hnRNP UはX染色体へのXist RNAの局在を制御する
長谷川優子・中川真一
(理化学研究所基幹研究所 中川RNA生物学研究室)
email:長谷川優子,中川真一
DOI: 10.7875/first.author.2010.035
The matrix protein hnRNP U is required for chromosomal localization of Xist RNA.
Yuko Hasegawa, Neil Brockdorff, Shinji Kawano, Kimiko Tsutui, Ken Tsutui, Shinichi Nakagawa
Developmental Cell, 19, 469-476 (2010)
ノンコーディングRNAの一種であるXist RNAはX染色体の不活性化における必須因子である.通常のmRNAは細胞質へと輸送されてタンパク質に翻訳されるが,このノンコーディングRNAは核内にとどまってX染色体の全体を覆うように局在化し,クロマチン制御因子であるPRC2の足場としてはたらくものと考えられている.Xist RNAをX染色体へ局在化させる機構は不明であったが,筆者らは,RNA結合タンパク質hnRNP UがXist RNAのX染色体への局在化に必須であることを見い出した.hnRNP UがないとX染色体の不活性化も正常に起こらない.今回の研究は,ノンコーディングRNAが関与するエピジェネティックな遺伝子発現の制御機構の新たな切り口を示すものである.
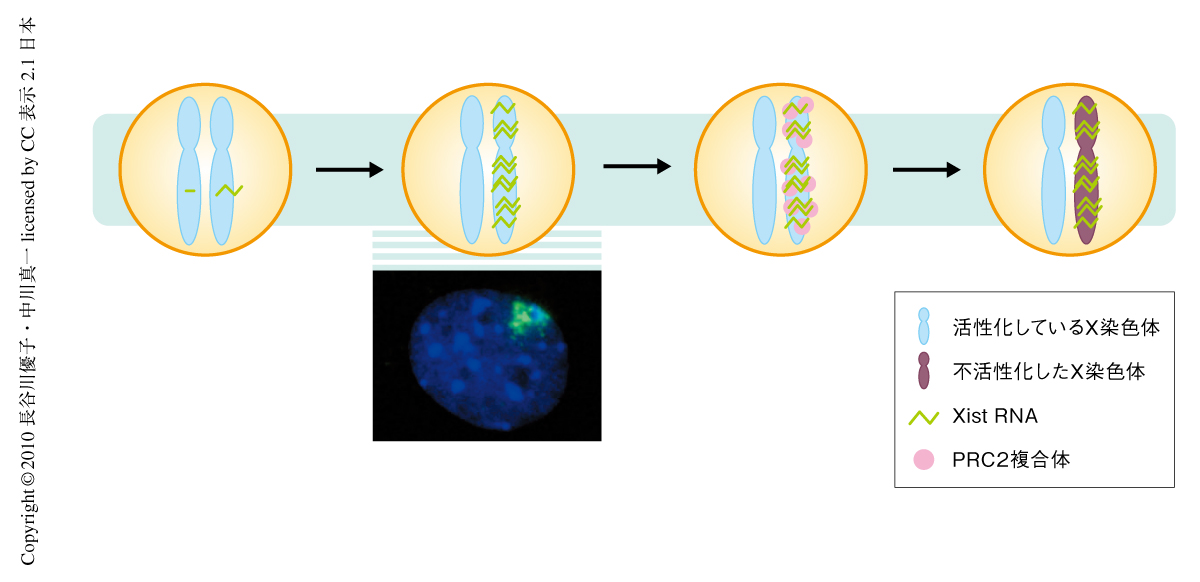
哺乳類の雌は自身のもつ2本のX染色体のうちの1本をほぼ全域にわたって転写不活性な状態に保っている.これはX染色体の不活性化とよばれる現象であり,エピジェネティックな遺伝子発現制御の一例としてよく知られている.非常に広範囲に及ぶこの遺伝子発現制御をささえる因子のひとつはXist RNA(inactive X specific transcripts RNA)である1-3).Xist RNAの遺伝子座はX染色体にあり,将来的に不活性化されるほうのX染色体から強い発現を示す.転写されたXist RNAはX染色体の全体に広がってそこにとどまる.クロマチン制御因子であるPRC2(polycomb repressor complex 2)複合体はXist RNA依存的にX染色体へとリクルートされることから4),Xist RNAはPRC2複合体の足場として機能しているものと考えられている(図1).したがって,Xist RNAの発現と局在化はX染色体の不活性化の引き金という位置づけにあって非常に重要な過程であるが,これをどのような分子機構がささえているのかは不明であった.
筆者らは,mini siRNAライブラリーを用いてRNA結合タンパク質のノックダウン実験を行い,Xist RNAの局在制御に関与する因子を探索した.その結果,hnRNP U(別名SAF-AあるいはSP120)を候補因子として見い出した.hnRNP UをノックダウンするとXist RNAはX染色体にとどまらず,核全体に散らばるような局在を示すようになった.局在の変化した原因としてXist RNAの“局在化シグナル”(Xist RNAがX染色体に局在化するのに必要とされるXist RNAの領域)が5),hnRNP Uのノックダウンによってなんらかの理由で欠けていることなどが考えられたが,ノーザンブロッティング解析により局在の変化したXist RNAにも局在化シグナルは存在していることが明らかになった.これは,hnRNP Uがない状況ではXist RNA自体は局在化シグナルを保有しているものの正しい局在がさまたげられていることを示唆していた.したがって,hnRNP UはXist RNAをX染色体に繋留する機能を担っていることが予想された.
つづいて,hnRNP UによるXist RNAの局在制御の詳細を明らかにするためドメイン解析を行った.hnRNP UはDNA結合ドメインであるSAFドメイン,機能未知なSPRYドメイン,RNA結合ドメインであるRGGドメインからなる6-8).全長のhnRNP Uとそれぞれのドメインを欠いたhnRNP Uコンストラクトを作製してこれを細胞に発現させ,Xist RNAとの相互作用を架橋-免疫沈降(cross linking immunoprecipitation:CLIP)法によって調べた.その結果,hnRNP UとXist RNAとは相互作用しており,RNA結合能をもつRGGドメインがこれを担っていることがわかった.また,Xist RNA側のhnRNP Uとの相互作用領域は局在化シグナルとおおよそ重なることも明らかになった.
以上の実験より明らかになった,hnRNP UのRNA結合ドメインを介したXist RNAとの相互作用が局在の制御に十分であるかどうかをレスキュー実験により確かめた.CLIP法で使用したコンストラクトにはsiRNA耐性型の変異が導入されており,これらを発現している細胞をsiRNAで処理すると内在性のhnRNP Uのみをノックダウンできる.このときのXist RNAの局在変化を観察したところ,予想どおり,RNA結合ドメイン欠損型のhnRNP UはXist RNAの局在変化を補完することができなかった.しかしながら,Xist RNAとの相互作用能をもつはずのDNA結合ドメイン欠損型のhnRNP Uもやはり局在変化を補うことができなかった.そこで,DNA結合ドメインとRNA結合ドメインの両方をもつコンストラクトを作製し,再度,レスキュー実験を行ったところ,このコンストラクトはXist RNAの局在を正常に保つ能力をもっていた.したがって,hnRNP UによるXist RNAの局在制御にはDNA結合ドメインとRNA結合ドメインの両方が必要であることがわかった.
最後に,hnRNP UのノックダウンによりXist RNAの局在が異常になることでX染色体の不活性化にどのような影響がでるかを調べた.X染色体の不活性化には,不活性化したX染色体を新たに確立する“開始”の段階と,いちど確立された不活性化したX染色体の“維持”に相当する段階とがあるが,Xist RNAがおもに関与しているのは“開始”であることが知られている9).X染色体にコードされている遺伝子(X-linked gene)の転写抑制を指標として,ES細胞の分化系を用いてhnRNP Uノックダウンによる“開始”過程への影響を調べた.その結果,hnRNP Uをノックダウンすると,片側のX染色体からのXist RNAの発現促進は見受けられるが,X染色体の全体への広がりが起こらずX染色体にコードされている遺伝子の転写抑制の確立が阻害されていた.一方,“維持”に相当する段階にあるマウス初代繊維芽細胞ではhnRNP Uをノックダウンしても転写抑制状態の解除は起こらなかった.また,ヒストン修飾に関しては,PRC2複合体の構成成分であるEzh2のX染色体へのリクルートが顕著に低下し,ES細胞分化系においてもマウス初代繊維芽細胞においても,X染色体へのヒストンH3の27番目のリジン残基のトリメチル化の蓄積がみられなくなっていた.以上の表現型はXistを欠損させた際のX染色体の不活性化への影響と同等のものであり,hnRNP UノックダウンによるX染色体の不活性化への影響はXist RNAの正常な機能が損なわれたことに起因することを強く示唆した.
以上の結果をまとめると,つぎのようなモデルが想像される.hnRNP UはSAFドメインを介してX染色体と相互作用し,RGGドメインを介してXist RNAと相互作用することで,あたかもXist RNAをX染色体に貼り付ける糊のように機能してX染色体の不活性化を制御する,というものである(図2).クロマチンのhnRNP U結合領域はマトリックス結合領域(matrix attachment region:MAR)とよばれるATに富む領域として以前から知られている.おそらく,X染色体にはあらかじめhnRNP Uが結合しており,Xist RNAが発現するやこれを繋留してX染色体から離れないようにしているのだろう.また,複数のグループによって,不活性化したX染色体にhnRNP Uが蓄積していることも示されている10,11).ただし,この蓄積はXist RNAに依存すること,ES細胞が分化してから観察されるようになることなどから,筆者らが提唱するモデルとは趣が異なるように思われる.hnRNP Uは核酸の存在により多量体化することが知られており,別グループらによって観察されたX染色体へのhnRNP Uの蓄積は,Xist RNAが初期量の(それほど多くはない)hnRNP Uによって繋留された“結果”としてみられる現象なのかもしれない.

最後に,“hnRNP U is required for chromosomal localization of Xist RNA”というこの論文のタイトルをひねくれた角度からみてみようと思う.このタイトルは,X染色体を“動かないもの”,Xist RNAを“動くもの”と仮定したときに成立する.かたや,Edithらは,2006年に発表した論文“A novel role for Xist RNA in the formation of a repressive nuclear compartment into which gene are recruited when silenced”において,Xist RNAが形成する“nuclear compartment”に発現抑制されるべき遺伝子が引き込まれる,と主張した12).“動くもの”と位置づけている対象が,筆者らと逆なのである.Edithらの主張に筆者らの今回の研究をすり合わせてみると,hnRNP Uのない状況で阻害されたのはこの“nuclear compartment”の形成である,といういい方ができる.実験結果をふまえると,hnRNP UのDNA結合能(すなわち,おそらくX染色体との相互作用)もXist RNAの正常な局在化に必要であるので,Xist RNAがX染色体の位置と無関係に勝手な場所に“nuclear compartment”を形成しているわけではなく,相互に動向を制御しあっているのであろう.不活性化したX染色体とは巨大なRNPなのであろうか.
近年,Xist RNA様の機能をもつ複数のノンコーディングRNAがみつかってきており,ノンコーディングRNAとエピジェネティクスとの関連が注目されている.この研究がこの分野の発展および理解に貢献する道標のひとつとなることを期待する.
略歴:東京大学大学院工学系研究科博士課程 在学中.
研究テーマ:核内ノンコーディングRNAの制御機構.
抱負:冷静に熱い仕事をする.
中川 真一(Shinichi Nakagawa)
理化学研究所基幹研究所 准主任研究員.
研究室URL:http://www.riken.jp/lab-www/nakagawa/index_j.html
© 2010 長谷川優子・中川真一 Licensed under CC 表示 2.1 日本
(理化学研究所基幹研究所 中川RNA生物学研究室)
email:長谷川優子,中川真一
DOI: 10.7875/first.author.2010.035
The matrix protein hnRNP U is required for chromosomal localization of Xist RNA.
Yuko Hasegawa, Neil Brockdorff, Shinji Kawano, Kimiko Tsutui, Ken Tsutui, Shinichi Nakagawa
Developmental Cell, 19, 469-476 (2010)
要 約
ノンコーディングRNAの一種であるXist RNAはX染色体の不活性化における必須因子である.通常のmRNAは細胞質へと輸送されてタンパク質に翻訳されるが,このノンコーディングRNAは核内にとどまってX染色体の全体を覆うように局在化し,クロマチン制御因子であるPRC2の足場としてはたらくものと考えられている.Xist RNAをX染色体へ局在化させる機構は不明であったが,筆者らは,RNA結合タンパク質hnRNP UがXist RNAのX染色体への局在化に必須であることを見い出した.hnRNP UがないとX染色体の不活性化も正常に起こらない.今回の研究は,ノンコーディングRNAが関与するエピジェネティックな遺伝子発現の制御機構の新たな切り口を示すものである.
はじめに
哺乳類の雌は自身のもつ2本のX染色体のうちの1本をほぼ全域にわたって転写不活性な状態に保っている.これはX染色体の不活性化とよばれる現象であり,エピジェネティックな遺伝子発現制御の一例としてよく知られている.非常に広範囲に及ぶこの遺伝子発現制御をささえる因子のひとつはXist RNA(inactive X specific transcripts RNA)である1-3).Xist RNAの遺伝子座はX染色体にあり,将来的に不活性化されるほうのX染色体から強い発現を示す.転写されたXist RNAはX染色体の全体に広がってそこにとどまる.クロマチン制御因子であるPRC2(polycomb repressor complex 2)複合体はXist RNA依存的にX染色体へとリクルートされることから4),Xist RNAはPRC2複合体の足場として機能しているものと考えられている(図1).したがって,Xist RNAの発現と局在化はX染色体の不活性化の引き金という位置づけにあって非常に重要な過程であるが,これをどのような分子機構がささえているのかは不明であった.
1.RNA結合タンパク質hnRNP UはXist RNAのX染色体への局在化に必要である
筆者らは,mini siRNAライブラリーを用いてRNA結合タンパク質のノックダウン実験を行い,Xist RNAの局在制御に関与する因子を探索した.その結果,hnRNP U(別名SAF-AあるいはSP120)を候補因子として見い出した.hnRNP UをノックダウンするとXist RNAはX染色体にとどまらず,核全体に散らばるような局在を示すようになった.局在の変化した原因としてXist RNAの“局在化シグナル”(Xist RNAがX染色体に局在化するのに必要とされるXist RNAの領域)が5),hnRNP Uのノックダウンによってなんらかの理由で欠けていることなどが考えられたが,ノーザンブロッティング解析により局在の変化したXist RNAにも局在化シグナルは存在していることが明らかになった.これは,hnRNP Uがない状況ではXist RNA自体は局在化シグナルを保有しているものの正しい局在がさまたげられていることを示唆していた.したがって,hnRNP UはXist RNAをX染色体に繋留する機能を担っていることが予想された.
2.hnRNP UはRNA結合ドメインを介してXist RNAと相互作用する
つづいて,hnRNP UによるXist RNAの局在制御の詳細を明らかにするためドメイン解析を行った.hnRNP UはDNA結合ドメインであるSAFドメイン,機能未知なSPRYドメイン,RNA結合ドメインであるRGGドメインからなる6-8).全長のhnRNP Uとそれぞれのドメインを欠いたhnRNP Uコンストラクトを作製してこれを細胞に発現させ,Xist RNAとの相互作用を架橋-免疫沈降(cross linking immunoprecipitation:CLIP)法によって調べた.その結果,hnRNP UとXist RNAとは相互作用しており,RNA結合能をもつRGGドメインがこれを担っていることがわかった.また,Xist RNA側のhnRNP Uとの相互作用領域は局在化シグナルとおおよそ重なることも明らかになった.
3.Xist RNAの局在制御にはhnRNP UのDNA結合ドメインとRNA結合ドメインの両方が必要である
以上の実験より明らかになった,hnRNP UのRNA結合ドメインを介したXist RNAとの相互作用が局在の制御に十分であるかどうかをレスキュー実験により確かめた.CLIP法で使用したコンストラクトにはsiRNA耐性型の変異が導入されており,これらを発現している細胞をsiRNAで処理すると内在性のhnRNP Uのみをノックダウンできる.このときのXist RNAの局在変化を観察したところ,予想どおり,RNA結合ドメイン欠損型のhnRNP UはXist RNAの局在変化を補完することができなかった.しかしながら,Xist RNAとの相互作用能をもつはずのDNA結合ドメイン欠損型のhnRNP Uもやはり局在変化を補うことができなかった.そこで,DNA結合ドメインとRNA結合ドメインの両方をもつコンストラクトを作製し,再度,レスキュー実験を行ったところ,このコンストラクトはXist RNAの局在を正常に保つ能力をもっていた.したがって,hnRNP UによるXist RNAの局在制御にはDNA結合ドメインとRNA結合ドメインの両方が必要であることがわかった.
4.Xist RNAの局在変化が及ぼすX染色体の不活性化への影響
最後に,hnRNP UのノックダウンによりXist RNAの局在が異常になることでX染色体の不活性化にどのような影響がでるかを調べた.X染色体の不活性化には,不活性化したX染色体を新たに確立する“開始”の段階と,いちど確立された不活性化したX染色体の“維持”に相当する段階とがあるが,Xist RNAがおもに関与しているのは“開始”であることが知られている9).X染色体にコードされている遺伝子(X-linked gene)の転写抑制を指標として,ES細胞の分化系を用いてhnRNP Uノックダウンによる“開始”過程への影響を調べた.その結果,hnRNP Uをノックダウンすると,片側のX染色体からのXist RNAの発現促進は見受けられるが,X染色体の全体への広がりが起こらずX染色体にコードされている遺伝子の転写抑制の確立が阻害されていた.一方,“維持”に相当する段階にあるマウス初代繊維芽細胞ではhnRNP Uをノックダウンしても転写抑制状態の解除は起こらなかった.また,ヒストン修飾に関しては,PRC2複合体の構成成分であるEzh2のX染色体へのリクルートが顕著に低下し,ES細胞分化系においてもマウス初代繊維芽細胞においても,X染色体へのヒストンH3の27番目のリジン残基のトリメチル化の蓄積がみられなくなっていた.以上の表現型はXistを欠損させた際のX染色体の不活性化への影響と同等のものであり,hnRNP UノックダウンによるX染色体の不活性化への影響はXist RNAの正常な機能が損なわれたことに起因することを強く示唆した.
5.hnRNP UによるXist RNAの局在制御モデル
以上の結果をまとめると,つぎのようなモデルが想像される.hnRNP UはSAFドメインを介してX染色体と相互作用し,RGGドメインを介してXist RNAと相互作用することで,あたかもXist RNAをX染色体に貼り付ける糊のように機能してX染色体の不活性化を制御する,というものである(図2).クロマチンのhnRNP U結合領域はマトリックス結合領域(matrix attachment region:MAR)とよばれるATに富む領域として以前から知られている.おそらく,X染色体にはあらかじめhnRNP Uが結合しており,Xist RNAが発現するやこれを繋留してX染色体から離れないようにしているのだろう.また,複数のグループによって,不活性化したX染色体にhnRNP Uが蓄積していることも示されている10,11).ただし,この蓄積はXist RNAに依存すること,ES細胞が分化してから観察されるようになることなどから,筆者らが提唱するモデルとは趣が異なるように思われる.hnRNP Uは核酸の存在により多量体化することが知られており,別グループらによって観察されたX染色体へのhnRNP Uの蓄積は,Xist RNAが初期量の(それほど多くはない)hnRNP Uによって繋留された“結果”としてみられる現象なのかもしれない.
おわりに
最後に,“hnRNP U is required for chromosomal localization of Xist RNA”というこの論文のタイトルをひねくれた角度からみてみようと思う.このタイトルは,X染色体を“動かないもの”,Xist RNAを“動くもの”と仮定したときに成立する.かたや,Edithらは,2006年に発表した論文“A novel role for Xist RNA in the formation of a repressive nuclear compartment into which gene are recruited when silenced”において,Xist RNAが形成する“nuclear compartment”に発現抑制されるべき遺伝子が引き込まれる,と主張した12).“動くもの”と位置づけている対象が,筆者らと逆なのである.Edithらの主張に筆者らの今回の研究をすり合わせてみると,hnRNP Uのない状況で阻害されたのはこの“nuclear compartment”の形成である,といういい方ができる.実験結果をふまえると,hnRNP UのDNA結合能(すなわち,おそらくX染色体との相互作用)もXist RNAの正常な局在化に必要であるので,Xist RNAがX染色体の位置と無関係に勝手な場所に“nuclear compartment”を形成しているわけではなく,相互に動向を制御しあっているのであろう.不活性化したX染色体とは巨大なRNPなのであろうか.
近年,Xist RNA様の機能をもつ複数のノンコーディングRNAがみつかってきており,ノンコーディングRNAとエピジェネティクスとの関連が注目されている.この研究がこの分野の発展および理解に貢献する道標のひとつとなることを期待する.
文 献
- Borsani, G., Tonlorenzi, R., Simmler, M. C. et al.: Characterization of a murine gene expressed from the inactive X chromosome. Nature, 351, 325-329 (1991)[PubMed]
- Brockdorff, N., Ashworth, A., Kay, G. F. et al.: Conservation of position and exclusive expression of mouse Xist from the inactive X chromosome. Nature, 351, 329-331 (1991)[PubMed]
- Penny, G. D., Kay, G. F., Sheardown, S. A. et al.: Requirement for Xist in X chromosome inactivation. Nature, 379, 131-137 (1996)[PubMed]
- Silva, J., Mak, W., Zvetkova, I. et al.: Establishment of histone h3 methylation on the inactive X chromosome requires transient recruitment of Eed-Enx1 polycomb group complexes. Dev. Cell, 4, 481-495 (2003)[PubMed]
- Wutz, A., Rasmussen, T. P. & Jaenisch, R.: Chromosomal silencing and localization are mediated by different domains of Xist RNA. Nat. Genet., 30, 167-174 (2002)[PubMed]
- Kiledjian, M. & Dreyfuss, G.: Primary structure and binding activity of the hnRNP U protein: binding RNA through RGG box. EMBO J., 11, 2655-2664 (1992)[PubMed]
- Romig, H., Fackelmayer, F. O., Renz, A. et al.: Characterization of SAF-A, a novel nuclear DNA binding protein from HeLa cells with high affinity for nuclear matrix/scaffold attachment DNA elements. EMBO J., 11, 3431-3440 (1992)[PubMed]
- Tsutsui, K., Okada, S., Watarai, S. et al.: Identification and characterization of a nuclear scaffold protein that binds the matrix attachment region DNA. J. Biol. Chem., 268, 12886-12894 (1993)[PubMed]
- Csankovszki, G., Panning, B., Bates, B. et al.: Conditional deletion of Xist disrupts histone macroH2A localization but not maintenance of X inactivation. Nat. Genet., 22, 323-324 (1999)[PubMed]
- Fackelmayer, F. O.: A stable proteinaceous structure in the territory of inactive X chromosomes. J. Biol. Chem., 280, 1720-1723 (2005)[PubMed]
- Pullirsch, D., Hartel, R., Kishimoto, H. et al.: The Trithorax group protein Ash2l and Saf-A are recruited to the inactive X chromosome at the onset of stable X inactivation. Development, 137, 935-943 (2010)[PubMed]
- Chaumeil, J., Le Baccon, P., Wutz, A. et al.: A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes Dev., 20, 2223-2237 (2006)[PubMed]
著者プロフィール
略歴:東京大学大学院工学系研究科博士課程 在学中.
研究テーマ:核内ノンコーディングRNAの制御機構.
抱負:冷静に熱い仕事をする.
中川 真一(Shinichi Nakagawa)
理化学研究所基幹研究所 准主任研究員.
研究室URL:http://www.riken.jp/lab-www/nakagawa/index_j.html
© 2010 長谷川優子・中川真一 Licensed under CC 表示 2.1 日本
