく反応しなかった.ほかの7つのprk変異体は正常に反応した.蛍光タンパク質との融合により局在を観察したところ,膜貫通型の受容体様キナーゼであるPRK6はおもに伸長する花粉管の先端の細胞膜に局在した.このことから,PRK6がAtLURE1を感知し,花粉管の先端成長の方向を制御するタンパク質であると考えられた.
2.PRKファミリーは花粉管の伸長とAtLURE1の感知を協調して制御する
花粉管の受容体と外部からのリガンドとの多対多のペアの存在を予想していたことにくわえ,トマトのPRKファミリーにおける先行研究においてLePRK1とLePRK2の相互作用の状態の変化が花粉管の発芽および伸長を制御することが示唆されていた6).そこで,複数のPRKファミリーが花粉管の伸長およびAtLURE1の感知を制御していると考え,prk多重変異体を作製した.多数の変異体を同時に解析するにあたり,AtLURE1をくわえた培地に花粉管を伸長させることにより花粉管の伸長およびAtLURE1への反応性を同時にみるという,簡便で用量に依存的なsemi-in vivoアッセイを新たに用いた.このアッセイでは,野生型の花粉管はうねうねと波うった伸長パターンを示したが,prk6変異体の花粉管はAtLURE1を含まない培地と同様に真っすぐ伸長した.PRK6と系統的にもっとも近いPRK3に対する変異を含めたprk3 prk6二重変異体,さらに,同じサブクレードに属するPRK8に対する変異を含めたprk3 prk6 prk8三重変異体は,prk6変異によりAtLURE1を感知できないことにくわえ花粉管の伸長の異常を示した.PRK6とは異なるサブクレードに属するPRK1およびPRK3に対するprk1 prk3二重変異体の花粉管は正常に伸長したが,AtLURE1への応答性が低下していた.prk1 prk3 prk6三重変異体はprk3 prk6二重変異体より重篤な花粉管の伸長の異常を示した.このsemi-in vivoアッセイにより,AtLURE1の感知および花粉管の伸長は複数のPRKファミリーとPRK6とが協調的にはたらくことにより制御されることが明らかにされた.
3.PRKファミリーは雌しべにおける花粉管の伸長および胚珠への誘引に重要な役割をはたす
雌しべにおけるPRKファミリーの機能について調べた.野生型と比べ,prk6変異体の花粉管は胚珠への誘引に若干の乱れが観察された.この弱い表現型は,AtLURE1をノックダウンした胚珠に対し低頻度で花粉管の誘引の異常がみられたことと矛盾しなかった3).prk3 prk6二重変異体では花粉管の伸長率は低下し胚珠への誘引が遅れた.prk3 prk6 prk8三重変異体およびprk1 prk3 prk6三重変異体においては,花粉管の伸長のより顕著な異常,および,胚珠の近傍まで伸長してきた花粉管に対する誘引頻度の低下が観察され,種子の発達率にも影響がみられた.これらの結果は,雌しべにおいてもPRKファミリーが花粉管の伸長および誘引物質の感知に重要な役割をはたすることを示した.トマトにおける研究から,LePRK2は花粉それ自体や花柱組織から分泌される複数のCRPファミリーのリガンドを受容することにより花粉管の発芽および伸長を制御することが示唆されている7,8).これらのことから,助細胞から分泌されるAtLURE1にくわえ,未同定の花粉管誘引ペプチドや花粉管の伸長の促進にかかわるペプチドなど,多数のリガンドを複数のPRKファミリーが感知することが予想された.
4.PRK6は花粉管の伸長に重要な役割をはたすタンパク質と相互作用する
トマトのLePRK1とLePRK2およびシロイヌナズナPRK2についての先行研究により,PRKファミリーは細胞内シグナル伝達スイッチとしてはたらくROPの活性化タンパク質ROPGEF(GEF:guanine nucleotide-exchange factor,グアニンヌクレオチド交換因子)と相互作用することが考えられた.実際に,BiFC(bimolecular fluorescence complementation,2分子蛍光相補性)法により,PRK6は花粉管において発現するROPGEFと相互作用することが示された.PRK6と花粉管で発現するROPGEFのひとつであるROPGEF12との結合領域をさらに探索するため,細胞内領域を段階的に欠失させたPRK6を用いて共免疫沈降実験を行った.タバコの葉における一過的な発現系を用いた実験により,キナーゼ領域を欠失させたPRK6でもROPGEF12との相互作用が確認された一方,膜貫通領域とキナーゼ領域のあいだの膜近傍領域も欠失させたPRK6では相互作用はみられなかった.この結果は,PRK6と花粉管の伸長に中心的な役割をはたすROPGEFとの相互作用にPRK6のキナーゼ領域は必要ないことを示唆した.
同様に,花粉管の伸長およびAtLURE1の感知に対するPRK6の細胞内領域の重要性について,変異体の相補実験により調べた.prk6変異体のAtLURE1非感受性の表現型は,キナーゼ領域を欠失させたPRK6の発現により相補されたが,膜近傍領域も欠失させたPRK6の発現では相補されなかった.prk3 prk6二重変異体における花粉管の伸長の異常の表現型は,全長のPRK6の発現では相補されたが,キナーゼ領域を欠失させたPRK6の発現では相補されなかった.
また,BiFC法により,PRK6はPRK6それ自体やPRK3,花粉管の伸長および胚珠への誘引にかかわる花粉管のタンパク質である受容体様細胞質キナーゼLIP1およびLIP2と相互作用しうることが示された.以上の結果より,PRK6はほかのPRKファミリーや細胞内のタンパク質と複合体を形成し,花粉管の伸長に中心的な役割をはたすROPGEFを介することにより,花粉管の伸長およびAtLURE1の感知を細胞内に伝達していると考えられた(図2).
5.PRK6は種に特異的なAtLURE1の認識に中心的な役割をはたす
AtLURE1は同じ種であるシロイヌナズナに対し効率的にはたらく3).そこで,同じアブラナ科の近縁種でありゲノム塩基配列が解読されているルベラナズナ(Capsella rubella)を用いて,PRK6がAtLURE1の認識の機能をもつかどうか調べた.野生型のルベラナズナの花粉管はシロイヌナズナのAtLURE1に反応しなかった一方,シロイヌナズナのPRK6を発現させたルベラナズナの花粉管はシロイヌナズナのAtLURE1に高効率で反応した.この結果は,種に特異的にはたらくAtLURE1の認識をPRK6が担うことを強く示唆した.
さらに,ルベラナズナのPRK6をシロイヌナズナのprk変異体に導入し表現型を相補するかどうか調べた.意外なことに,ルベラナズナPRK6の発現はprk6変異体のAtLURE1非感受性の表現型を相補した.しかしながら,prk3 prk6二重変異体などの多重変異体にルベラナズナPRK6を発現させたところ,花粉管の伸長の異常はシロイヌナズナのPRK6と同様に相補したものの,AtLURE1非感受性の表現型は部分的に相補しただけであった.さきに述べたprk1 prk3二重変異体の花粉管においてAtLURE1の感受性が低下していたことを含め,ここまでの結果から,PRK6はほかのPRKファミリーと協調して種に特異的なAtLURE1を認識することが示された(図2).
6.外部からのAtLURE1のシグナルに応答してPRK6が花粉管の伸長方向を標識する
PRK6は花粉管の先端の細胞膜に集中して局在していた.花粉管の先端部では活発なエクソサイトーシスやエンドサイトーシスにより先端成長が制御されている.そこで,外部からのAtLURE1のシグナルにより花粉管が伸長方向を変化させる際,花粉管の先端のPRK6がどのように挙動するかを,semi-in vivo花粉管誘引アッセイとタイムラプス観察を組み合わせて解析した.その結果,花粉管が真っすぐに伸長している際には蛍光タンパク質で標識したPRK6は花粉管の先端の細胞膜に左右均等に局在した.AtLURE1を含むゼラチンビーズを花粉管の片側に作用させたところ,花粉管がビーズをおいた位置にむかい伸長方向を変化させるまえに,PRK6がビーズをおいた側に非対称に蓄積するようすが観察された.花粉管の先端部においてPRK6の局在がかたよったのち,花粉管はその方向にむかいふくらんでいき,先端の伸長方向が変化した.このことから,外部からのAtLURE1のシグナルにより花粉管の伸長に重要なタンパク質と相互作用するPRK6の局在が制御されることで,花粉管の伸長方向の変化がひき起こされると考えられた(図2).
おわりに
この研究により,PRK6を中心とした複数のPRKファミリーがシロイヌナズナの花粉管誘引ペプチドAtLURE1の認識に必須であることが明らかにされた.中国の研究グループは,この論文と連報で,AtLURE1へのノックアウト変異体の応答欠損の表現型は弱いものの,AtLURE1と結合するMDIS1およびMIK1とMIK2と名づけられた受容体の同定を報告した9).MDIS1は花粉管において高発現しており,筆者らの解析の候補にもあった.MIK1およびMIK2はそれぞれLRR XIおよびLRR XIIの異なるサブファミリーに属する受容体であるが,二重変異体の解析,および,AtLURE1に依存的にそれぞれMDIS1とヘテロ二量体を形成するという結果が報告された.筆者らは,AtLURE1は誘引活性に重要な塩基性の領域をもつことを示しており,その領域を介した粘着的な相互作用がさまたげとなりAtLURE1とPRKファミリーとの特異的な結合を示すにはいたっていない.今後,AtLURE1とPRKファミリーとの相互作用のさらなる解析や,PRKファミリーとMDIS1およびMIKとの関係を調べることにより,AtLURE1の受容機構が明らかにされると考えられる.また,AtLURE1のほかにも存在するであろう誘引物質を同定し,これら受容体との関係を調べる必要もある.
今回,ROPGEFなど花粉管の伸長に重要なタンパク質と相互作用するPRKファミリーがAtLURE1の感知に必須であることが示された.AtLURE1の受容によりROPシグナル伝達系が活性化されるかどうかは明らかにされなかったが,伸長方向の変化のまえにPRK6が非対称に局在することは興味深い.AtLURE1による花粉管の内部におけるタンパク質の活性化だけでなく,PRK6の局在の制御が細胞外からの誘引シグナルの微小な変化を感知するため重要なしくみなのかもしれない.AtLURE1と花粉管の先端における受容体の挙動や細胞内シグナル伝達タンパク質の活性化をリアルタイムで解析することにより,活発に先端成長する花粉管における方向の制御の分子機構が明らかにされると考えられる.
文 献
- Higashiyama, T. & Takeuchi, H.: The mechanism and key molecules involved in pollen tube guidance. Annu. Rev. Plant Biol., 66, 393-413 (2015)[PubMed]
- Okuda, S., Tsutsui, H., Shiina, K. et al.: Defensin-like polypeptide LUREs are pollen tube attractants secreted from synergid cells. Nature, 458, 357-361 (2009)[PubMed]
- Takeuchi, H. & Higashiyama, T.: A species-specific cluster of defensin-like genes encodes diffusible pollen tube attractants in Arabidopsis. PLoS Biol., 10, e1001449 (2012)[PubMed]
- Marton, M. L., Fastner, A., Uebler, S. et al.: Overcoming hybridization barriers by the secretion of the maize pollen tube attractant ZmEA1 from Arabidopsis ovules. Curr. Biol., 22, 1194-1198 (2012)[PubMed]
- Maruyama, D., Volz, R., Takeuchi, H. et al.: Rapid elimination of the persistent synergid through a cell fusion mechanism. Cell, 161, 907-918 (2015)[PubMed] [新着論文レビュー]
- Wengier, D., Valsecchi, I., Cabanas, M. L. et al.: The receptor kinases LePRK1 and LePRK2 associate in pollen and when expressed in yeast, but dissociate in the presence of style extract. Proc. Natl. Acad. Sci. USA, 100, 6860-6865 (2003)[PubMed]
- Tang, W., Ezcurra, I., Muschietti, J. et al.: A cysteine-rich extracellular protein, LAT52, interacts with the extracellular domain of the pollen receptor kinase LePRK2. Plant Cell, 14, 2277-2287 (2002)[PubMed]
- Tang, W., Kelley, D., Ezcurra, I. et al.: LeSTIG1, an extracellular binding partner for the pollen receptor kinases LePRK1 and LePRK2, promotes pollen tube growth in vitro. Plant J., 39, 343-353 (2004)[PubMed]
- Wang, T., Liang. L., Xue, Y. et al.: A receptor heteromer mediates the male perception of female attractants in plants. Nature, 531, 241-244 (2016)[PubMed]
著者プロフィール
略歴:2013年 名古屋大学大学院理学研究科 修了,同年 科学技術振興機構ERATO東山ライブホロニクスプロジェクト 研究員を経て,2016年よりオーストリアGregor Mendel Institute研究員.
東山 哲也(Tetsuya Higashiyama)
名古屋大学トランスフォーマティブ生命分子研究所 教授.
研究室URL:http://www.higashiyama-lab.com
© 2016 武内秀憲・東山哲也 Licensed under CC 表示 2.1 日本
