寄生植物ストライガのもつストリゴラクトン受容体を可視化する
萩原伸也・吉村柾彦・土屋雄一朗
(名古屋大学トランスフォーマティブ生命分子研究所)
email:萩原伸也,土屋雄一朗
DOI: 10.7875/first.author.2015.106
Probing strigolactone receptors in Striga hermonthica with fluorescence.
Yuichiro Tsuchiya, Masahiko Yoshimura, Yoshikatsu Sato, Keiko Kuwata, Shigeo Toh, Duncan Holbrook-Smith, Hua Zhang, Peter McCourt, Kenichiro Itami, Toshinori Kinoshita, Shinya Hagihara
Science, 349, 864-868 (2015)
ストライガは穀物の根に寄生し養分や水分を奪う有害な植物である.その農業被害は年間1兆円をこえるとされ,アフリカにおける食糧問題の一因になっている.ストライガは植物ホルモンのひとつである宿主植物の放出するストリゴラクトンを感知して発芽する.このしくみを理解することはストライガの対策の糸口になると考えられているが,その鍵となるストリゴラクトン受容体はこれまで同定されていなかった.筆者らは,ストリゴラクトン受容体がはたらく過程を可視化できる分子を設計して合成し,これを活用することによりストライガのもつストリゴラクトン受容体を同定した.さらに,この分子を用いて,ストライガの発芽の過程においてストリゴラクトン受容体のはたらくようすを可視化することに成功した.この研究の成果はストライガの発芽を制御する薬剤の開発を加速し,将来的には,アフリカにおける食糧問題の解決への貢献が期待される.
アフリカにおいてストライガが猛威をふるっている.ストライガは特殊な根を生やしそれを穀物の根に侵入させて養分や水分を奪う寄生植物で,“魔女の雑草”とよばれている.とくにトウモロコシやイネなど主要な穀物に寄生し,収穫量を減らしたり,ときにはあたり一帯の田畑を全滅させたりする.こうした被害をうけている土地は日本の国土にも匹敵する40万km2にも及んでいる.土壌に広まったストライガの種は宿主植物が近くにくるまで,長いときは数十年も土の中で待ちつづける.そして,宿主植物の存在を感知すると発芽して寄生し,水分や養分を吸い取って成長する.種のまま休眠しているストライガを駆除するのはむずかしく,いちどストライガに汚染された土地では穀物の生産は困難になる.ストライガによる農業被害は年間1兆円をこえるといわれ,アフリカのみならず,食糧安全保障に対する世界的な脅威になっている1).
ストライガが宿主植物を認識するしくみにおいて鍵となるのは植物ホルモンのひとつストリゴラクトンである2).その構造は植物の種により少しずつ異なり,これまでに,少なくとも17種類のストリゴラクトンがみつかっている3).植物はリンや窒素などの養分が不足するとストリゴラクトンの合成量を増やす.ストリゴラクトンには植物の枝分かれを抑えるはたらきがあり,植物は自らの成長を抑制することにより養分の消費を抑える4).同時に,根からストリゴラクトンを放出し土壌に存在する菌根菌をよびよせる5).菌根菌はリンや窒素などの養分を植物に供給する共生菌の一種で,植物は菌根菌との共生により貧栄養状態を脱しようとする.このように,植物は環境に応答するためストリゴラクトンを合成し土壌へと放出する.ストライガはこのストリゴラクトンを感知して発芽する.この機構を分子レベルで解明することができればストライガの発芽を制御する分子の開発につながるが,長いあいだ,ストライガのもつストリゴラクトン受容体は同定されていなかった.
では,枝分かれの制御におけるストリゴラクトン受容体は何か.最近,イネ,シロイヌナズナ,ペチュニアにおける研究から,D14ファミリータンパク質であることが明らかにされた6).D14ファミリータンパク質はα/βヒドロラーゼフォールドスーパーファミリーに属し,ストリゴラクトンと結合してシグナルを伝達するとともに,ストリゴラクトンを加水分解する活性をもつ7).発芽と分枝はいっけんかなり異なる生理現象であるが,非天然物質を含め,これまでに知られているストライガの発芽を刺激する物質はD14ファミリータンパク質に受容されることから,ストライガのもつストリゴラクトン受容体はD14と類似したリガンド選択性をもつと考えられる8).
一方,山火事の煙に含まれるカリキンなど,これまで,非寄生植物において発芽を誘導する物質もいくつかみつかっている9,10).これらの物質はD14のホモログであるHTLに受容されるが,ストライガの発芽を誘導しない.すなわち,ストライガのもつストリゴラクトン受容体は,D14と類似したリガンド選択性と,HTLと類似した機能の2つの性質をあわせもつと推定された.
植物科学にかぎらず,タンパク質の生理機能を調べるには変異体を用いるなど遺伝学的な手法をとることが多い.しかし,ストライガは寄生しないと育たないため実験室における生育がむずかしく,遺伝学的な手法の適応が困難であった.このため,ストライガにストリゴラクトン受容体の候補となるタンパク質が存在しても,これらのタンパク質の機能をin vivoで確かめる効率的な手法がなかった.これが,ストライガにおいてストリゴラクトン受容体の同定が進まなかった要因であった.
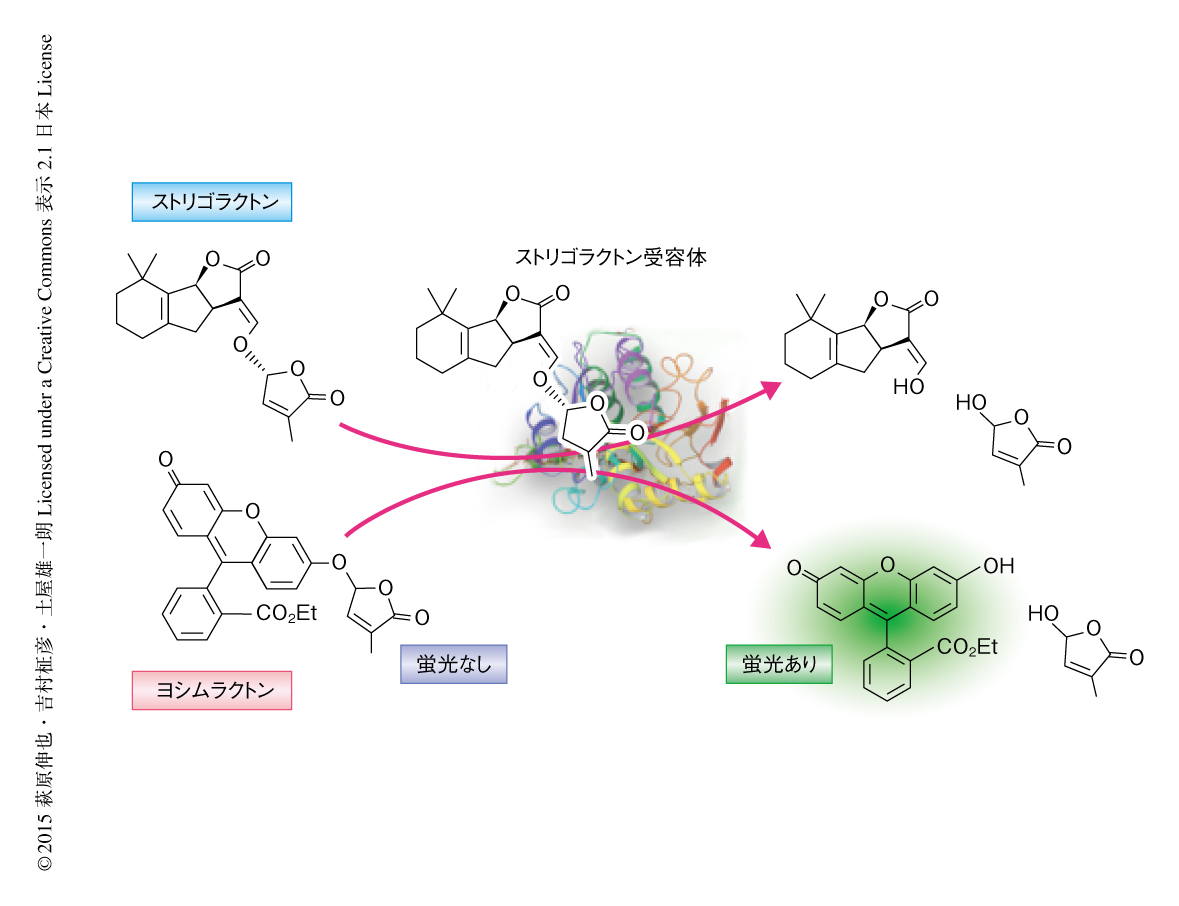
そこで,遺伝学的な手法を用いずにストリゴラクトン受容体の機能を評価する手法を開発した.D14がストリゴラクトンを加水分解することに着目し,D14により加水分解されると蛍光を発するストリゴラクトン様の分子を合成した.この分子を組換えD14にくわえたところ,蛍光の経時的な増加が観測され,設計どおりD14と結合し加水分解されることがわかった.一方,この分子はHTLには加水分解されなかったため,天然のストリゴラクトンと類似した受容体に対する選択性をもつと考えられた.また,シロイヌナズナにおいて分枝を抑制する効果がみられたことから,この分子はin vivoにおいて発蛍光性ストリゴラクトンとして機能することが確認され,ヨシムラクトンと命名された(図1).
ストライガの種子をヨシムラクトンにより処理したところ,発芽を誘導する効果がみられた.さらに,発芽した種子において蛍光が観測されたことから,ヨシムラクトンはストライガにおいても発蛍光性ストリゴラクトンとして機能することが示された.逆にいうと,ストライガのもつストリゴラクトン受容体はヨシムラクトンを加水分解する活性をもっていた.
一方,ストライガのRNA-seqデータベースを検索したところ,D14やHTLと相同性をもつ12個のタンパク質がみつかった.これらのタンパク質それぞれに対しヨシムラクトンを作用させたところ,10個において蛍光が観測された.すなわち,これらのタンパク質がストライガのもつストリゴラクトン受容体であると考えられた.これらのタンパク質はストライガに特有のものでありHTLから派生していた.つまり,非寄生植物において発芽にかかわる受容体であるHTLが,寄生植物としての進化の過程においてストリゴラクトンを感知するようになったものと考えられた.
なぜ,10種類ものストリゴラクトン受容体が必要なのか.受容体によるヨシムラクトンの加水分解活性を指標として,おのおのの受容体のさまざまなストリゴラクトンに対する親和性を見積もった.その結果,受容体ごとに強く結合するストリゴラクトンの種類に違いがみられた.このことから,ストライガはさまざまな受容体をもつことにより多くの種類の宿主植物に寄生できるよう進化してきたものと考えられた.
ヨシムラクトンをくわえてストライガの発芽するようすを連続撮影したところ,最初に根の側の先端において強く光り,それがストライガの種の全体へ波のように拡がったのち,いったん光が消え,ふたたび光りながら根が伸びていくのが観察された.この光は,ストリゴラクトンの受容のダイナミクスを表わしていた.発芽した種子のすべてにおいて同じ挙動がみられたことから,このダイナミクスはストライガの発芽と強く関連していると考えられた.ここにエチレン生合成阻害剤をくわえると蛍光が大幅に減少したことから,エチレンがストリゴラクトンの受容を促進していることが明らかにされた.一方,以前から,ストリゴラクトンがエチレンの生合成を誘導することが知られており,今回の結果とあわせると,ストリゴラクトンとエチレンは互いにシグナルを増幅しあっていることになった.ストライガはこのシグナルの増幅の機構により土壌に存在するわずかなストリゴラクトンを検出していると考えられた.ストライガの種は根の先端で宿主植物が近くにいるのを感知し,その情報を種の全体に伝達して休眠状態を解除し,宿主植物にむけ根を伸ばしていく.今回の研究の成果から,ストライガは宿主植物の放出するストリゴラクトンを巧妙に活用して寄生にかかわる一連の過程を制御していることが明らかにされた.
有機合成化学者と植物生物学者がそれぞれの分野の壁をこえて共同研究をすることにより,ストライガが宿主植物を感知して発芽するしくみの一端が明らかにされた.ストリゴラクトン受容体が明らかになったいま,ストライガの発芽を制御する薬剤の探索が飛躍的に加速すると考えられる.今回,開発されたヨシムラクトンはこのような薬剤の探索においても効果を発揮し,ストライガによる農業被害の解決にさらに貢献すると期待される.
略歴:2003年 京都大学大学院工学研究科 修了,同年 理化学研究所 研究員,2007年 スイスGeneva大学 博士研究員,2008年 東北大学多元物質科学研究所 助教を経て,2013年より名古屋大学トランスフォーマティブ生命分子研究所 特任准教授.
吉村 柾彦(Masahiko Yoshimura)
名古屋大学大学院理学研究科博士課程 在学中.
土屋 雄一朗(Yuichiro Tsuchiya)
名古屋大学トランスフォーマティブ生命分子研究所 博士研究員.
© 2015 萩原伸也・吉村柾彦・土屋雄一朗 Licensed under CC 表示 2.1 日本
(名古屋大学トランスフォーマティブ生命分子研究所)
email:萩原伸也,土屋雄一朗
DOI: 10.7875/first.author.2015.106
Probing strigolactone receptors in Striga hermonthica with fluorescence.
Yuichiro Tsuchiya, Masahiko Yoshimura, Yoshikatsu Sato, Keiko Kuwata, Shigeo Toh, Duncan Holbrook-Smith, Hua Zhang, Peter McCourt, Kenichiro Itami, Toshinori Kinoshita, Shinya Hagihara
Science, 349, 864-868 (2015)
要 約
ストライガは穀物の根に寄生し養分や水分を奪う有害な植物である.その農業被害は年間1兆円をこえるとされ,アフリカにおける食糧問題の一因になっている.ストライガは植物ホルモンのひとつである宿主植物の放出するストリゴラクトンを感知して発芽する.このしくみを理解することはストライガの対策の糸口になると考えられているが,その鍵となるストリゴラクトン受容体はこれまで同定されていなかった.筆者らは,ストリゴラクトン受容体がはたらく過程を可視化できる分子を設計して合成し,これを活用することによりストライガのもつストリゴラクトン受容体を同定した.さらに,この分子を用いて,ストライガの発芽の過程においてストリゴラクトン受容体のはたらくようすを可視化することに成功した.この研究の成果はストライガの発芽を制御する薬剤の開発を加速し,将来的には,アフリカにおける食糧問題の解決への貢献が期待される.
はじめに
アフリカにおいてストライガが猛威をふるっている.ストライガは特殊な根を生やしそれを穀物の根に侵入させて養分や水分を奪う寄生植物で,“魔女の雑草”とよばれている.とくにトウモロコシやイネなど主要な穀物に寄生し,収穫量を減らしたり,ときにはあたり一帯の田畑を全滅させたりする.こうした被害をうけている土地は日本の国土にも匹敵する40万km2にも及んでいる.土壌に広まったストライガの種は宿主植物が近くにくるまで,長いときは数十年も土の中で待ちつづける.そして,宿主植物の存在を感知すると発芽して寄生し,水分や養分を吸い取って成長する.種のまま休眠しているストライガを駆除するのはむずかしく,いちどストライガに汚染された土地では穀物の生産は困難になる.ストライガによる農業被害は年間1兆円をこえるといわれ,アフリカのみならず,食糧安全保障に対する世界的な脅威になっている1).
1.ストライガの発芽とストリゴラクトン
ストライガが宿主植物を認識するしくみにおいて鍵となるのは植物ホルモンのひとつストリゴラクトンである2).その構造は植物の種により少しずつ異なり,これまでに,少なくとも17種類のストリゴラクトンがみつかっている3).植物はリンや窒素などの養分が不足するとストリゴラクトンの合成量を増やす.ストリゴラクトンには植物の枝分かれを抑えるはたらきがあり,植物は自らの成長を抑制することにより養分の消費を抑える4).同時に,根からストリゴラクトンを放出し土壌に存在する菌根菌をよびよせる5).菌根菌はリンや窒素などの養分を植物に供給する共生菌の一種で,植物は菌根菌との共生により貧栄養状態を脱しようとする.このように,植物は環境に応答するためストリゴラクトンを合成し土壌へと放出する.ストライガはこのストリゴラクトンを感知して発芽する.この機構を分子レベルで解明することができればストライガの発芽を制御する分子の開発につながるが,長いあいだ,ストライガのもつストリゴラクトン受容体は同定されていなかった.
2.非寄生植物のもつストリゴラクトン受容体
では,枝分かれの制御におけるストリゴラクトン受容体は何か.最近,イネ,シロイヌナズナ,ペチュニアにおける研究から,D14ファミリータンパク質であることが明らかにされた6).D14ファミリータンパク質はα/βヒドロラーゼフォールドスーパーファミリーに属し,ストリゴラクトンと結合してシグナルを伝達するとともに,ストリゴラクトンを加水分解する活性をもつ7).発芽と分枝はいっけんかなり異なる生理現象であるが,非天然物質を含め,これまでに知られているストライガの発芽を刺激する物質はD14ファミリータンパク質に受容されることから,ストライガのもつストリゴラクトン受容体はD14と類似したリガンド選択性をもつと考えられる8).
一方,山火事の煙に含まれるカリキンなど,これまで,非寄生植物において発芽を誘導する物質もいくつかみつかっている9,10).これらの物質はD14のホモログであるHTLに受容されるが,ストライガの発芽を誘導しない.すなわち,ストライガのもつストリゴラクトン受容体は,D14と類似したリガンド選択性と,HTLと類似した機能の2つの性質をあわせもつと推定された.
3.加水分解されると蛍光を発するストリゴラクトン様の分子の設計
植物科学にかぎらず,タンパク質の生理機能を調べるには変異体を用いるなど遺伝学的な手法をとることが多い.しかし,ストライガは寄生しないと育たないため実験室における生育がむずかしく,遺伝学的な手法の適応が困難であった.このため,ストライガにストリゴラクトン受容体の候補となるタンパク質が存在しても,これらのタンパク質の機能をin vivoで確かめる効率的な手法がなかった.これが,ストライガにおいてストリゴラクトン受容体の同定が進まなかった要因であった.
そこで,遺伝学的な手法を用いずにストリゴラクトン受容体の機能を評価する手法を開発した.D14がストリゴラクトンを加水分解することに着目し,D14により加水分解されると蛍光を発するストリゴラクトン様の分子を合成した.この分子を組換えD14にくわえたところ,蛍光の経時的な増加が観測され,設計どおりD14と結合し加水分解されることがわかった.一方,この分子はHTLには加水分解されなかったため,天然のストリゴラクトンと類似した受容体に対する選択性をもつと考えられた.また,シロイヌナズナにおいて分枝を抑制する効果がみられたことから,この分子はin vivoにおいて発蛍光性ストリゴラクトンとして機能することが確認され,ヨシムラクトンと命名された(図1).
4.発蛍光性ストリゴラクトンを用いたストライガの発芽の解析
ストライガの種子をヨシムラクトンにより処理したところ,発芽を誘導する効果がみられた.さらに,発芽した種子において蛍光が観測されたことから,ヨシムラクトンはストライガにおいても発蛍光性ストリゴラクトンとして機能することが示された.逆にいうと,ストライガのもつストリゴラクトン受容体はヨシムラクトンを加水分解する活性をもっていた.
一方,ストライガのRNA-seqデータベースを検索したところ,D14やHTLと相同性をもつ12個のタンパク質がみつかった.これらのタンパク質それぞれに対しヨシムラクトンを作用させたところ,10個において蛍光が観測された.すなわち,これらのタンパク質がストライガのもつストリゴラクトン受容体であると考えられた.これらのタンパク質はストライガに特有のものでありHTLから派生していた.つまり,非寄生植物において発芽にかかわる受容体であるHTLが,寄生植物としての進化の過程においてストリゴラクトンを感知するようになったものと考えられた.
なぜ,10種類ものストリゴラクトン受容体が必要なのか.受容体によるヨシムラクトンの加水分解活性を指標として,おのおのの受容体のさまざまなストリゴラクトンに対する親和性を見積もった.その結果,受容体ごとに強く結合するストリゴラクトンの種類に違いがみられた.このことから,ストライガはさまざまな受容体をもつことにより多くの種類の宿主植物に寄生できるよう進化してきたものと考えられた.
5.ストライガの発芽におけるストリゴラクトンの受容のダイナミクス
ヨシムラクトンをくわえてストライガの発芽するようすを連続撮影したところ,最初に根の側の先端において強く光り,それがストライガの種の全体へ波のように拡がったのち,いったん光が消え,ふたたび光りながら根が伸びていくのが観察された.この光は,ストリゴラクトンの受容のダイナミクスを表わしていた.発芽した種子のすべてにおいて同じ挙動がみられたことから,このダイナミクスはストライガの発芽と強く関連していると考えられた.ここにエチレン生合成阻害剤をくわえると蛍光が大幅に減少したことから,エチレンがストリゴラクトンの受容を促進していることが明らかにされた.一方,以前から,ストリゴラクトンがエチレンの生合成を誘導することが知られており,今回の結果とあわせると,ストリゴラクトンとエチレンは互いにシグナルを増幅しあっていることになった.ストライガはこのシグナルの増幅の機構により土壌に存在するわずかなストリゴラクトンを検出していると考えられた.ストライガの種は根の先端で宿主植物が近くにいるのを感知し,その情報を種の全体に伝達して休眠状態を解除し,宿主植物にむけ根を伸ばしていく.今回の研究の成果から,ストライガは宿主植物の放出するストリゴラクトンを巧妙に活用して寄生にかかわる一連の過程を制御していることが明らかにされた.
おわりに
有機合成化学者と植物生物学者がそれぞれの分野の壁をこえて共同研究をすることにより,ストライガが宿主植物を感知して発芽するしくみの一端が明らかにされた.ストリゴラクトン受容体が明らかになったいま,ストライガの発芽を制御する薬剤の探索が飛躍的に加速すると考えられる.今回,開発されたヨシムラクトンはこのような薬剤の探索においても効果を発揮し,ストライガによる農業被害の解決にさらに貢献すると期待される.
文 献
- Ejeta, G.: The Striga scourge in Africa: a growing pandemic. in Integrating New Technologies for Striga Control (Ejeta, G. & Gressel, J. eds.), pp. 3-16, World Scientific Publishing, Singapore (2007)
- Cook, C. E., Whichard, L. P., Turner, B. et al.: Germination of witchweed (Striga lutea Lour.): isolation and properties of a potent stimulant. Science, 154, 1189-1190 (1966)[PubMed]
- Xie, X., Yoneyama, K. & Yoneyama, K.: The strigolactone story. Annu. Rev. Phytopathol., 48, 93-117 (2010)[PubMed]
- Umehara, M., Hanada, A., Yoshida, S. et al.: Inhibition of shoot branching by new terpenoid plant hormones. Nature, 455, 195-200 (2008)[PubMed]
- Akiyama, K., Matsuzaki, K. & Hayashi, H.: Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature, 435, 824-827 (2005)[PubMed]
- Arite, T., Umehara, M., Ishikawa, S. et al.: d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol., 50, 1416-1424 (2009)[PubMed]
- Hamiaux, C., Drummond, R. S., Janssen, B. J. et al.: DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone. Curr. Biol., 22, 2032-2036 (2012)[PubMed]
- Fukui, K., Ito, S., Ueno, K. et al.: New branching inhibitors and their potential as strigolactone mimics in rice. Bioorg. Med. Chem. Lett., 21, 4905-4908 (2011)[PubMed]
- Waters, M. T., Nelson, D. C., Scaffidi, A. et al.: Specialisation within the DWARF14 protein family confers distinct responses to karrikins and strigolactones in Arabidopsis. Development, 139, 1285-1295 (2012)[PubMed]
- Tsuchiya, Y., Vidaurre, D., Toh, S. et al.: A small-molecule screen identifies new functions for the plant hormone strigolactone. Nat. Chem. Biol., 6, 741-749 (2010)[PubMed] [新着論文レビュー]
著者プロフィール
略歴:2003年 京都大学大学院工学研究科 修了,同年 理化学研究所 研究員,2007年 スイスGeneva大学 博士研究員,2008年 東北大学多元物質科学研究所 助教を経て,2013年より名古屋大学トランスフォーマティブ生命分子研究所 特任准教授.
吉村 柾彦(Masahiko Yoshimura)
名古屋大学大学院理学研究科博士課程 在学中.
土屋 雄一朗(Yuichiro Tsuchiya)
名古屋大学トランスフォーマティブ生命分子研究所 博士研究員.
© 2015 萩原伸也・吉村柾彦・土屋雄一朗 Licensed under CC 表示 2.1 日本
