円口類から解き明かされる脳の領域化の進化的な起源
菅原文昭・倉谷 滋
(理化学研究所 倉谷形態進化研究室)
email:菅原文昭,倉谷 滋
DOI: 10.7875/first.author.2016.015
Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain.
Fumiaki Sugahara, Juan Pascual-Anaya, Yasuhiro Oisi, Shigehiro Kuraku, Shin-ichi Aota, Noritaka Adachi, Wataru Takagi, Tamami Hirai, Noboru Sato, Yasunori Murakami, Shigeru Kuratani
Nature, 531, 97-100 (2016)
脊椎動物の脳は複雑かつ精密に領域化された器官であり,おのおのの領域が進化の過程においていつ獲得されたのかについては不明な点が多い.今回,筆者らは,現生の脊椎動物のうち最初に分岐した,顎のない円口類のうちヌタウナギの脳の発生の過程を世界ではじめて詳細に観察し,とくに,脳室の形態,神経線維の走行,遺伝子の発現パターンから脳のおのおのの領域の同定を試みた.円口類のもうひとつの系統であるヤツメウナギも同時に解析した結果,これまで円口類にはないとされてきた大脳基底核の一部,および,小脳の発生する領域が,じつは存在することが明らかにされた.これにより,脊椎動物の基本的な領域をつくるしくみは,円口類が分岐してから軟骨魚類の分岐までに獲得されたとするこれまでの考えとは異なり,円口類の分岐よりまえ,すなわち,脊椎動物がはじめて誕生した5億年以上まえに成立したことがわかった.
脊椎動物の誕生は古生代カンブリア紀ごろと考えられている.そのころの脊椎動物の脳がどのようなものであったのかを想像するのはむずかしい.脳は軟組織であるため化石として残りにくく,絶滅した動物の化石からわかることはかぎられるためである.一方,現生の脊椎動物でもっとも古くに分岐したのは顎のない“円口類”で,ヌタウナギおよびヤツメウナギの2つの群のみが現存する.この円口類は5億年以上まえに顎のある脊椎動物“顎口類”とたもとを分かった系統なので,円口類と顎口類とを比較しそれらに共通の要素があれば,それは脊椎動物の共通祖先がもっていた形質であるとみなせる.このような方法により発生期の脳を調べ,脳をつくる遺伝子の発現の様式を円口類と顎口類とで比較すれば,脳の祖先的な状態およびその変遷をある程度まで推定することができる.
これまで,円口類の発生の研究にはおもにヤツメウナギが使用されてきた.これは,繁殖期のヤツメウナギが比較的容易に入手でき,受精卵を得るのも可能だからである.先行研究において,ヤツメウナギと顎口類の脳の発生における最大の違いは,1)ヤツメウナギの胚は大脳(終脳)のもっとも腹側の領域である内側基底核隆起をもたない,2)ヤツメウナギの胚は小脳の発生の場となる菱脳唇をもたない,の2つの点であるといわれていた.
1)について,顎口類の内側側基底核隆起は大脳基底核の一部である淡蒼球に分化し運動の制御にかかわる.さらに,内側基底核隆起からGABA作動性抑制ニューロンが生じ大脳皮質の介在ニューロンになる.つまり,内側基底核隆起は特定の領域あるいはニューロンの発生のための原基としてはたらくことから,大脳の皮質および基底核が機能するうえで欠かせない存在なのである.ところが,これまでの研究において,ヤツメウナギの胚には内側基底核隆起を特異化するNkx2.1遺伝子およびShh遺伝子の相同遺伝子の発現がみられず,また,成体においても淡蒼球および皮質においてGABA作動性の介在ニューロンはみつかっていなかった1).Nkx2.1ノックアウトマウスは淡蒼球を欠き,皮質のGABA作動性の介在ニューロンもいちじるしく減少していることから,Nkx2.1ノックアウトマウスはヤツメウナギの“表現型模写”あるいは“先祖返り”の状態を示すと理解されていた.
2)について,小脳は体性感覚や平衡感覚を受容し運動を制御する部分であるが,円口類においてはこの部分が未発達である.つまり,ヌタウナギには小脳がなく,ヤツメウナギにおいては未発達な小脳と一部の交連繊維がみられるにとどまる.顎口類の小脳は,後脳(その形態から,“菱脳”ともよばれる)の背側部である“菱脳唇”の前方から生じ,この領域はPax6遺伝子やWnt1遺伝子などの発現により特徴づけられる.ところが,ヤツメウナギの胚には後脳の背側においてPax6遺伝子の発現がみられないといわれてきた2).Pax6遺伝子は小脳の顆粒細胞層の分化および移動に必須の遺伝子でもあるので,ヤツメウナギは小脳が発生する場としての菱脳唇が進化的に未分化な状態を保持していると解釈されてきた.
以上の知見から,内側基底核隆起および菱脳唇という2つの発生における原基は,円口類と顎口類が分岐したのち,顎口類の進化の過程において新たに獲得され発達したと考えられてきたのである.
しかし,近年,この考えに対しいくつかの疑問や反論が提示された.ひとつは,ヤツメウナギの進化の過程において内側基底核隆起および菱脳唇は2次的に退化したのではないか,という疑問である3).当然ながら,ヤツメウナギは“原始的”な動物ではなく,われわれと同じ5億年の進化をへてきているので,この過程において脳の2次的な改変や退化は起こりうる.もうひとつは,ヤツメウナギの成体において淡蒼球に類似した領域,および,皮質に抗GABA抗体に陽性を示す細胞が観察されたという報告である4,5).これらは顎口類においては内側基底核隆起から分化するため,ヤツメウナギの胚にも未発見の内側基底核隆起が存在すると考えるのが自然であろうが,ヤツメウナギにおいては顎口類とは異なる発生の過程をへてこれらの領域および細胞が生じているという可能性も否定できない.
これらの疑問のうち,前者については,円口類のもうひとつの系統であるヌタウナギを解析する必要が指摘されてきた.しかし,近年まで,ヌタウナギの発生の過程は謎につつまれていた.というのも,ヌタウナギはほとんどの種が深海に生息し,受精卵の入手がきわめて困難だからである.しかし,筆者らのグループは,2007年に世界ではじめて日本産ヌタウナギ(Eptatretus burgeri,図1a)の受精卵を人工的に得ることに成功し,以来,毎年,少数ではあるが胚が得られ,いまでは多くの発生学的な知見が得られる状況になっている6).また,後者については,ヤツメウナギの胚における内側基底核隆起の有無の再検証が必要と考えた.

2011年に島根県沖で捕獲したヌタウナギから得た受精卵のうち,おもに2つを使用し解析した.まず,ヌタウナギの脳の発生の形態的な観察および記載を行った.ヌタウナギの脳の発生過程の記載については,19世紀末および20世紀初頭にかけて偶発的に得られた胚により断片的な報告があるが7,8),これらはみな,硬い卵殻がついたままの卵をエタノール固定したせいで胚がいちじるしく変形しており,正しい形態やおのおのの部位の同定が困難であった.そこで,正しく固定した胚のパラフィン切片から立体再構築像を作成し,脳の形態的な発生を観察し記載した.この結果,ほかの脊椎動物とはきわめて異なった外形を示し,内部も脳室がほぼ消失するなど非常に特異な形態を示すヌタウナギの脳(図1b)も,その発生の過程においては脊椎動物としての基本的な形態を踏襲していることがわかった.
ヌタウナギより脳のおのおのの領域に特徴的に発現するFoxG1遺伝子,Emx1/2遺伝子,Pax6遺伝子,Nkx2.1遺伝子,Shh遺伝子の相同遺伝子をクローニングし,その発現のパターンを観察した.また,神経染色により初期の神経回路および交連繊維を同定し,その位置関係などを手がかりとして脳のおのおのの領域の同定を試みた.この結果,大脳(外套,外套下部),間脳(前視床,視床,視蓋前域,視床下部など),中脳,後脳といった顎口類のもつ基本的な脳の領域が,ヌタウナギにも存在することがわかった(図2a).ただし,松果体は成体のヌタウナギの脳にみられないが,発生の過程においてもこれを見い出すことはできなかった.だからといって,これが原始的な特徴を示しているわけではない.なぜならば,ヤツメウナギは発達した松果体をもつからである.つまり,松果体はヌタウナギの系統において2次的に失われた可能性が高い.
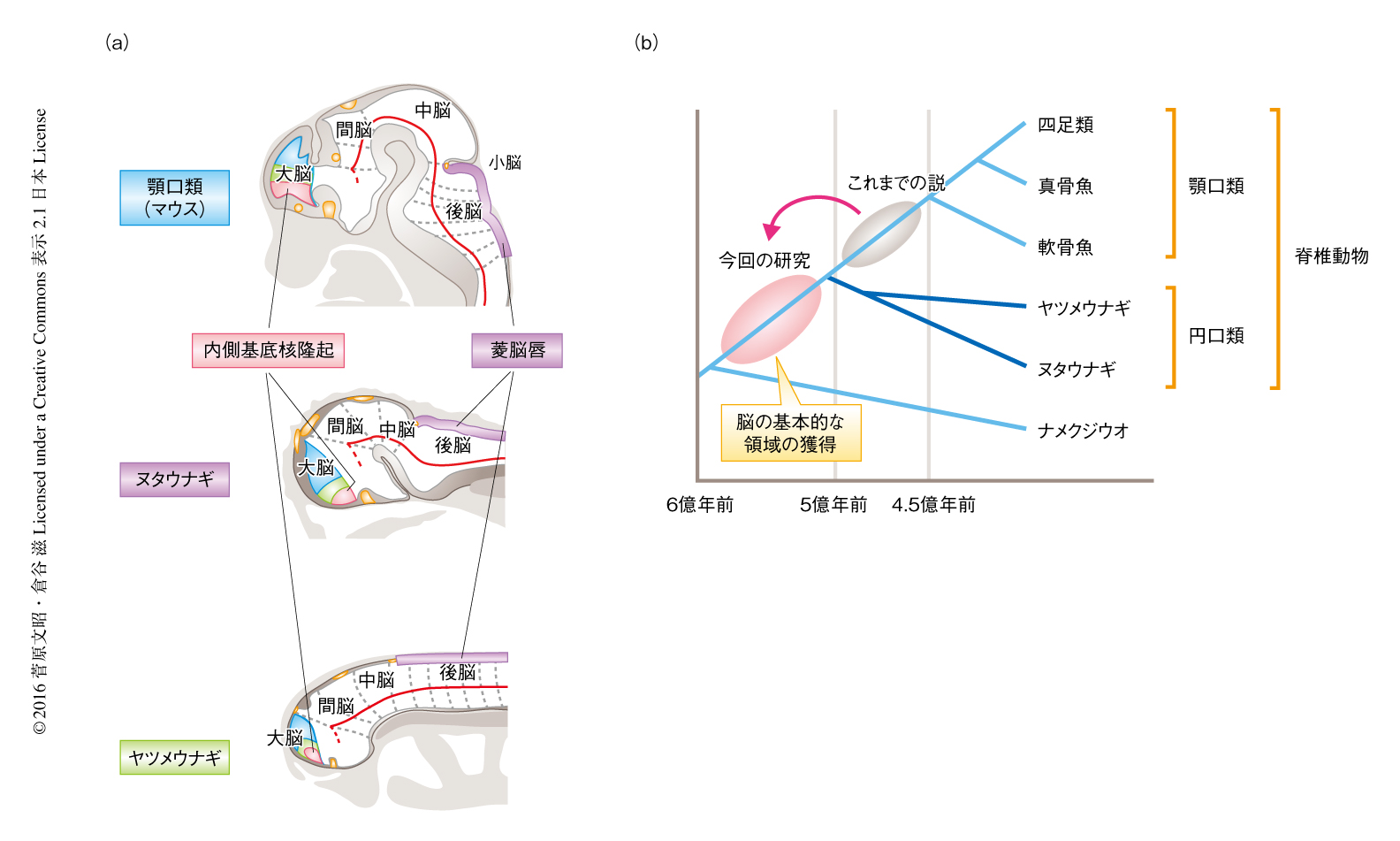
ヌタウナギの胚において内側基底核隆起および菱脳唇の有無について調べた.転写因子Nkx2.1は顎口類においてShhと協調して内側基底核隆起を特異化する.この両者の相同遺伝子の発現を調べた結果,ともにヌタウナギの胚の終脳腹側において発現が観察された.さらに,顎口類において菱脳唇に発現するPax6遺伝子およびAtoh1遺伝子の発現もヌタウナギの後脳背側においてみられた.すなわち,ヌタウナギの脳には,顎口類と同様に,内側基底核隆起および菱脳唇が存在することがわかった(図2a).これは,ヤツメウナギから得られた知見とは大きく異なっていた.
これらの結果から,つぎの2つの進化のシナリオが考えられた.1)ヌタウナギおよび顎口類においてみられた内側基底核隆起および菱脳唇は共通祖先の段階で獲得されたが,ヤツメウナギの系統においてこの2つの領域は2次的に退化した.2)共通祖先の段階において内側基底核隆起および菱脳唇は獲得されておらず,ヌタウナギおよび顎口類においてそれぞれ独立に獲得された,つまり,収斂進化が起こった.これに対し,ヤツメウナギは祖先的な状態を保持している.
これらのシナリオのどちらであるかを検証するため,日本産カワヤツメ(Lethenteron japonicum)の胚を用いて再検証した.近年,公開されたカワヤツメのゲノム塩基配列を使用し9),既知であったNkx2.1遺伝子の相同遺伝子であるNkx2.1/2.4A遺伝子にくわえ,パラログ遺伝子としてNkx2.1/2.4B遺伝子およびNkx2.1/2.4C遺伝子を新規にクローニングした.パラログ遺伝子とは,遺伝子の重複により生じた複数の遺伝子のことである.これらの遺伝子の発現を発生後期のヤツメウナギの胚において観察したところ,既知のNkx2.1/2.4A遺伝子とは異なり,新規のNkx2.1/2.4B遺伝子およびNkx2.1/2.4C遺伝子の発現が大脳腹側において観察された.さらに,菱脳唇についても,既知であったPax6相同遺伝子にくわえPax6B遺伝子を新規にクローニングしたところ,Pax6B遺伝子も後脳背側において発現がみられた.つまり,ヤツメウナギにはないとされていた内側基底核隆起および菱脳唇はいずれも存在しているらしかった(図2a).先行研究においては,ゲノム情報が不足していたことから円口類に何個のパラログ遺伝子があるのか不明だったこともあり,発現していないパラログ遺伝子のみを観察してしまっていたものと考えられた.
ヤツメウナギの後脳においては,Pax6遺伝子にくわえ,Atoh1遺伝子およびPtf1a遺伝子の発現も確認された.Atoh1遺伝子およびPtf1a遺伝子は,それぞれ,顎口類の小脳の層を構成する顆粒細胞およびプルキンエ細胞の分化にきわめて重要である.それらの遺伝子が,形態的に小脳をもたないヤツメウナギの後脳において機能していたことはきわめて興味深い.すなわち,小脳を獲得するまえの段階においてすでにAtoh1遺伝子およびPtf1a遺伝子が菱脳背側の細胞において機能しており,それを下地として顎口類の系統において小脳が獲得されたとみるべきらしい.小脳の前駆体はすべての脊椎動物を産み出した共通祖先に存在していたのだ.
これまで,内側基底核隆起および菱脳唇という2つの脳の領域は,円口類が分岐してから軟骨魚類が分岐する4億5000万年まえのあいだに獲得されたと考えられていた.しかし,今回の研究により,内側基底核隆起および菱脳唇の獲得は円口類と顎口類の共通祖先が存在したころ,すなわち,5億年以上まえにまでさかのぼることになった(図2b).脊椎動物の脳の領域は進化の過程において段階的に獲得されてきたのではなく,脊椎動物の共通祖先の段階ですでに成立していたという新たな進化のシナリオが提示された(図2b).ヒトにも用いられている脳の基本的な発生の構造は,これまで考えられてきたより古い起源をもつらしい.
1960年代より“脳の三位一体説”が提唱され,ヒトの脳は爬虫類脳(大脳基底核),哺乳類原脳(辺縁系),哺乳類脳(新皮質)に分けることができ,これらは進化の過程で段階的に獲得された,と唱えられた10).この説はいまでも一般の脳進化の認識の根底に流れている.しかし,近年の進化発生学の研究による知見および今回の研究により,この理解は変わりつつある.すなわち,脳はまずおおまかな区画をつくり,その区画における複雑化および特異化がそれぞれの動物の系統においてなされたのである.都市の成立の過程にたとえると,市街地が中心部から郊外へと拡大していくのではなく,平城京のようにさきに土地を区分し,そのあとおのおのの区域がにぎわってくる,というイメージだろうか.
この研究において,筆者らは,脊椎動物の初期に脳のおおまかな区画や枠組みの大部分が成立したと論じた.しかしもちろん,脳はヒトにいたるまでにさらなる複雑化をとげている.では,脳のいったいどこがもっともいちじるしく進化したのだろうか? その重要なもののひとつは大脳新皮質の獲得だろう.大脳新皮質は哺乳類においてとくに発達したが,近年,爬虫類や鳥類においても類似した領域がみつかっている.さらには,最近,ヤツメウナギの大脳皮質にも哺乳類の新皮質にある運動野に似た場所のあるらしいことがわかってきた.この大脳新皮質の起源はどこまでさかのぼることができ,そして,どのような進化的な変遷をへてわれわれの“知性”が生まれたのか,非常に魅力的な研究テーマである.また,小脳はすべての顎口類にみられ,体性感覚や平衡感覚を受容し運動を制御しているが,ヒトを含む哺乳類では小脳は大脳新皮質と共同して高度な知覚機能を担うことがわかっている.このような制御中枢としての小脳はどのような発生プログラムの変遷をへて獲得されたのだろうか? このことについては,この研究において小脳の獲得の“前夜”の状態であるとわかった円口類と,小脳をもつ動物のなかでもっとも古くに分岐したサメなどの軟骨魚類との比較が,より詳細な進化の経緯を教えてくれるだろう.
略歴:2011年 神戸大学大学院理学研究科 修了,理化学研究所発生・再生科学総合研究センター 研究員を経て,2013年より兵庫医科大学 講師.
研究テーマ:脊椎動物の頭部の進化発生学.
倉谷 滋(Shigeru Kuratani)
理化学研究所 主任研究員.
研究室URL:http://www.cdb.riken.jp/emo/japanese/indexj.html
© 2016 菅原文昭・倉谷 滋 Licensed under CC 表示 2.1 日本
(理化学研究所 倉谷形態進化研究室)
email:菅原文昭,倉谷 滋
DOI: 10.7875/first.author.2016.015
Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain.
Fumiaki Sugahara, Juan Pascual-Anaya, Yasuhiro Oisi, Shigehiro Kuraku, Shin-ichi Aota, Noritaka Adachi, Wataru Takagi, Tamami Hirai, Noboru Sato, Yasunori Murakami, Shigeru Kuratani
Nature, 531, 97-100 (2016)
要 約
脊椎動物の脳は複雑かつ精密に領域化された器官であり,おのおのの領域が進化の過程においていつ獲得されたのかについては不明な点が多い.今回,筆者らは,現生の脊椎動物のうち最初に分岐した,顎のない円口類のうちヌタウナギの脳の発生の過程を世界ではじめて詳細に観察し,とくに,脳室の形態,神経線維の走行,遺伝子の発現パターンから脳のおのおのの領域の同定を試みた.円口類のもうひとつの系統であるヤツメウナギも同時に解析した結果,これまで円口類にはないとされてきた大脳基底核の一部,および,小脳の発生する領域が,じつは存在することが明らかにされた.これにより,脊椎動物の基本的な領域をつくるしくみは,円口類が分岐してから軟骨魚類の分岐までに獲得されたとするこれまでの考えとは異なり,円口類の分岐よりまえ,すなわち,脊椎動物がはじめて誕生した5億年以上まえに成立したことがわかった.
はじめに
脊椎動物の誕生は古生代カンブリア紀ごろと考えられている.そのころの脊椎動物の脳がどのようなものであったのかを想像するのはむずかしい.脳は軟組織であるため化石として残りにくく,絶滅した動物の化石からわかることはかぎられるためである.一方,現生の脊椎動物でもっとも古くに分岐したのは顎のない“円口類”で,ヌタウナギおよびヤツメウナギの2つの群のみが現存する.この円口類は5億年以上まえに顎のある脊椎動物“顎口類”とたもとを分かった系統なので,円口類と顎口類とを比較しそれらに共通の要素があれば,それは脊椎動物の共通祖先がもっていた形質であるとみなせる.このような方法により発生期の脳を調べ,脳をつくる遺伝子の発現の様式を円口類と顎口類とで比較すれば,脳の祖先的な状態およびその変遷をある程度まで推定することができる.
これまで,円口類の発生の研究にはおもにヤツメウナギが使用されてきた.これは,繁殖期のヤツメウナギが比較的容易に入手でき,受精卵を得るのも可能だからである.先行研究において,ヤツメウナギと顎口類の脳の発生における最大の違いは,1)ヤツメウナギの胚は大脳(終脳)のもっとも腹側の領域である内側基底核隆起をもたない,2)ヤツメウナギの胚は小脳の発生の場となる菱脳唇をもたない,の2つの点であるといわれていた.
1)について,顎口類の内側側基底核隆起は大脳基底核の一部である淡蒼球に分化し運動の制御にかかわる.さらに,内側基底核隆起からGABA作動性抑制ニューロンが生じ大脳皮質の介在ニューロンになる.つまり,内側基底核隆起は特定の領域あるいはニューロンの発生のための原基としてはたらくことから,大脳の皮質および基底核が機能するうえで欠かせない存在なのである.ところが,これまでの研究において,ヤツメウナギの胚には内側基底核隆起を特異化するNkx2.1遺伝子およびShh遺伝子の相同遺伝子の発現がみられず,また,成体においても淡蒼球および皮質においてGABA作動性の介在ニューロンはみつかっていなかった1).Nkx2.1ノックアウトマウスは淡蒼球を欠き,皮質のGABA作動性の介在ニューロンもいちじるしく減少していることから,Nkx2.1ノックアウトマウスはヤツメウナギの“表現型模写”あるいは“先祖返り”の状態を示すと理解されていた.
2)について,小脳は体性感覚や平衡感覚を受容し運動を制御する部分であるが,円口類においてはこの部分が未発達である.つまり,ヌタウナギには小脳がなく,ヤツメウナギにおいては未発達な小脳と一部の交連繊維がみられるにとどまる.顎口類の小脳は,後脳(その形態から,“菱脳”ともよばれる)の背側部である“菱脳唇”の前方から生じ,この領域はPax6遺伝子やWnt1遺伝子などの発現により特徴づけられる.ところが,ヤツメウナギの胚には後脳の背側においてPax6遺伝子の発現がみられないといわれてきた2).Pax6遺伝子は小脳の顆粒細胞層の分化および移動に必須の遺伝子でもあるので,ヤツメウナギは小脳が発生する場としての菱脳唇が進化的に未分化な状態を保持していると解釈されてきた.
以上の知見から,内側基底核隆起および菱脳唇という2つの発生における原基は,円口類と顎口類が分岐したのち,顎口類の進化の過程において新たに獲得され発達したと考えられてきたのである.
しかし,近年,この考えに対しいくつかの疑問や反論が提示された.ひとつは,ヤツメウナギの進化の過程において内側基底核隆起および菱脳唇は2次的に退化したのではないか,という疑問である3).当然ながら,ヤツメウナギは“原始的”な動物ではなく,われわれと同じ5億年の進化をへてきているので,この過程において脳の2次的な改変や退化は起こりうる.もうひとつは,ヤツメウナギの成体において淡蒼球に類似した領域,および,皮質に抗GABA抗体に陽性を示す細胞が観察されたという報告である4,5).これらは顎口類においては内側基底核隆起から分化するため,ヤツメウナギの胚にも未発見の内側基底核隆起が存在すると考えるのが自然であろうが,ヤツメウナギにおいては顎口類とは異なる発生の過程をへてこれらの領域および細胞が生じているという可能性も否定できない.
これらの疑問のうち,前者については,円口類のもうひとつの系統であるヌタウナギを解析する必要が指摘されてきた.しかし,近年まで,ヌタウナギの発生の過程は謎につつまれていた.というのも,ヌタウナギはほとんどの種が深海に生息し,受精卵の入手がきわめて困難だからである.しかし,筆者らのグループは,2007年に世界ではじめて日本産ヌタウナギ(Eptatretus burgeri,図1a)の受精卵を人工的に得ることに成功し,以来,毎年,少数ではあるが胚が得られ,いまでは多くの発生学的な知見が得られる状況になっている6).また,後者については,ヤツメウナギの胚における内側基底核隆起の有無の再検証が必要と考えた.
1.ヌタウナギの脳の発生の形態学的な記載
2011年に島根県沖で捕獲したヌタウナギから得た受精卵のうち,おもに2つを使用し解析した.まず,ヌタウナギの脳の発生の形態的な観察および記載を行った.ヌタウナギの脳の発生過程の記載については,19世紀末および20世紀初頭にかけて偶発的に得られた胚により断片的な報告があるが7,8),これらはみな,硬い卵殻がついたままの卵をエタノール固定したせいで胚がいちじるしく変形しており,正しい形態やおのおのの部位の同定が困難であった.そこで,正しく固定した胚のパラフィン切片から立体再構築像を作成し,脳の形態的な発生を観察し記載した.この結果,ほかの脊椎動物とはきわめて異なった外形を示し,内部も脳室がほぼ消失するなど非常に特異な形態を示すヌタウナギの脳(図1b)も,その発生の過程においては脊椎動物としての基本的な形態を踏襲していることがわかった.
2.遺伝子の発現および神経線維の走行による脳のおのおのの領域の同定
ヌタウナギより脳のおのおのの領域に特徴的に発現するFoxG1遺伝子,Emx1/2遺伝子,Pax6遺伝子,Nkx2.1遺伝子,Shh遺伝子の相同遺伝子をクローニングし,その発現のパターンを観察した.また,神経染色により初期の神経回路および交連繊維を同定し,その位置関係などを手がかりとして脳のおのおのの領域の同定を試みた.この結果,大脳(外套,外套下部),間脳(前視床,視床,視蓋前域,視床下部など),中脳,後脳といった顎口類のもつ基本的な脳の領域が,ヌタウナギにも存在することがわかった(図2a).ただし,松果体は成体のヌタウナギの脳にみられないが,発生の過程においてもこれを見い出すことはできなかった.だからといって,これが原始的な特徴を示しているわけではない.なぜならば,ヤツメウナギは発達した松果体をもつからである.つまり,松果体はヌタウナギの系統において2次的に失われた可能性が高い.
3.内側基底核隆起および菱脳唇における遺伝子の発現
ヌタウナギの胚において内側基底核隆起および菱脳唇の有無について調べた.転写因子Nkx2.1は顎口類においてShhと協調して内側基底核隆起を特異化する.この両者の相同遺伝子の発現を調べた結果,ともにヌタウナギの胚の終脳腹側において発現が観察された.さらに,顎口類において菱脳唇に発現するPax6遺伝子およびAtoh1遺伝子の発現もヌタウナギの後脳背側においてみられた.すなわち,ヌタウナギの脳には,顎口類と同様に,内側基底核隆起および菱脳唇が存在することがわかった(図2a).これは,ヤツメウナギから得られた知見とは大きく異なっていた.
4.ヤツメウナギの胚の再解析
これらの結果から,つぎの2つの進化のシナリオが考えられた.1)ヌタウナギおよび顎口類においてみられた内側基底核隆起および菱脳唇は共通祖先の段階で獲得されたが,ヤツメウナギの系統においてこの2つの領域は2次的に退化した.2)共通祖先の段階において内側基底核隆起および菱脳唇は獲得されておらず,ヌタウナギおよび顎口類においてそれぞれ独立に獲得された,つまり,収斂進化が起こった.これに対し,ヤツメウナギは祖先的な状態を保持している.
これらのシナリオのどちらであるかを検証するため,日本産カワヤツメ(Lethenteron japonicum)の胚を用いて再検証した.近年,公開されたカワヤツメのゲノム塩基配列を使用し9),既知であったNkx2.1遺伝子の相同遺伝子であるNkx2.1/2.4A遺伝子にくわえ,パラログ遺伝子としてNkx2.1/2.4B遺伝子およびNkx2.1/2.4C遺伝子を新規にクローニングした.パラログ遺伝子とは,遺伝子の重複により生じた複数の遺伝子のことである.これらの遺伝子の発現を発生後期のヤツメウナギの胚において観察したところ,既知のNkx2.1/2.4A遺伝子とは異なり,新規のNkx2.1/2.4B遺伝子およびNkx2.1/2.4C遺伝子の発現が大脳腹側において観察された.さらに,菱脳唇についても,既知であったPax6相同遺伝子にくわえPax6B遺伝子を新規にクローニングしたところ,Pax6B遺伝子も後脳背側において発現がみられた.つまり,ヤツメウナギにはないとされていた内側基底核隆起および菱脳唇はいずれも存在しているらしかった(図2a).先行研究においては,ゲノム情報が不足していたことから円口類に何個のパラログ遺伝子があるのか不明だったこともあり,発現していないパラログ遺伝子のみを観察してしまっていたものと考えられた.
ヤツメウナギの後脳においては,Pax6遺伝子にくわえ,Atoh1遺伝子およびPtf1a遺伝子の発現も確認された.Atoh1遺伝子およびPtf1a遺伝子は,それぞれ,顎口類の小脳の層を構成する顆粒細胞およびプルキンエ細胞の分化にきわめて重要である.それらの遺伝子が,形態的に小脳をもたないヤツメウナギの後脳において機能していたことはきわめて興味深い.すなわち,小脳を獲得するまえの段階においてすでにAtoh1遺伝子およびPtf1a遺伝子が菱脳背側の細胞において機能しており,それを下地として顎口類の系統において小脳が獲得されたとみるべきらしい.小脳の前駆体はすべての脊椎動物を産み出した共通祖先に存在していたのだ.
5.脊椎動物における脳の進化のシナリオ
これまで,内側基底核隆起および菱脳唇という2つの脳の領域は,円口類が分岐してから軟骨魚類が分岐する4億5000万年まえのあいだに獲得されたと考えられていた.しかし,今回の研究により,内側基底核隆起および菱脳唇の獲得は円口類と顎口類の共通祖先が存在したころ,すなわち,5億年以上まえにまでさかのぼることになった(図2b).脊椎動物の脳の領域は進化の過程において段階的に獲得されてきたのではなく,脊椎動物の共通祖先の段階ですでに成立していたという新たな進化のシナリオが提示された(図2b).ヒトにも用いられている脳の基本的な発生の構造は,これまで考えられてきたより古い起源をもつらしい.
おわりに
1960年代より“脳の三位一体説”が提唱され,ヒトの脳は爬虫類脳(大脳基底核),哺乳類原脳(辺縁系),哺乳類脳(新皮質)に分けることができ,これらは進化の過程で段階的に獲得された,と唱えられた10).この説はいまでも一般の脳進化の認識の根底に流れている.しかし,近年の進化発生学の研究による知見および今回の研究により,この理解は変わりつつある.すなわち,脳はまずおおまかな区画をつくり,その区画における複雑化および特異化がそれぞれの動物の系統においてなされたのである.都市の成立の過程にたとえると,市街地が中心部から郊外へと拡大していくのではなく,平城京のようにさきに土地を区分し,そのあとおのおのの区域がにぎわってくる,というイメージだろうか.
この研究において,筆者らは,脊椎動物の初期に脳のおおまかな区画や枠組みの大部分が成立したと論じた.しかしもちろん,脳はヒトにいたるまでにさらなる複雑化をとげている.では,脳のいったいどこがもっともいちじるしく進化したのだろうか? その重要なもののひとつは大脳新皮質の獲得だろう.大脳新皮質は哺乳類においてとくに発達したが,近年,爬虫類や鳥類においても類似した領域がみつかっている.さらには,最近,ヤツメウナギの大脳皮質にも哺乳類の新皮質にある運動野に似た場所のあるらしいことがわかってきた.この大脳新皮質の起源はどこまでさかのぼることができ,そして,どのような進化的な変遷をへてわれわれの“知性”が生まれたのか,非常に魅力的な研究テーマである.また,小脳はすべての顎口類にみられ,体性感覚や平衡感覚を受容し運動を制御しているが,ヒトを含む哺乳類では小脳は大脳新皮質と共同して高度な知覚機能を担うことがわかっている.このような制御中枢としての小脳はどのような発生プログラムの変遷をへて獲得されたのだろうか? このことについては,この研究において小脳の獲得の“前夜”の状態であるとわかった円口類と,小脳をもつ動物のなかでもっとも古くに分岐したサメなどの軟骨魚類との比較が,より詳細な進化の経緯を教えてくれるだろう.
文 献
- Murakami, Y., Uchida, K., Rijli, F. M. et al.: Evolution of the brain developmental plan: insights from agnathans. Dev. Biol., 280, 249-259 (2005)[PubMed]
- Murakami, Y., Ogasawara, M., Sugahara, F. et al.: Identification and expression of the lamprey Pax6 gene: evolutionary origin of the segmented brain of vertebrates. Development, 128, 3521-3531 (2001)[PubMed]
- Sugahara, F., Murakami, Y., Adachi, N. et al.: Evolution of the regionalization and patterning of the vertebrate telencephalon: what can we learn from cyclostomes? Curr. Opin. Genet. Dev., 23, 475-483 (2013)[PubMed]
- Stephenson-Jones, M., Samuelsson, E., Ericsson, J. et al.: Evolutionary conservation of the basal ganglia as a common vertebrate mechanism for action selection. Curr. Biol., 21, 1081-1091 (2011)[PubMed]
- Pombal, M. A., Alvarez-Otero, R., Perez-Fernandez, J. et al.: Development and organization of the lamprey telencephalon with special reference to the GABAergic system. Front. Neuroanat., 5, 20 (2011)[PubMed]
- Ota, K. G., Kuraku, S. & Kuratani, S.: Hagfish embryology with reference to the evolution of the neural crest. Nature, 446, 672-675 (2007)[PubMed]
- von Kupffer, C.: Studien zur vergleichenden Entwicklungsgeschichte des Kopfes der Kranioten, Heft 4: Zur Kopfentwicklung von Bdellostoma. Lehmann, Munich (1900)
- Conel, J. L.: The development of the brain of Bdellostoma stouti I. External growth changes. J. Comp. Neurol., 47, 343-403 (1929)
- Mehta, T. K., Ravi, V., Yamasaki, S. et al.: Evidence for at least six Hox clusters in the Japanese lamprey (Lethenteron japonicum). Proc. Natl .Acad. Sci. USA, 110, 16044-16049 (2013)[PubMed]
- MacLean, P. D.: The Triune Brain in Evolution: Role in Paleocerebral Functions. Springer-Verlag, New York (1990)
著者プロフィール
略歴:2011年 神戸大学大学院理学研究科 修了,理化学研究所発生・再生科学総合研究センター 研究員を経て,2013年より兵庫医科大学 講師.
研究テーマ:脊椎動物の頭部の進化発生学.
倉谷 滋(Shigeru Kuratani)
理化学研究所 主任研究員.
研究室URL:http://www.cdb.riken.jp/emo/japanese/indexj.html
© 2016 菅原文昭・倉谷 滋 Licensed under CC 表示 2.1 日本
