根寄生植物ストライガのもつストリゴラクトン受容体は高い感受性をもつ
藤 茂雄
(カナダToronto大学Department of Cell and Systems Biology)
email:藤 茂雄
DOI: 10.7875/first.author.2015.128
Structure-function analysis identifies highly sensitive strigolactone receptors in Striga.
Shigeo Toh, Duncan Holbrook-Smith, Peter J. Stogios, Olena Onopriyenko, Shelley Lumba, Yuichiro Tsuchiya, Alexei Savchenko, Peter McCourt
Science, 350, 203-207 (2015)
ストリゴラクトンは内生の植物ホルモンであり,分化,菌類との相互作用,そして,根寄生植物における宿主植物の認識において重要なはたらきを担う.筆者らは,ストライガとよばれる根寄生植物のもつストリゴラクトン受容体をコードすると考えられる11個の遺伝子について,モデル植物であるシロイヌナズナに導入することによりその生理活性を調べた.その結果,非寄生植物の1万倍以上に相当する非常に高感受性のひとつを含む,4つの重要なストリゴラクトン受容体の同定に成功した.さらに,それらのうちのひとつの結晶構造を解析した結果,ストライガのもつトリゴラクトン受容体はストリゴラクトンの結合するポケットの大きさが大きくかたちも異なっており,それによりさまざまなストリゴラクトンに応答できるよう進化した可能性が考えられた.さらに,この超高感受性のストリゴラクトン受容体をもつシロイヌナズナの種子をバイオセンサーとする,ストリゴラクトンの簡易的なバイオアッセイ系の開発にも成功した.この研究の成果は,ストライガのもつストリゴラクトン受容体についての理解を促進し,将来的には,ストライガの発芽を制御する化合物の開発や耐性をもつ品種の育種への応用が考えられ,アフリカにおける食糧問題の解決への貢献が期待される.
内生の植物ホルモンであるストリゴラクトンは,2008年,イネおよびシロイヌナズナにおいて枝分かれを制御するホルモンとしての機能が明らかにされた1).さらに,植物の根から放出されるストリゴラクトンはアーバスキュラー菌根菌との共生において重要なシグナルとしてはたらき2),じつに80%以上の植物がこれらのシグナルを窒素固定において必要としていると考えられている.そもそも,ストリゴラクトンはストライガとよばれる根寄生植物の発芽を誘導する化学物質として発見された3).多くの植物は貧栄養条件にさらされると根から多量のストリゴラクトンを放出し,それにより菌根の形成を促進すると考えられている4).つまり,ストライガはこの宿主植物からのシグナルをハイジャックして宿主植物が近くにいることを示すシグナルとして利用し,発芽していることが明らかにされた.実際,ストライガの被害は貧栄養条件にある農作地ほど大きく,ストライガはひとたびストリゴラクトンを受容すると発芽し,すぐに根を宿主植物の根に貫入して栄養を搾取する.ストライガによる被害は国連により食糧問題のなかでも最大の脅威と認定され,とくにアフリカの農業に大きな被害をもたらしている5).アフリカの耕作地のじつに2/3がすでにストライガにより汚染されており,その面積は40万km2にも及ぶ.これにより,少なくとも25カ国の1億人が被害をうけているとされている.
これまでの研究により,α/βヒドロラーゼがストリゴラクトンの受容において中心的な役割を担うことが示されている.たとえば,α/βヒドロラーゼファミリーに属するD14はイネにおいて枝分かれに重要なストリゴラクトン受容体である6,7).一方,D14と同じファミリーに属するHTLは,シロイヌナズナにおいて発芽に重要なはたらきを担うことが明らかにされた8).ストライガの発芽にはストリゴラクトンが必要なことから,HTLはストライガの発芽においても重要な役割を担うことが推測された.ストライガのRNA-seqデータベースを検索したところ,HTL遺伝子と相同性をもつ11個の遺伝子がみつかった9,10).根寄生植物であるストライガはその生育に宿主植物を必要とすることや,他殖性で種子を得るのには複数の個体が必要であることから,研究室で扱うのは非常に困難でより簡便な研究手法がもとめられていた.また,11個の遺伝子それぞれの機能を明らかにすることは,ストライガを使っていては困難であった.そこで,11個の遺伝子それぞれすべてをシロイヌナズナに別々に導入して発現させ,それぞれにコードされるタンパク質の生体における活性を調べた.その結果,9個のタンパク質が生体においてストリゴラクトン受容体としての活性をもっていた.また,筆者らにより開発されたストリゴラクトン様の活性をもつアゴニストを用いた機能解析により11),4個のタンパク質についてはアゴニストの構造に対する特異性が低下し,より多くのアゴニストに対して応答することが明らかにされた.それぞれの植物からそれぞれ異なる構造をもつストリゴラクトンが放出されることから,このような性質はストライガにとりより多くの宿主植物に対して応答するため重要であったのかもしれない.さらに,その4個のタンパク質のうちの1つShHTL7を発現したシロイヌナズナの種子は,10 pMのストリゴラクトンにも応答して発芽が誘導された.これは,耳かき1杯のストリゴラクトンを50 mプールに溶かした濃度に匹敵する.この実験による重要な発見は,ストライガにおいて超高感受性をもつストリゴラクトン受容体としてShHTL7を同定したこと,そして,ストリゴラクトン受容体の単独の進化のみによりこのような超高感受性の獲得を説明する可能性を示したことである.
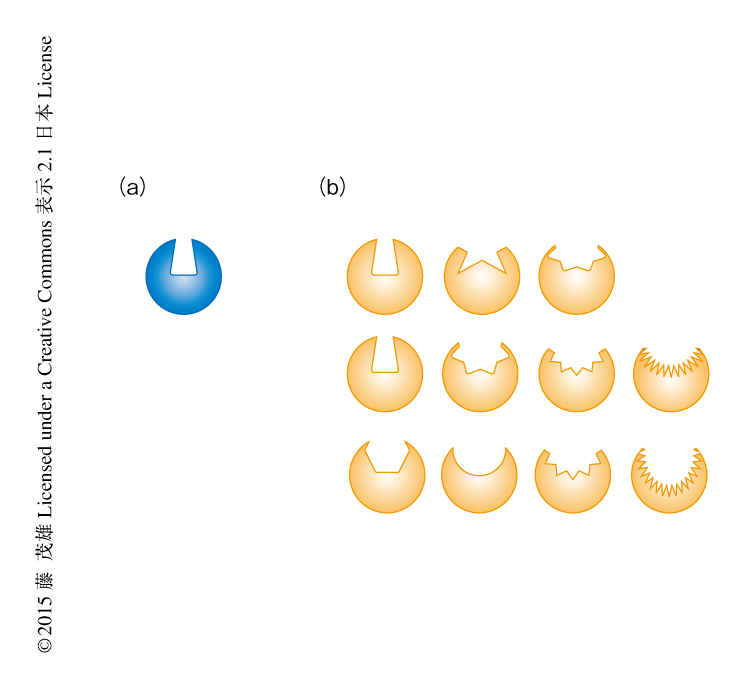
ストライガのもつストリゴラクトン受容体はどのように超高感受性を獲得したのかを明らかにするため,結晶構造解析を試みた.HTL遺伝子と相同性をもつ11個の遺伝子にコードされるタンパク質すべての結晶化を試みたところ,重要な4個のタンパク質のうちの1つShHTL5の結晶化に成功した.その結晶構造を非寄生植物のHTLと比較したところ,ストリゴラクトンの結合するポケットのサイズが約2.5倍大きく,また,その入口の大きさも大きくなっていた(図1).ShHTL5の結晶構造から得られた情報をもとに,ストリゴラクトンに対し超高感受性をもつShHTL7のアミノ酸配列をみたところ,もっとも特徴的に異なる配列をもっていた.このことから,ストライガはストリゴラクトンの結合するポケットの部分に変異を起こすことにより,ストリゴラクトンに対する感受性を高めると同時に,さまざまな構造をもつストリゴラクトンにも応答できるという,いっけん相反する機能を同時に実現したと考えられた.この実験による重要な発見は,ストリゴラクトン受容体の構造を明らかにしてストリゴラクトンの結合するポケットの大きさの増大が重要であることを示し,また,それにかかわるアミノ酸残基の候補を明らかにしたことである.
ストライガの防除にむけての重要なファクターとして,化合物による発芽の制御および耐性をもつ品種の育種があげられる.しかしながら,さきに述べたとおり,ストライガは実験には不向きであること,また,有害な植物であることから,実際に研究を行える研究機関は限られてしまう.そこで,超高感受性をもつストリゴラクトン受容体であるShHTL7を利用してシロイヌナズナの種子を使ったバイオアッセイ系を構築できないか検討した.その結果,実際にイネを生育させ,その根のまわりにShHTL7を発現させたシロイヌナズナの種子を散布したところ,イネの根からの浸出液によりシロイヌナズナの発芽が促進された.このことから,このバイオアッセイ系により,少なくともイネにおける育種が可能であることが示された(図2).

ストライガのもつストリゴラクトン受容体は非寄生植物のものと比較して,ストリゴラクトンの結合するポケットが大きくまた超高感受性へと変異していた.筆者らは,このようなストリゴラクトン受容体の変異がストライガのような根寄生植物の特殊な発芽様式および生活環を可能にしていると推測した.しかしながら,ストライガのもつストリゴラクトン受容体ShHTL7がどのようにしてストリゴラクトンに対する選択性のあいまいさと超高感受性とを同時に可能にしているのかはいまだに謎のままであり,今後のさらなる解析が必要である.また,実際にアフリカの大地に蔓延しているストライガの防除に取り組むためには,まだまだ多くの課題が残されている.しかしながら,今回の発見により研究者が標的とすべきストリゴラクトン受容体が明らかにされた.これにより,ストライガの防除にむけた研究が活発化し飛躍的に促進されることを期待したい.
略歴:2008年 明治大学大学院農学研究科 修了,同年 同 博士研究員を経て,2009年よりカナダToronto大学 博士研究員.
研究テーマ:ストリゴラクトンのシグナル伝達の機構.
© 2015 藤 茂雄 Licensed under CC 表示 2.1 日本
(カナダToronto大学Department of Cell and Systems Biology)
email:藤 茂雄
DOI: 10.7875/first.author.2015.128
Structure-function analysis identifies highly sensitive strigolactone receptors in Striga.
Shigeo Toh, Duncan Holbrook-Smith, Peter J. Stogios, Olena Onopriyenko, Shelley Lumba, Yuichiro Tsuchiya, Alexei Savchenko, Peter McCourt
Science, 350, 203-207 (2015)
要 約
ストリゴラクトンは内生の植物ホルモンであり,分化,菌類との相互作用,そして,根寄生植物における宿主植物の認識において重要なはたらきを担う.筆者らは,ストライガとよばれる根寄生植物のもつストリゴラクトン受容体をコードすると考えられる11個の遺伝子について,モデル植物であるシロイヌナズナに導入することによりその生理活性を調べた.その結果,非寄生植物の1万倍以上に相当する非常に高感受性のひとつを含む,4つの重要なストリゴラクトン受容体の同定に成功した.さらに,それらのうちのひとつの結晶構造を解析した結果,ストライガのもつトリゴラクトン受容体はストリゴラクトンの結合するポケットの大きさが大きくかたちも異なっており,それによりさまざまなストリゴラクトンに応答できるよう進化した可能性が考えられた.さらに,この超高感受性のストリゴラクトン受容体をもつシロイヌナズナの種子をバイオセンサーとする,ストリゴラクトンの簡易的なバイオアッセイ系の開発にも成功した.この研究の成果は,ストライガのもつストリゴラクトン受容体についての理解を促進し,将来的には,ストライガの発芽を制御する化合物の開発や耐性をもつ品種の育種への応用が考えられ,アフリカにおける食糧問題の解決への貢献が期待される.
はじめに
内生の植物ホルモンであるストリゴラクトンは,2008年,イネおよびシロイヌナズナにおいて枝分かれを制御するホルモンとしての機能が明らかにされた1).さらに,植物の根から放出されるストリゴラクトンはアーバスキュラー菌根菌との共生において重要なシグナルとしてはたらき2),じつに80%以上の植物がこれらのシグナルを窒素固定において必要としていると考えられている.そもそも,ストリゴラクトンはストライガとよばれる根寄生植物の発芽を誘導する化学物質として発見された3).多くの植物は貧栄養条件にさらされると根から多量のストリゴラクトンを放出し,それにより菌根の形成を促進すると考えられている4).つまり,ストライガはこの宿主植物からのシグナルをハイジャックして宿主植物が近くにいることを示すシグナルとして利用し,発芽していることが明らかにされた.実際,ストライガの被害は貧栄養条件にある農作地ほど大きく,ストライガはひとたびストリゴラクトンを受容すると発芽し,すぐに根を宿主植物の根に貫入して栄養を搾取する.ストライガによる被害は国連により食糧問題のなかでも最大の脅威と認定され,とくにアフリカの農業に大きな被害をもたらしている5).アフリカの耕作地のじつに2/3がすでにストライガにより汚染されており,その面積は40万km2にも及ぶ.これにより,少なくとも25カ国の1億人が被害をうけているとされている.
1.ストライガのもつストリゴラクトン受容体の同定
これまでの研究により,α/βヒドロラーゼがストリゴラクトンの受容において中心的な役割を担うことが示されている.たとえば,α/βヒドロラーゼファミリーに属するD14はイネにおいて枝分かれに重要なストリゴラクトン受容体である6,7).一方,D14と同じファミリーに属するHTLは,シロイヌナズナにおいて発芽に重要なはたらきを担うことが明らかにされた8).ストライガの発芽にはストリゴラクトンが必要なことから,HTLはストライガの発芽においても重要な役割を担うことが推測された.ストライガのRNA-seqデータベースを検索したところ,HTL遺伝子と相同性をもつ11個の遺伝子がみつかった9,10).根寄生植物であるストライガはその生育に宿主植物を必要とすることや,他殖性で種子を得るのには複数の個体が必要であることから,研究室で扱うのは非常に困難でより簡便な研究手法がもとめられていた.また,11個の遺伝子それぞれの機能を明らかにすることは,ストライガを使っていては困難であった.そこで,11個の遺伝子それぞれすべてをシロイヌナズナに別々に導入して発現させ,それぞれにコードされるタンパク質の生体における活性を調べた.その結果,9個のタンパク質が生体においてストリゴラクトン受容体としての活性をもっていた.また,筆者らにより開発されたストリゴラクトン様の活性をもつアゴニストを用いた機能解析により11),4個のタンパク質についてはアゴニストの構造に対する特異性が低下し,より多くのアゴニストに対して応答することが明らかにされた.それぞれの植物からそれぞれ異なる構造をもつストリゴラクトンが放出されることから,このような性質はストライガにとりより多くの宿主植物に対して応答するため重要であったのかもしれない.さらに,その4個のタンパク質のうちの1つShHTL7を発現したシロイヌナズナの種子は,10 pMのストリゴラクトンにも応答して発芽が誘導された.これは,耳かき1杯のストリゴラクトンを50 mプールに溶かした濃度に匹敵する.この実験による重要な発見は,ストライガにおいて超高感受性をもつストリゴラクトン受容体としてShHTL7を同定したこと,そして,ストリゴラクトン受容体の単独の進化のみによりこのような超高感受性の獲得を説明する可能性を示したことである.
2.ストライガのもつストリゴラクトン受容体の構造
ストライガのもつストリゴラクトン受容体はどのように超高感受性を獲得したのかを明らかにするため,結晶構造解析を試みた.HTL遺伝子と相同性をもつ11個の遺伝子にコードされるタンパク質すべての結晶化を試みたところ,重要な4個のタンパク質のうちの1つShHTL5の結晶化に成功した.その結晶構造を非寄生植物のHTLと比較したところ,ストリゴラクトンの結合するポケットのサイズが約2.5倍大きく,また,その入口の大きさも大きくなっていた(図1).ShHTL5の結晶構造から得られた情報をもとに,ストリゴラクトンに対し超高感受性をもつShHTL7のアミノ酸配列をみたところ,もっとも特徴的に異なる配列をもっていた.このことから,ストライガはストリゴラクトンの結合するポケットの部分に変異を起こすことにより,ストリゴラクトンに対する感受性を高めると同時に,さまざまな構造をもつストリゴラクトンにも応答できるという,いっけん相反する機能を同時に実現したと考えられた.この実験による重要な発見は,ストリゴラクトン受容体の構造を明らかにしてストリゴラクトンの結合するポケットの大きさの増大が重要であることを示し,また,それにかかわるアミノ酸残基の候補を明らかにしたことである.
3.ストライガのもつストリゴラクトン受容体のバイオセンサーとしての利用
ストライガの防除にむけての重要なファクターとして,化合物による発芽の制御および耐性をもつ品種の育種があげられる.しかしながら,さきに述べたとおり,ストライガは実験には不向きであること,また,有害な植物であることから,実際に研究を行える研究機関は限られてしまう.そこで,超高感受性をもつストリゴラクトン受容体であるShHTL7を利用してシロイヌナズナの種子を使ったバイオアッセイ系を構築できないか検討した.その結果,実際にイネを生育させ,その根のまわりにShHTL7を発現させたシロイヌナズナの種子を散布したところ,イネの根からの浸出液によりシロイヌナズナの発芽が促進された.このことから,このバイオアッセイ系により,少なくともイネにおける育種が可能であることが示された(図2).
おわりに
ストライガのもつストリゴラクトン受容体は非寄生植物のものと比較して,ストリゴラクトンの結合するポケットが大きくまた超高感受性へと変異していた.筆者らは,このようなストリゴラクトン受容体の変異がストライガのような根寄生植物の特殊な発芽様式および生活環を可能にしていると推測した.しかしながら,ストライガのもつストリゴラクトン受容体ShHTL7がどのようにしてストリゴラクトンに対する選択性のあいまいさと超高感受性とを同時に可能にしているのかはいまだに謎のままであり,今後のさらなる解析が必要である.また,実際にアフリカの大地に蔓延しているストライガの防除に取り組むためには,まだまだ多くの課題が残されている.しかしながら,今回の発見により研究者が標的とすべきストリゴラクトン受容体が明らかにされた.これにより,ストライガの防除にむけた研究が活発化し飛躍的に促進されることを期待したい.
文 献
- Umehara, M., Hanada, A., Yoshida, S. et al.: Inhibition of shoot branching by new terpenoid plant hormones. Nature, 455, 195-200 (2008)[PubMed]
- Akiyama, K., Matsuzaki, K. & Hayashi, H.: Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature, 435, 824-827 (2005)[PubMed]
- Cook, C. E., Whichard, L. P., Turner, B. et al.: Germination of witchweed (Striga lutea Lour.): isolation and properties of a potent stimulant. Science, 154, 1189-1190 (1966)[PubMed]
- Xie, X., Yoneyama, K. & Yoneyama, K.: The strigolactone story. Annu. Rev. Phytopathol., 48, 93-117 (2010)[PubMed]
- Ejeta, G.: The Striga scourge in Africa: a growing pandemic. in Integrating New Technologies for Striga Control (Ejeta, G. & Gressel, J. eds.), pp. 3-16, World Scientific Publishing, Singapore (2007)
- Arite, T., Umehara, M., Ishikawa, S. et al.: d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol., 50, 1416-1424 (2009)[PubMed]
- Hamiaux, C., Drummond, R. S., Janssen, B. J. et al.: DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone. Curr. Biol., 22, 2032-2036 (2012)[PubMed]
- Waters, M. T., Nelson, D. C., Scaffidi, A. et al.: Specialisation within the DWARF14 protein family confers distinct responses to karrikins and strigolactones in Arabidopsis. Development, 139, 1285-1295 (2012)[PubMed]
- Conn, E. C., Bythell-Douglas, R., Neumann, D. et al.: Convergent evolution of strigolactone perception enabled host detection in parasitic plants. Science, 349, 540-543 (2015)[PubMed]
- Tsuchiya, Y., Yoshimura, M., Sato, Y. et al.: Probing strigolactone receptors in Striga hermonthica with fluorescence. Science, 349, 864-868 (2015)[PubMed] [新着論文レビュー]
- Toh, S., Holbrook-Smith, D., Stokes, M. et al.: Detection of parasitic plant suicide germination compounds using a high-throughput Arabidopsis HTL/KAI2 strigolactone perception system. Chem. Biol., 21, 988-998 (2014)[PubMed]
著者プロフィール
略歴:2008年 明治大学大学院農学研究科 修了,同年 同 博士研究員を経て,2009年よりカナダToronto大学 博士研究員.
研究テーマ:ストリゴラクトンのシグナル伝達の機構.
© 2015 藤 茂雄 Licensed under CC 表示 2.1 日本
