胚のなかで中胚葉領域と内胚葉領域はどのように決まるのか
高鳥 直士
(首都大学東京大学院理工学研究科 生命科学専攻発生生物学研究室)
email:高鳥直士
DOI: 10.7875/first.author.2015.143
Polarization of PI3K activity initiated by ooplasmic segregation guides nuclear migration in the mesendoderm.
Naohito Takatori, Kouhei Oonuma, Hiroki Nishida, Hidetoshi Saiga
Developmental Cell, 35, 333-343 (2015)
胚葉の運命分離は動物胚のなかで多様な細胞がつくられる過程の基礎である.脊索動物であるホヤでは中胚葉と内胚葉の運命分離の過程において,中内胚葉細胞の核が将来の中胚葉側に移動して転写因子NotをコードするmRNAを中胚葉側の細胞質に局在させることにより運命分離を起こすことがわかっていた.しかし,中内胚葉細胞の核が移動する向き,いい換えれば,将来の中胚葉領域がどのように決まるのかはわかっていなかった.今回,核の移動方向を決める機構を解析した結果,中内胚葉細胞におけるPI3キナーゼの局在に依存したホスファチジルイノシトール3,4,5-トリスリン酸の局在が核の移動方向を決定することが示唆された.また,受精の直後の卵細胞質の再配置により母性PI3キナーゼが局在し,そののち,母性タンパク質の局所的な活性に依存して胚性PI3キナーゼが局在することにより,中内胚葉細胞におけるPI3キナーゼの局在が形成されることが示唆された.この研究において,中胚葉と内胚葉の運命分離にいたる因果関係の連鎖の枠組みが受精の直後にまで遡及された.
われわれの体を構成する多様な細胞がどのようにしてたったひとつの卵細胞からつくられるのかを明らかにすることは発生生物学の主要な命題のひとつである.多様な細胞が形成される過程の初期に外胚葉,中胚葉,内胚葉の3つの胚葉がつくられ,それぞれから神経や表皮,筋肉や血管,消化管や肺といった器官や組織がつくられる.細胞の運命決定の過程を受精から順に理解するためには胚葉を形成する機構を明らかにすることが必要だが,細胞ひとつひとつのレベルで分子機構にまでふみ込んで胚葉の形成機構が解明されている例は多くはない1).線虫においては,中内胚葉細胞と隣接する細胞からのWntシグナルによって中内胚葉細胞に極性が形成されることにより,娘細胞のあいだで中胚葉と内胚葉の運命分離が起こる2-4).脊索動物であるホヤにおいては,16細胞期の中内胚葉細胞が分裂し32細胞期において中胚葉細胞と内胚葉細胞が形成される.
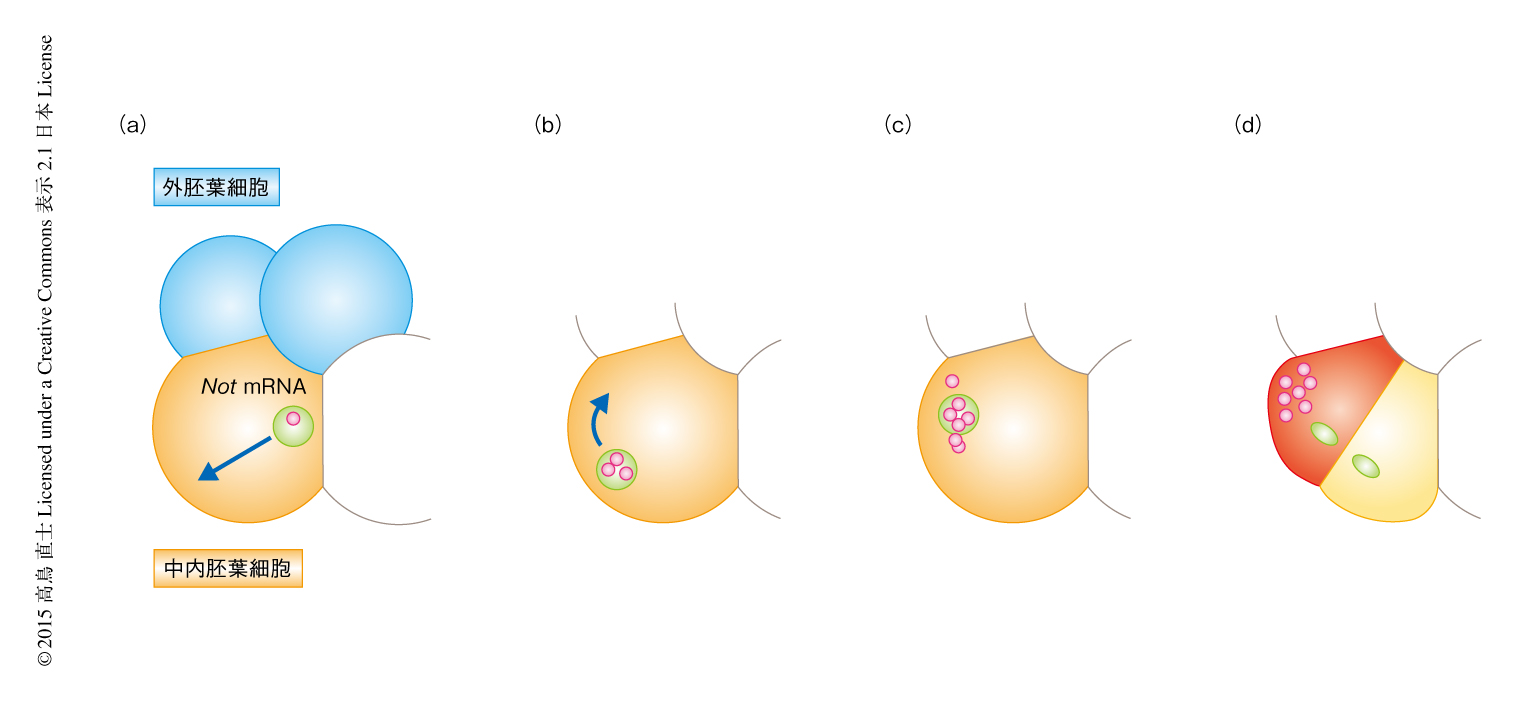
筆者らは,以前の研究において,Notとよばれるホメオドメイン転写因子がこの運命分離に重要であることを明らかにした5).Notの発現を抑制した胚においては予定中胚葉細胞がすべて内胚葉細胞になった.NotをコードするmRNAの発現パターンはすでに発表されていたが6),中内胚葉細胞および娘細胞における発現パターンを詳細に調べたところ,NotをコードするmRNAは中内胚葉細胞において転写されたのち,中胚葉細胞へと非対称に受け継がれ,中胚葉細胞では転写されないことが示唆された5).中胚葉細胞に非対称に分配されたNotをコードするmRNAからの翻訳を細胞に特異的なモルフォリーノオリゴヌクレオチドの顕微注入により阻害すると,予定中胚葉細胞が内胚葉へと分化した.反対に,NotをコードするmRNAの顕微注入により異所的にNotを発現させた内胚葉細胞は,低い確率ではあったが中胚葉へと分化した.これらの結果から,NotをコードするmRNAの中胚葉細胞への非対称な分配が中胚葉と内胚葉の運命分離に重要であることがわかってきた.また,NotをコードするmRNAの分配の過程において,中内胚葉細胞の核が将来の中胚葉細胞を生じる側に移動すること,移動の過程ではNotをコードするmRNAのほとんどが核のなかに検出されることがわかった5).これらの結果から,中内胚葉細胞の核が移動することによりNotをコードするmRNAを将来の中胚葉側へと移動させ,局在したNotをコードするmRNAが一方の娘細胞に受け継がれることにより運命分離が起こる,というモデルが提唱された(図1).この過程において鍵になるのが,中内胚葉細胞の核の移動である.ホヤの胚の卵割パターンは原腸陥入まで個体のあいだで差がなく,必ず決まった位置に決まった個数の中胚葉細胞が形成される.したがって,中内胚葉細胞の核の移動方向はなんらかの機構の制御のもとにあると想像されたが,その機構はまったく不明であった.
核の移動方向を決める因子を探るため,核に局在する蛍光タンパク質を発現させた胚を蛍光顕微鏡のもとで観察しつつさまざまな阻害剤で処理することにより,核の移動方向を異常にする阻害剤を探索した.その結果,PI3キナーゼαサブユニットに対する特異的な阻害剤で処理した胚は,核が将来の中胚葉側ではなく細胞の中央へと移動した.PI3キナーゼの発現を抑制した胚,あるいは,PI3キナーゼを過剰に発現した胚においても,核の移動方向の異常が観察された.免疫染色法により,PI3キナーゼは中内胚葉細胞の中胚葉側の細胞膜に局在することが示唆された.PI3キナーゼは細胞膜に存在するホスファチジルイノシトール4,5-ビスリン酸をリン酸化してホスファチジルイノシトール3,4,5-トリスリン酸を産生する.ホスファチジルイノシトール3,4,5-トリスリン酸はAkt/PKBなどを細胞膜にリクルートすることによりPI3キナーゼシグナルを伝達するセカンドメッセンジャーとしてはたらく.中内胚葉細胞においては,ホスファチジルイノシトール3,4,5-トリスリン酸が中胚葉側の細胞膜にかたよって検出された.PI3キナーゼの基質であるホスファチジルイノシトール4,5-ビスリン酸,および,ホスファチジルイノシトール3,4,5-トリスリン酸を脱リン酸化してホスファチジルイノシトール4,5-ビスリン酸を産生するPTENの局在を調べたが,中胚葉側あるいは内胚葉側の細胞膜のどちらかにかたよっているという結果は得られなかった.また,PI3キナーゼの発現を抑制あるいは異所的に発現させた胚においては,ホスファチジルイノシトール3,4,5-トリスリン酸の中胚葉側へのかたよりが乱された.これらの結果から,PI3キナーゼのかたよりに依存してホスファチジルイノシトール3,4,5-トリスリン酸が中胚葉側にかたよることがわかった.
PI3キナーゼとホスファチジルイノシトール3,4,5-トリスリン酸はともに中胚葉側にかたよっていたことから,どちらが核の移動方向を決めるのに重要かを区別するため,中内胚葉細胞にホスファチジルイノシトール3,4,5-トリスリン酸を顕微注入した.その結果,核は中胚葉側ではなく細胞の中央へと移動した.また,PTENの発現を抑制した胚においてはホスファチジルイノシトール3,4,5-トリスリン酸のかたよりおよび核の移動方向が異常になったが,PI3キナーゼの局在は正常であった.これらの結果から,核の移動方向はPI3キナーゼの局在に依存したホスファチジルイノシトール3,4,5-トリスリン酸のかたよりにより決定されることが示唆された.
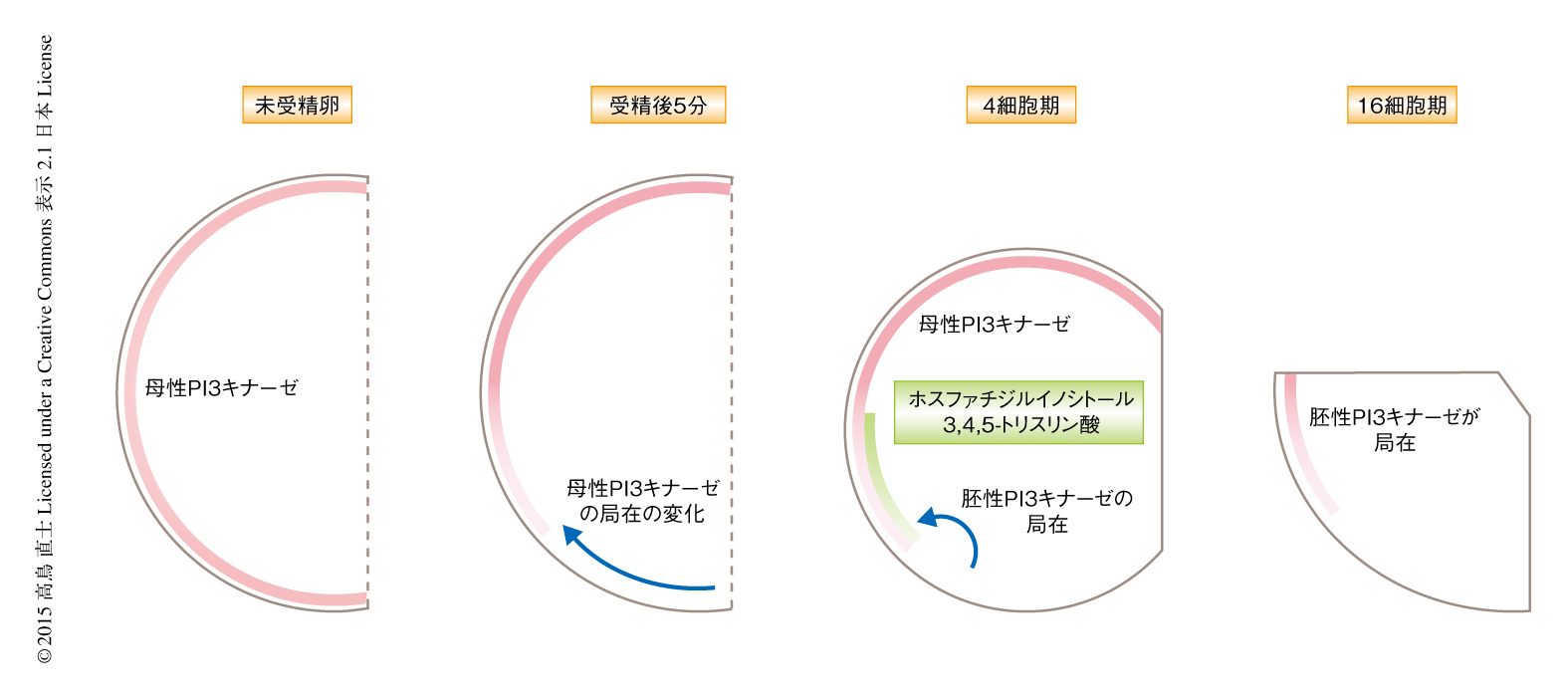
PI3キナーゼは中内胚葉細胞において中胚葉側にかたよっていた.このかたよりがいつ形成されたのかを調べるため,未受精卵から核の移動にいたるまでの発生の過程におけるPI3キナーゼの局在を免疫染色法により調べた.未受精卵において母性PI3キナーゼは将来の内胚葉領域である植物極を中心にゆるやかな濃度勾配で細胞膜の近傍に存在していた.受精ののちPI3キナーゼの分布は変化し,動物極を中心とした濃度勾配が観察され,PI3キナーゼは将来の中胚葉側に多く内胚葉側に少ないというかたよりが現われた.このかたよりは2細胞期以降も一貫して観察され,中内胚葉細胞において核が移動する16細胞期まで観察された.受精ののち一貫して中胚葉側にPI3キナーゼが多いのだとすると,ホスファチジルイノシトール3,4,5-トリスリン酸が産生され中胚葉側に核を移動させる極性が形成されるのは16細胞期だけとかぎらなくなる.おのおののステージにおいてホスファチジルイノシトール3,4,5-トリスリン酸の局在を免疫染色法により観察したところ,4細胞期および16細胞期において将来の中胚葉領域に限局していた.阻害剤による一過的な処理により,4細胞期あるいは16細胞期のどちらにおいてもホスファチジルイノシトール3,4,5-トリスリン酸は16細胞期における核の移動方向の決定に重要であること,4細胞期のホスファチジルイノシトール3,4,5-トリスリン酸は16細胞期においてPI3キナーゼを将来の中胚葉側に局在させるのに重要であることが示唆された.PI3キナーゼの発現を抑制した胚の観察から,4細胞期のPI3キナーゼは母性タンパク質,16細胞期のPI3キナーゼは胚性タンパク質であることが示唆された.これらの結果から,母性PI3キナーゼが将来の中胚葉領域において活性化しホスファチジルイノシトール3,4,5-トリスリン酸が局所的に産生された結果,胚性PI3キナーゼが中胚葉領域に局在すると考えられた.
母性PI3キナーゼの局在はどのように形成されるのだろうか? ホヤにおいては,受精ののちの2段階の卵細胞質の再配置により胚における極性が顕在化する7).PI3キナーゼの局在が変化する時期は,第1段階である受精の直後の植物極側への卵表層の細胞質の移動の時期に一致していた.受精の直後の卵細胞質の再配置はアクチン線維に依存して進行する8).そこで,PI3キナーゼの局在がアクチン線維に依存して変化するかどうか調べたところ,アクチン重合阻害剤により処理した胚においてはPI3キナーゼの局在の変化は観察されなかった.受精ののちのアクチン線維の植物極への移動とPI3キナーゼの局在の変化の進行は分単位で一致していた.これらの結果から,受精の直後の卵細胞質の再配置にともない,PI3キナーゼの局在が変化するというかたちで中胚葉と内胚葉の違いが顕在化するという仮説がたてられた.
中胚葉と内胚葉の運命分離において重要な役割をはたす核の移動方向を決定するタンパク質を見い出し,その局在を形成し維持する機構の枠組みが受精の直後にまでさかのぼり明らかにされた.母性PI3キナーゼの局在の変化により形成された中内胚葉細胞における極性がPI3キナーゼの活性により維持され,中内胚葉細胞において核の移動方向を規定し,核の移動により転写因子NotをコードするmRNAが非対称に分配されることで運命分離が起こる,という中胚葉と内胚葉の運命分離の機構が考えられた(図2).中胚葉と内胚葉の運命分離にいたる過程にはまだ不明な点が多い.受精の直後のアクチン線維の凝集方向と母性PI3キナーゼの移動方向は逆であり,アクチン線維にそった輸送や局所的な分解などさまざまなモデルを考えることはできるが,PI3キナーゼの局在変化の機構はいまだ不明である.4細胞期に中胚葉領域に限局して観察されるホスファチジルイノシトール3,4,5-トリスリン酸がどのようにして胚性PI3キナーゼを局在させるのか,16細胞期においてホスファチジルイノシトール3,4,5-トリスリン酸の局在にしたがい核を移動させる機構など,ほかにも不明な点はある.中胚葉と内胚葉の運命分離の機構の包括的な理解には,今後,さらなる解析が必要である.核の移動に依存した極性の顕在化は今後も多くの細胞においてみつかるのではないだろうか.この研究が,そうした研究の一助になれば幸いである.
略歴:2005年 京都大学大学院理学研究科博士課程 修了,2012年 首都大学東京大学院理工学研究科 助教を経て,2015年より同 准教授.
研究テーマ:細胞の極性を形成する機構.核の移動方向を決める分子機構.中胚葉と内胚葉の運命分離の機構.
関心事:生命研究の方法論史.甘いもの.
© 2015 高鳥 直士 Licensed under CC 表示 2.1 日本
(首都大学東京大学院理工学研究科 生命科学専攻発生生物学研究室)
email:高鳥直士
DOI: 10.7875/first.author.2015.143
Polarization of PI3K activity initiated by ooplasmic segregation guides nuclear migration in the mesendoderm.
Naohito Takatori, Kouhei Oonuma, Hiroki Nishida, Hidetoshi Saiga
Developmental Cell, 35, 333-343 (2015)
要 約
胚葉の運命分離は動物胚のなかで多様な細胞がつくられる過程の基礎である.脊索動物であるホヤでは中胚葉と内胚葉の運命分離の過程において,中内胚葉細胞の核が将来の中胚葉側に移動して転写因子NotをコードするmRNAを中胚葉側の細胞質に局在させることにより運命分離を起こすことがわかっていた.しかし,中内胚葉細胞の核が移動する向き,いい換えれば,将来の中胚葉領域がどのように決まるのかはわかっていなかった.今回,核の移動方向を決める機構を解析した結果,中内胚葉細胞におけるPI3キナーゼの局在に依存したホスファチジルイノシトール3,4,5-トリスリン酸の局在が核の移動方向を決定することが示唆された.また,受精の直後の卵細胞質の再配置により母性PI3キナーゼが局在し,そののち,母性タンパク質の局所的な活性に依存して胚性PI3キナーゼが局在することにより,中内胚葉細胞におけるPI3キナーゼの局在が形成されることが示唆された.この研究において,中胚葉と内胚葉の運命分離にいたる因果関係の連鎖の枠組みが受精の直後にまで遡及された.
はじめに
われわれの体を構成する多様な細胞がどのようにしてたったひとつの卵細胞からつくられるのかを明らかにすることは発生生物学の主要な命題のひとつである.多様な細胞が形成される過程の初期に外胚葉,中胚葉,内胚葉の3つの胚葉がつくられ,それぞれから神経や表皮,筋肉や血管,消化管や肺といった器官や組織がつくられる.細胞の運命決定の過程を受精から順に理解するためには胚葉を形成する機構を明らかにすることが必要だが,細胞ひとつひとつのレベルで分子機構にまでふみ込んで胚葉の形成機構が解明されている例は多くはない1).線虫においては,中内胚葉細胞と隣接する細胞からのWntシグナルによって中内胚葉細胞に極性が形成されることにより,娘細胞のあいだで中胚葉と内胚葉の運命分離が起こる2-4).脊索動物であるホヤにおいては,16細胞期の中内胚葉細胞が分裂し32細胞期において中胚葉細胞と内胚葉細胞が形成される.
筆者らは,以前の研究において,Notとよばれるホメオドメイン転写因子がこの運命分離に重要であることを明らかにした5).Notの発現を抑制した胚においては予定中胚葉細胞がすべて内胚葉細胞になった.NotをコードするmRNAの発現パターンはすでに発表されていたが6),中内胚葉細胞および娘細胞における発現パターンを詳細に調べたところ,NotをコードするmRNAは中内胚葉細胞において転写されたのち,中胚葉細胞へと非対称に受け継がれ,中胚葉細胞では転写されないことが示唆された5).中胚葉細胞に非対称に分配されたNotをコードするmRNAからの翻訳を細胞に特異的なモルフォリーノオリゴヌクレオチドの顕微注入により阻害すると,予定中胚葉細胞が内胚葉へと分化した.反対に,NotをコードするmRNAの顕微注入により異所的にNotを発現させた内胚葉細胞は,低い確率ではあったが中胚葉へと分化した.これらの結果から,NotをコードするmRNAの中胚葉細胞への非対称な分配が中胚葉と内胚葉の運命分離に重要であることがわかってきた.また,NotをコードするmRNAの分配の過程において,中内胚葉細胞の核が将来の中胚葉細胞を生じる側に移動すること,移動の過程ではNotをコードするmRNAのほとんどが核のなかに検出されることがわかった5).これらの結果から,中内胚葉細胞の核が移動することによりNotをコードするmRNAを将来の中胚葉側へと移動させ,局在したNotをコードするmRNAが一方の娘細胞に受け継がれることにより運命分離が起こる,というモデルが提唱された(図1).この過程において鍵になるのが,中内胚葉細胞の核の移動である.ホヤの胚の卵割パターンは原腸陥入まで個体のあいだで差がなく,必ず決まった位置に決まった個数の中胚葉細胞が形成される.したがって,中内胚葉細胞の核の移動方向はなんらかの機構の制御のもとにあると想像されたが,その機構はまったく不明であった.
1.中内胚葉細胞の核の移動方向は局所的なPI3キナーゼシグナルにより決定される
核の移動方向を決める因子を探るため,核に局在する蛍光タンパク質を発現させた胚を蛍光顕微鏡のもとで観察しつつさまざまな阻害剤で処理することにより,核の移動方向を異常にする阻害剤を探索した.その結果,PI3キナーゼαサブユニットに対する特異的な阻害剤で処理した胚は,核が将来の中胚葉側ではなく細胞の中央へと移動した.PI3キナーゼの発現を抑制した胚,あるいは,PI3キナーゼを過剰に発現した胚においても,核の移動方向の異常が観察された.免疫染色法により,PI3キナーゼは中内胚葉細胞の中胚葉側の細胞膜に局在することが示唆された.PI3キナーゼは細胞膜に存在するホスファチジルイノシトール4,5-ビスリン酸をリン酸化してホスファチジルイノシトール3,4,5-トリスリン酸を産生する.ホスファチジルイノシトール3,4,5-トリスリン酸はAkt/PKBなどを細胞膜にリクルートすることによりPI3キナーゼシグナルを伝達するセカンドメッセンジャーとしてはたらく.中内胚葉細胞においては,ホスファチジルイノシトール3,4,5-トリスリン酸が中胚葉側の細胞膜にかたよって検出された.PI3キナーゼの基質であるホスファチジルイノシトール4,5-ビスリン酸,および,ホスファチジルイノシトール3,4,5-トリスリン酸を脱リン酸化してホスファチジルイノシトール4,5-ビスリン酸を産生するPTENの局在を調べたが,中胚葉側あるいは内胚葉側の細胞膜のどちらかにかたよっているという結果は得られなかった.また,PI3キナーゼの発現を抑制あるいは異所的に発現させた胚においては,ホスファチジルイノシトール3,4,5-トリスリン酸の中胚葉側へのかたよりが乱された.これらの結果から,PI3キナーゼのかたよりに依存してホスファチジルイノシトール3,4,5-トリスリン酸が中胚葉側にかたよることがわかった.
PI3キナーゼとホスファチジルイノシトール3,4,5-トリスリン酸はともに中胚葉側にかたよっていたことから,どちらが核の移動方向を決めるのに重要かを区別するため,中内胚葉細胞にホスファチジルイノシトール3,4,5-トリスリン酸を顕微注入した.その結果,核は中胚葉側ではなく細胞の中央へと移動した.また,PTENの発現を抑制した胚においてはホスファチジルイノシトール3,4,5-トリスリン酸のかたよりおよび核の移動方向が異常になったが,PI3キナーゼの局在は正常であった.これらの結果から,核の移動方向はPI3キナーゼの局在に依存したホスファチジルイノシトール3,4,5-トリスリン酸のかたよりにより決定されることが示唆された.
2.中内胚葉細胞におけるPI3キナーゼの局在は母性PI3キナーゼの活性に依存する
PI3キナーゼは中内胚葉細胞において中胚葉側にかたよっていた.このかたよりがいつ形成されたのかを調べるため,未受精卵から核の移動にいたるまでの発生の過程におけるPI3キナーゼの局在を免疫染色法により調べた.未受精卵において母性PI3キナーゼは将来の内胚葉領域である植物極を中心にゆるやかな濃度勾配で細胞膜の近傍に存在していた.受精ののちPI3キナーゼの分布は変化し,動物極を中心とした濃度勾配が観察され,PI3キナーゼは将来の中胚葉側に多く内胚葉側に少ないというかたよりが現われた.このかたよりは2細胞期以降も一貫して観察され,中内胚葉細胞において核が移動する16細胞期まで観察された.受精ののち一貫して中胚葉側にPI3キナーゼが多いのだとすると,ホスファチジルイノシトール3,4,5-トリスリン酸が産生され中胚葉側に核を移動させる極性が形成されるのは16細胞期だけとかぎらなくなる.おのおののステージにおいてホスファチジルイノシトール3,4,5-トリスリン酸の局在を免疫染色法により観察したところ,4細胞期および16細胞期において将来の中胚葉領域に限局していた.阻害剤による一過的な処理により,4細胞期あるいは16細胞期のどちらにおいてもホスファチジルイノシトール3,4,5-トリスリン酸は16細胞期における核の移動方向の決定に重要であること,4細胞期のホスファチジルイノシトール3,4,5-トリスリン酸は16細胞期においてPI3キナーゼを将来の中胚葉側に局在させるのに重要であることが示唆された.PI3キナーゼの発現を抑制した胚の観察から,4細胞期のPI3キナーゼは母性タンパク質,16細胞期のPI3キナーゼは胚性タンパク質であることが示唆された.これらの結果から,母性PI3キナーゼが将来の中胚葉領域において活性化しホスファチジルイノシトール3,4,5-トリスリン酸が局所的に産生された結果,胚性PI3キナーゼが中胚葉領域に局在すると考えられた.
3.母性PI3キナーゼは卵細胞質の再配置により移動する
母性PI3キナーゼの局在はどのように形成されるのだろうか? ホヤにおいては,受精ののちの2段階の卵細胞質の再配置により胚における極性が顕在化する7).PI3キナーゼの局在が変化する時期は,第1段階である受精の直後の植物極側への卵表層の細胞質の移動の時期に一致していた.受精の直後の卵細胞質の再配置はアクチン線維に依存して進行する8).そこで,PI3キナーゼの局在がアクチン線維に依存して変化するかどうか調べたところ,アクチン重合阻害剤により処理した胚においてはPI3キナーゼの局在の変化は観察されなかった.受精ののちのアクチン線維の植物極への移動とPI3キナーゼの局在の変化の進行は分単位で一致していた.これらの結果から,受精の直後の卵細胞質の再配置にともない,PI3キナーゼの局在が変化するというかたちで中胚葉と内胚葉の違いが顕在化するという仮説がたてられた.
おわりに
中胚葉と内胚葉の運命分離において重要な役割をはたす核の移動方向を決定するタンパク質を見い出し,その局在を形成し維持する機構の枠組みが受精の直後にまでさかのぼり明らかにされた.母性PI3キナーゼの局在の変化により形成された中内胚葉細胞における極性がPI3キナーゼの活性により維持され,中内胚葉細胞において核の移動方向を規定し,核の移動により転写因子NotをコードするmRNAが非対称に分配されることで運命分離が起こる,という中胚葉と内胚葉の運命分離の機構が考えられた(図2).中胚葉と内胚葉の運命分離にいたる過程にはまだ不明な点が多い.受精の直後のアクチン線維の凝集方向と母性PI3キナーゼの移動方向は逆であり,アクチン線維にそった輸送や局所的な分解などさまざまなモデルを考えることはできるが,PI3キナーゼの局在変化の機構はいまだ不明である.4細胞期に中胚葉領域に限局して観察されるホスファチジルイノシトール3,4,5-トリスリン酸がどのようにして胚性PI3キナーゼを局在させるのか,16細胞期においてホスファチジルイノシトール3,4,5-トリスリン酸の局在にしたがい核を移動させる機構など,ほかにも不明な点はある.中胚葉と内胚葉の運命分離の機構の包括的な理解には,今後,さらなる解析が必要である.核の移動に依存した極性の顕在化は今後も多くの細胞においてみつかるのではないだろうか.この研究が,そうした研究の一助になれば幸いである.
文 献
- Rodaway, A. & Patient, R.: Mesendoderm. an ancient germ layer? Cell, 105, 169-172 (2001)[PubMed]
- Goldstein, B.: Establishment of gut fate in the E lineage of C. elegans: the roles of lineage-dependent mechanisms and cell interactions. Development, 118, 1267-1277 (1993)[PubMed]
- Goldstein, B.: Induction of gut in Caenorhabditis elegans embryos. Nature, 357, 255-257 (1992)[PubMed]
- Goldstein, B., Takeshita, H., Mizumoto, K. et al.: Wnt signals can function as positional cues in establishing cell polarity. Dev. Cell, 10, 391-396 (2006)[PubMed]
- Takatori, N., Kumano, G., Saiga, H. et al.: Segregation of germ layer fates by nuclear migration-dependent localization of Not mRNA. Dev. Cell, 19, 589-598 (2010)[PubMed]
- Utsumi, N., Shimojima, Y. & Saiga, H.: Analysis of ascidian Not genes highlights their evolutionarily conserved and derived features of structure and expression in development. Dev. Genes Evol., 214, 460-465 (2004)[PubMed]
- Sardet, C., Paix, A., Prodon, F. et al.: From oocyte to 16-cell stage: cytoplasmic and cortical reorganizations that pattern the ascidian embryo. Dev. Dyn., 236, 1716-1731 (2007)[PubMed]
- Sasakura, Y., Ogasawara, M. & Makabe, K. W.: Two pathways of maternal RNA localization at the posterior-vegetal cytoplasm in early ascidian embryos. Dev. Biol., 220, 365-378 (2000)[PubMed]
著者プロフィール
略歴:2005年 京都大学大学院理学研究科博士課程 修了,2012年 首都大学東京大学院理工学研究科 助教を経て,2015年より同 准教授.
研究テーマ:細胞の極性を形成する機構.核の移動方向を決める分子機構.中胚葉と内胚葉の運命分離の機構.
関心事:生命研究の方法論史.甘いもの.
© 2015 高鳥 直士 Licensed under CC 表示 2.1 日本
