アブラナ科植物の自家不和合性におけるトランス作用性のsmall RNAを介した優劣性発現の分子機構
樽谷芳明・高山誠司
(奈良先端科学技術大学院大学バイオサイエンス研究科 細胞生物学専攻細胞間情報学講座)
email:樽谷芳明,高山誠司
DOI: 10.7875/first.author.2010.031
Trans-acting small RNA determines dominance relationships in Brassica self-incompatibility.
Yoshiaki Tarutani, Hiroshi Shiba, Megumi Iwano, Tomohiro Kakizaki, Go Suzuki, Masao Watanabe, Akira Isogai, Seiji Takayama
Nature, 466, 983-986 (2010)
有性生殖で生まれる子どもは両親から1組ずつの遺伝子を受け継ぐ.受け継いだ2組の対立遺伝子のうち,“メンデルの優性の法則”に従っていずれか一方の形質のみが現れる場合が数多く知られている.しかしながら,なぜ一方の対立遺伝子の形質のみが現れるのか,そのしくみに関しては未解明の部分が多い.このしくみのひとつにせまるため,筆者らは,アブラナ科植物の自家不和合性において花粉側S因子をコードするSP11遺伝子の優劣性に着目した.この遺伝子は多数の対立遺伝子をもつが,筆者らはこれまで,一部のヘテロ株において片方の対立遺伝子の発現が完全に抑制されること,このとき,劣性側対立遺伝子のプロモーター領域が葯タペート組織において特異的にメチル化されることを見い出してきた.今回,劣性SP11対立遺伝子のプロモーター領域の特異的なメチル化が,優性SP11対立遺伝子の近傍のゲノム領域からつくられるsmall RNAによって制御されていることを明らかにした.
子どもは両親から1組ずつの遺伝子を受け継ぐが,いずれか一方の形質のみを示す場合が多く知られており,この現象は“メンデルの優性の法則”として古くから知られている.そのしくみについては,優性側の対立遺伝子が機能的なタンパク質をコードするのに対し劣性側の対立遺伝子は機能を欠失しているためである,と基本的には理解されてきた.しかし,筆者らは,アブラナ科植物における自家不和合性という現象を解析する過程で,いずれの対立遺伝子も機能的なタンパク質をコードしているにもかかわらず,優性側の対立遺伝子の影響で劣性側の対立遺伝子の機能が消失してしまう現象を見い出してきた.詳細な解析の結果,優性側対立遺伝子の近傍の逆位反復配列部位からつくられる24塩基のsmall RNAにより,劣性側対立遺伝子のプロモーター領域がメチル化され,その結果,劣性側対立遺伝子の発現が抑制されていることが明らかになった.優劣性という古典的な遺伝学の現象にエピジェネティックな遺伝子発現制御機構が関与していることを示すはじめての例として,この研究成果について従来の研究経緯を含め紹介する.
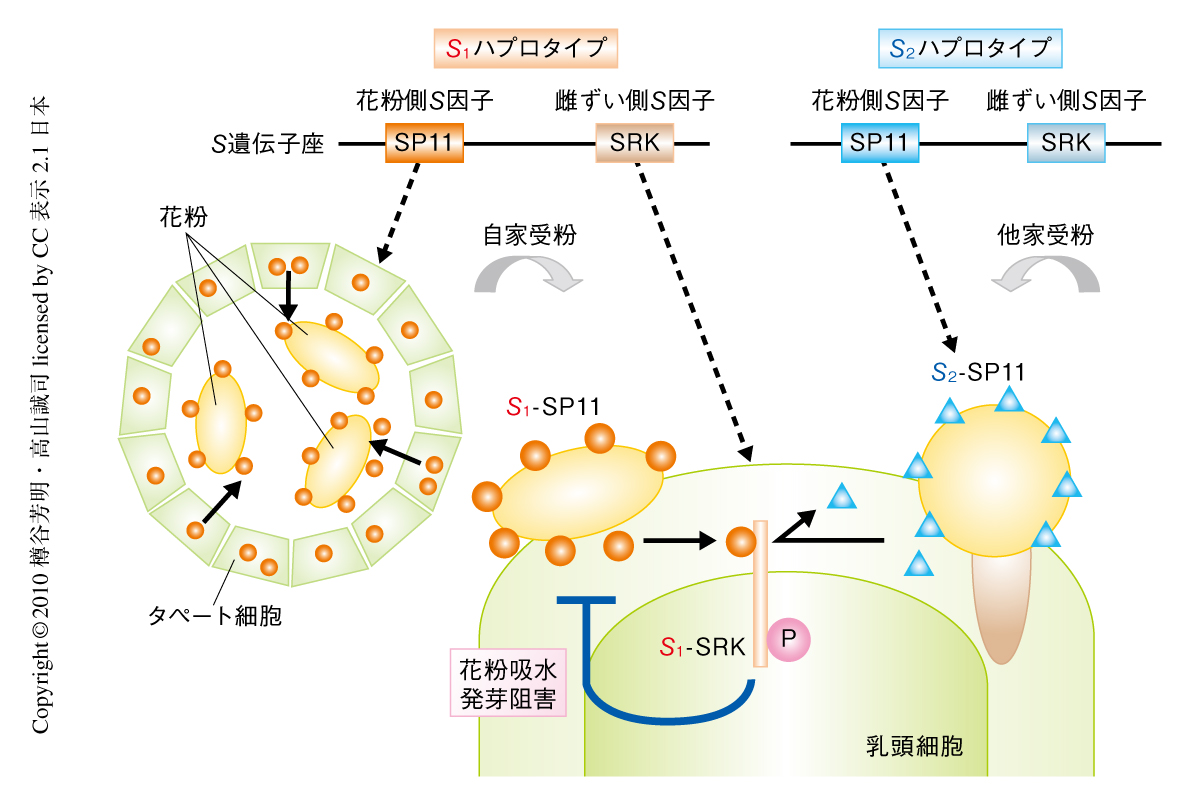
多くの植物は受粉から受精にいたる有性生殖の過程において,自己と非自己の花粉を識別し自殖を抑制する自家不和合性とよばれる性質をもつ1).自己と非自己の識別はS遺伝子座上にコードされる花粉側S因子と雌ずい側S因子とによって制御されている.両方のS因子は多数の複対立遺伝子によりコードされており多型性を示すが,花粉側のS因子遺伝子と雌ずい側のS因子遺伝子とは密接に連鎖しており組換えが起こらないので,両方の遺伝子のセットをSハプロタイプ(S1,S2,…,Sn)とよんでいる.基本的に,花粉と雌ずいとが同一のSハプロタイプに由来する花粉S因子と雌ずいS因子とを発現しているとき,その受粉は不和合となる.アブラナ科植物では,花粉側S因子としてSP11/SCRというシステイン残基に富む低分子量のタンパク質が2-4),雌ずい側S因子としてSRKという受容体型キナーゼが5),それぞれ同定されている.そして,受粉の際,同一のSハプロタイプに由来するSP11リガンドとSRK受容体が雌ずい先端の乳頭細胞で出会うと(自家受粉),SRKを活性化(自己リン酸化)し,不和合反応を誘起することが示されている6,7)(図1,左).一方,SP11とSRKが異なるSハプロタイプに由来する場合は(他家受粉),不和合反応は誘起されず,花粉の発芽および伸長が起こり受精にいたる(図1,右).
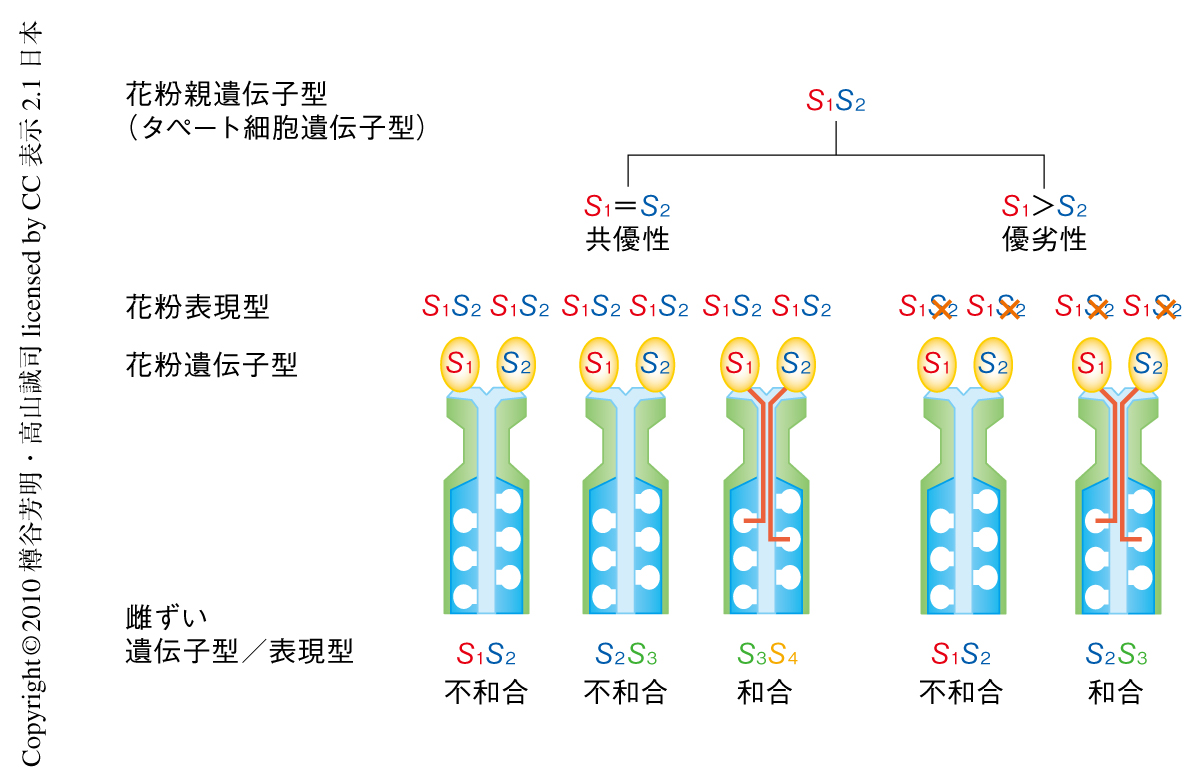
花粉側S因子であるSP11は配偶体(n)の花粉ではなく胞子体(2n)の葯のタペート組織でつくられ,花粉の成熟にともなって花粉表層に移行する4).したがって,2種類のSハプロタイプをもつヘテロ株の花粉は,花粉親の2種類のSハプロタイプの形質をあわせもつことになる.しかし,Sハプロタイプの組合せによっては片方のSハプロタイプの形質が消失してしまう,つまり,Sハプロタイプのあいだで優劣の関係が生じる場合があることが古くから示されてきた8).たとえば,S1S2ヘテロ体の花粉は,通常(S1とS2とが共優性)の場合,S1S2という表現型を示し,S1あるいはS2をもつS1S2やS2S3の雌ずいに対して不和合,まったく異なる組合せとなるS3S4の雌ずいに対して和合となる(図2,左).一方,S1がS2に対して優性の場合には花粉は優性であるS1の表現型のみを示し,S1S2の雌ずいに対して不和合となるが,S2S3の雌ずいに対しては和合となる(図2,右).実際に,アブラナ科植物Brassica rapaにおいて多数のSハプロタイプのあいだの優劣性の関係が調べられた結果,Sハプロタイプは,一般的に花粉優性を示すクラスI(S8,S9,S12,S52など)と,それらとの組合せでは花粉劣性となるクラスII(S29,S40,S44,S60など)の2つに大きく分類されることが示された8-10).また,クラスIのSハプロタイプのあいだでは共優性を示す場合の多いこと,クラスIIのSハプロタイプのあいだには直線的な優劣性の関係(S44 > S60 > S40 > S29)の存在することも明らかになってきた.
花粉におけるSハプロタイプのあいだの優劣性の原因を明らかにするため,優劣性を生じる複数のSヘテロ株を作出し,SP11対立遺伝子の発現を調べた.in situハイブリダイゼーションによる解析の結果,劣性SP11対立遺伝子は劣性ホモ株では葯タペート組織で発現しているが,優性/劣性ヘテロ株ではその発現はホモ株の数万分の1にまで抑制されていた9).一方,優性SP11対立遺伝子はヘテロ株でもタペート組織で発現していた.さらに,直線的な優劣性の関係が存在するクラスIIどうしの組合せとなるヘテロ株でも,すべての組合せにおいて相対的に劣性となるSP11対立遺伝子の発現が抑制されていた10).
つぎに,優性/劣性ヘテロ株でみられる劣性SP11対立遺伝子に特異的な発現抑制がどのような機構で起こるのかを検証した.植物におけるDNAメチル化は,ヒストン修飾とならんでエピジェネティックな遺伝子発現制御機構のおもな原因である.そこで,劣性SP11対立遺伝子の発現抑制にDNAメチル化が関与しているかどうかを検証した.SP11対立遺伝子の発現部位である葯タペート組織を効率よく回収する方法を考案し,優性/劣性ヘテロ株と劣性ホモ株の葯タペート組織におけるSP11対立遺伝子のDNAメチル化パターンをバイサルファイトシークエンス法により調べた.その結果,優性/劣性ヘテロ株における劣性SP11対立遺伝子のプロモーター領域が特異的にde novo DNAメチル化されていた11).このde novo DNAメチル化は,CG配列,CHG配列,CHH配列(H:A,C,T)のすべてでみられ,タペート組織以外では認められないこと,SP11対立遺伝子の発現がはじまる直前に起こっていることが明らかとなった11).
この現象には,そのメチル化パターンからsmall RNAを介したメチル化機構(RNA-directed DNA methylation)が関与しているものと考えられた.そこで,優性SハプロタイプであるクラスIに属するS9とS12のS遺伝子座を調べたところ,劣性SP11対立遺伝子のメチル化領域と相同な配列を含む逆位反復配列がみつかった.この配列は,PCR法によりクラスIに属するS8とS52からも見い出された.RT-PCR法の結果,この領域はタペート組織が存在する時期の葯で発現しており,雌ずいや葉での発現は認められなかった.
つぎに,S9S9ホモ株の葯を材料とし,劣性SP11対立遺伝子のメチル化領域と相同なsmall RNAのスクリーニングを行ったところ,さきに見い出した領域から産出された24塩基のsmall RNA(SP11 methylation inducer:Smi)を得た.Smiは劣性SP11対立遺伝子メチル化領域と19塩基中18塩基の一致という高い相同性を示した.in situハイブリダイゼーションによる解析の結果,Smiは葯タペート組織に特異的に,劣性SP11対立遺伝子のメチル化がはじまるまえから存在していることが明らかとなった.
予想に反して,Smiは劣性SハプロタイプであるS60,S40,S29からもみつかった.劣性Smiは,優性Smiと比較して,10番目の塩基がUからAへと置換されており,その結果,劣性SP11対立遺伝子のメチル化領域への相同性が19塩基中17塩基の一致に下がっていた.また,その存在量も優性Smiと比べ非常に少なかった.これらの結果から,存在量が多く相同性も高い優性Smiが劣性SP11対立遺伝子のメチル化をひき起こし発現を抑制しているのではないかと考えられた.
実際に,優性Smiが劣性SP11対立遺伝子のメチル化をひき起こせるかどうかを検証するため,形質転換アブラナを作出した.優性Smi(S9-Smi)のゲノム領域を導入した劣性Sハプロタイプホモ株はすべて自家和合となっており,その原因は花粉側の自家不和合性の表現型の消失であった.この優性Smiゲノム領域を導入した劣性Sハプロタイプホモ株における劣性SP11対立遺伝子の発現およびメチル化状態を調べたところ,優性/劣性ヘテロ株と同様にその発現は著しく抑制されており,プロモーター領域の顕著なメチル化が認められた.以上の結果から,優性/劣性ヘテロ株において,優性Smiが劣性SP11対立遺伝子プロモーター領域特異的なde novo DNAメチル化をひき起こし,劣性SP11対立遺伝子の発現が抑制されることが示された.
劣性Smiが劣性SP11対立遺伝子の発現を抑制できない原因を探るため,優性Smiゲノム領域に劣性Smiを産出するよう1塩基置換を導入(S9-SmiT10A)したゲノム領域を用い,形質転換アブラナを作出した.この1塩基置換優性Smiゲノム領域導入S60S60ホモ株は,劣性Smiを優性Smiと同じ程度まで発現したにもかかわらず花粉における自家不和合性の表現型を消失していなかった.また,SP11対立遺伝子であるS60の発現レベルやメチル化状態もS60S60ホモ株と同様であった.これらの結果から,劣性SP11対立遺伝子のプロモーター領域に対して,優性Smiは10番目の塩基が一致しているが劣性Smiは一致していないため,劣性Smiは劣性SP11対立遺伝子の発現を抑制できないものと考えられた.
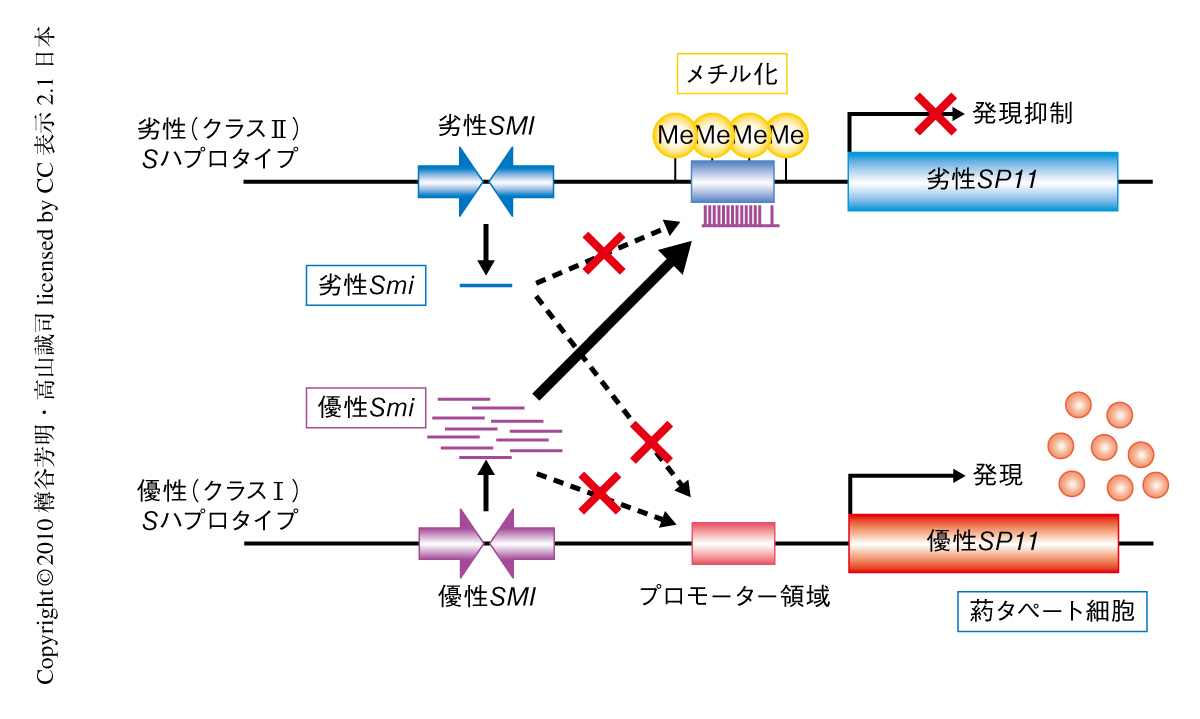
アブラナ科植物の花粉における優劣性の発現機構に関するモデルを示す(図3).優劣の関係にあるSハプロタイプをもつヘテロ株のタペート細胞においては,優性Smiがトランスに作用し,劣性SP11対立遺伝子のプロモーター領域に特異的なde novo DNAメチル化をひき起こし,劣性SP11対立遺伝子の発現が抑制される.一方,優性SP11対立遺伝子のプロモーター領域のメチル化は起こらず,優性SP11対立遺伝子は発現する.その結果,優性/劣性ヘテロ株のタペート細胞では優性SP11タンパク質のみが生産され,花粉は優性Sハプロタイプの形質のみを示す.
この研究により,クラスIとクラスIIのSハプロタイプのあいだに認められる優劣性の関係がSmiというsmall RNAを介してエピジェネティックに制御されていることが明らかとなった.しかし,前述のように,クラスIとクラスIIのSハプロタイプのあいだにある優劣性だけでなく,クラスIIのSハプロタイプのあいだにも直線的な優劣性が存在する.このクラスIIのSハプロタイプのあいだにある優劣性についてはSmiで説明することはできず,分子機構は未解明のまま残されている.また,同様な優劣性の関係は,今回,解析したB. rapaのみならず,ほかのアブラナ科植物やヒルガオ科植物などほかの胞子体型の自家不和合性植物において広く認められる.これらの優劣性現象にも類似の機構が関与しているのかどうか,今後,明らかにしていく必要がある.
この研究では,アブラナ科植物の自家不和合性における花粉の優劣性において,優性SP11対立遺伝子の近傍の領域に由来するsmall RNAがトランスに作用し,時期特異的および組織特異的な劣性SP11対立遺伝子のde novo DNAメチル化をひき起こして,その結果,劣性SP11対立遺伝子の発現が抑制されることを明らかにした.今回の発見は,動植物を問わず広く存在する単一対立遺伝子性の遺伝子発現制御系に新たなモデルを提唱するものであり,制御機構の解明に新たな展開をもたらすものと期待される.
略歴:2004年 東京大学大学院農学生命科学研究科博士課程 修了,同年 同 農学研究員,同年 奈良先端科学技術大学院大学バイオサイエンス研究科 ポスドクを経て,2010年より国立遺伝学研究所 助教.
研究テーマ:シロイヌナズナにおけるエピジェネティックな遺伝子発現制御機構の解明.
抱負:遊び心を忘れずに,植物のエピジェネティックな遺伝子発現制御機構の解明に取り組んでいきたい.
高山 誠司(Seiji Takayama)
略歴:1986年 東京大学大学院農学研究科博士課程 修了(農学博士),1995年 奈良先端科学技術大学院大学バイオサイエンス研究科 助教授を経て,2006年より同 教授.
研究テーマ:植物の細胞間情報伝達.
研究室URL:http://bsw3.naist.jp/takayama/index.html
© 2010 樽谷芳明・高山誠司 Licensed under CC 表示 2.1 日本
(奈良先端科学技術大学院大学バイオサイエンス研究科 細胞生物学専攻細胞間情報学講座)
email:樽谷芳明,高山誠司
DOI: 10.7875/first.author.2010.031
Trans-acting small RNA determines dominance relationships in Brassica self-incompatibility.
Yoshiaki Tarutani, Hiroshi Shiba, Megumi Iwano, Tomohiro Kakizaki, Go Suzuki, Masao Watanabe, Akira Isogai, Seiji Takayama
Nature, 466, 983-986 (2010)
要 約
有性生殖で生まれる子どもは両親から1組ずつの遺伝子を受け継ぐ.受け継いだ2組の対立遺伝子のうち,“メンデルの優性の法則”に従っていずれか一方の形質のみが現れる場合が数多く知られている.しかしながら,なぜ一方の対立遺伝子の形質のみが現れるのか,そのしくみに関しては未解明の部分が多い.このしくみのひとつにせまるため,筆者らは,アブラナ科植物の自家不和合性において花粉側S因子をコードするSP11遺伝子の優劣性に着目した.この遺伝子は多数の対立遺伝子をもつが,筆者らはこれまで,一部のヘテロ株において片方の対立遺伝子の発現が完全に抑制されること,このとき,劣性側対立遺伝子のプロモーター領域が葯タペート組織において特異的にメチル化されることを見い出してきた.今回,劣性SP11対立遺伝子のプロモーター領域の特異的なメチル化が,優性SP11対立遺伝子の近傍のゲノム領域からつくられるsmall RNAによって制御されていることを明らかにした.
はじめに
子どもは両親から1組ずつの遺伝子を受け継ぐが,いずれか一方の形質のみを示す場合が多く知られており,この現象は“メンデルの優性の法則”として古くから知られている.そのしくみについては,優性側の対立遺伝子が機能的なタンパク質をコードするのに対し劣性側の対立遺伝子は機能を欠失しているためである,と基本的には理解されてきた.しかし,筆者らは,アブラナ科植物における自家不和合性という現象を解析する過程で,いずれの対立遺伝子も機能的なタンパク質をコードしているにもかかわらず,優性側の対立遺伝子の影響で劣性側の対立遺伝子の機能が消失してしまう現象を見い出してきた.詳細な解析の結果,優性側対立遺伝子の近傍の逆位反復配列部位からつくられる24塩基のsmall RNAにより,劣性側対立遺伝子のプロモーター領域がメチル化され,その結果,劣性側対立遺伝子の発現が抑制されていることが明らかになった.優劣性という古典的な遺伝学の現象にエピジェネティックな遺伝子発現制御機構が関与していることを示すはじめての例として,この研究成果について従来の研究経緯を含め紹介する.
1.自家不和合性
多くの植物は受粉から受精にいたる有性生殖の過程において,自己と非自己の花粉を識別し自殖を抑制する自家不和合性とよばれる性質をもつ1).自己と非自己の識別はS遺伝子座上にコードされる花粉側S因子と雌ずい側S因子とによって制御されている.両方のS因子は多数の複対立遺伝子によりコードされており多型性を示すが,花粉側のS因子遺伝子と雌ずい側のS因子遺伝子とは密接に連鎖しており組換えが起こらないので,両方の遺伝子のセットをSハプロタイプ(S1,S2,…,Sn)とよんでいる.基本的に,花粉と雌ずいとが同一のSハプロタイプに由来する花粉S因子と雌ずいS因子とを発現しているとき,その受粉は不和合となる.アブラナ科植物では,花粉側S因子としてSP11/SCRというシステイン残基に富む低分子量のタンパク質が2-4),雌ずい側S因子としてSRKという受容体型キナーゼが5),それぞれ同定されている.そして,受粉の際,同一のSハプロタイプに由来するSP11リガンドとSRK受容体が雌ずい先端の乳頭細胞で出会うと(自家受粉),SRKを活性化(自己リン酸化)し,不和合反応を誘起することが示されている6,7)(図1,左).一方,SP11とSRKが異なるSハプロタイプに由来する場合は(他家受粉),不和合反応は誘起されず,花粉の発芽および伸長が起こり受精にいたる(図1,右).
2.アブラナ科植物の花粉側S因子の表現型における優劣性
花粉側S因子であるSP11は配偶体(n)の花粉ではなく胞子体(2n)の葯のタペート組織でつくられ,花粉の成熟にともなって花粉表層に移行する4).したがって,2種類のSハプロタイプをもつヘテロ株の花粉は,花粉親の2種類のSハプロタイプの形質をあわせもつことになる.しかし,Sハプロタイプの組合せによっては片方のSハプロタイプの形質が消失してしまう,つまり,Sハプロタイプのあいだで優劣の関係が生じる場合があることが古くから示されてきた8).たとえば,S1S2ヘテロ体の花粉は,通常(S1とS2とが共優性)の場合,S1S2という表現型を示し,S1あるいはS2をもつS1S2やS2S3の雌ずいに対して不和合,まったく異なる組合せとなるS3S4の雌ずいに対して和合となる(図2,左).一方,S1がS2に対して優性の場合には花粉は優性であるS1の表現型のみを示し,S1S2の雌ずいに対して不和合となるが,S2S3の雌ずいに対しては和合となる(図2,右).実際に,アブラナ科植物Brassica rapaにおいて多数のSハプロタイプのあいだの優劣性の関係が調べられた結果,Sハプロタイプは,一般的に花粉優性を示すクラスI(S8,S9,S12,S52など)と,それらとの組合せでは花粉劣性となるクラスII(S29,S40,S44,S60など)の2つに大きく分類されることが示された8-10).また,クラスIのSハプロタイプのあいだでは共優性を示す場合の多いこと,クラスIIのSハプロタイプのあいだには直線的な優劣性の関係(S44 > S60 > S40 > S29)の存在することも明らかになってきた.
3.劣性SP11対立遺伝子に特異的な発現抑制
花粉におけるSハプロタイプのあいだの優劣性の原因を明らかにするため,優劣性を生じる複数のSヘテロ株を作出し,SP11対立遺伝子の発現を調べた.in situハイブリダイゼーションによる解析の結果,劣性SP11対立遺伝子は劣性ホモ株では葯タペート組織で発現しているが,優性/劣性ヘテロ株ではその発現はホモ株の数万分の1にまで抑制されていた9).一方,優性SP11対立遺伝子はヘテロ株でもタペート組織で発現していた.さらに,直線的な優劣性の関係が存在するクラスIIどうしの組合せとなるヘテロ株でも,すべての組合せにおいて相対的に劣性となるSP11対立遺伝子の発現が抑制されていた10).
4.劣性SP11対立遺伝子のプロモーター領域に特異的なDNAメチル化
つぎに,優性/劣性ヘテロ株でみられる劣性SP11対立遺伝子に特異的な発現抑制がどのような機構で起こるのかを検証した.植物におけるDNAメチル化は,ヒストン修飾とならんでエピジェネティックな遺伝子発現制御機構のおもな原因である.そこで,劣性SP11対立遺伝子の発現抑制にDNAメチル化が関与しているかどうかを検証した.SP11対立遺伝子の発現部位である葯タペート組織を効率よく回収する方法を考案し,優性/劣性ヘテロ株と劣性ホモ株の葯タペート組織におけるSP11対立遺伝子のDNAメチル化パターンをバイサルファイトシークエンス法により調べた.その結果,優性/劣性ヘテロ株における劣性SP11対立遺伝子のプロモーター領域が特異的にde novo DNAメチル化されていた11).このde novo DNAメチル化は,CG配列,CHG配列,CHH配列(H:A,C,T)のすべてでみられ,タペート組織以外では認められないこと,SP11対立遺伝子の発現がはじまる直前に起こっていることが明らかとなった11).
5.優性SP11対立遺伝子の近傍に由来するsmall RNAによる劣性SP11対立遺伝子の発現抑制
この現象には,そのメチル化パターンからsmall RNAを介したメチル化機構(RNA-directed DNA methylation)が関与しているものと考えられた.そこで,優性SハプロタイプであるクラスIに属するS9とS12のS遺伝子座を調べたところ,劣性SP11対立遺伝子のメチル化領域と相同な配列を含む逆位反復配列がみつかった.この配列は,PCR法によりクラスIに属するS8とS52からも見い出された.RT-PCR法の結果,この領域はタペート組織が存在する時期の葯で発現しており,雌ずいや葉での発現は認められなかった.
つぎに,S9S9ホモ株の葯を材料とし,劣性SP11対立遺伝子のメチル化領域と相同なsmall RNAのスクリーニングを行ったところ,さきに見い出した領域から産出された24塩基のsmall RNA(SP11 methylation inducer:Smi)を得た.Smiは劣性SP11対立遺伝子メチル化領域と19塩基中18塩基の一致という高い相同性を示した.in situハイブリダイゼーションによる解析の結果,Smiは葯タペート組織に特異的に,劣性SP11対立遺伝子のメチル化がはじまるまえから存在していることが明らかとなった.
予想に反して,Smiは劣性SハプロタイプであるS60,S40,S29からもみつかった.劣性Smiは,優性Smiと比較して,10番目の塩基がUからAへと置換されており,その結果,劣性SP11対立遺伝子のメチル化領域への相同性が19塩基中17塩基の一致に下がっていた.また,その存在量も優性Smiと比べ非常に少なかった.これらの結果から,存在量が多く相同性も高い優性Smiが劣性SP11対立遺伝子のメチル化をひき起こし発現を抑制しているのではないかと考えられた.
実際に,優性Smiが劣性SP11対立遺伝子のメチル化をひき起こせるかどうかを検証するため,形質転換アブラナを作出した.優性Smi(S9-Smi)のゲノム領域を導入した劣性Sハプロタイプホモ株はすべて自家和合となっており,その原因は花粉側の自家不和合性の表現型の消失であった.この優性Smiゲノム領域を導入した劣性Sハプロタイプホモ株における劣性SP11対立遺伝子の発現およびメチル化状態を調べたところ,優性/劣性ヘテロ株と同様にその発現は著しく抑制されており,プロモーター領域の顕著なメチル化が認められた.以上の結果から,優性/劣性ヘテロ株において,優性Smiが劣性SP11対立遺伝子プロモーター領域特異的なde novo DNAメチル化をひき起こし,劣性SP11対立遺伝子の発現が抑制されることが示された.
劣性Smiが劣性SP11対立遺伝子の発現を抑制できない原因を探るため,優性Smiゲノム領域に劣性Smiを産出するよう1塩基置換を導入(S9-SmiT10A)したゲノム領域を用い,形質転換アブラナを作出した.この1塩基置換優性Smiゲノム領域導入S60S60ホモ株は,劣性Smiを優性Smiと同じ程度まで発現したにもかかわらず花粉における自家不和合性の表現型を消失していなかった.また,SP11対立遺伝子であるS60の発現レベルやメチル化状態もS60S60ホモ株と同様であった.これらの結果から,劣性SP11対立遺伝子のプロモーター領域に対して,優性Smiは10番目の塩基が一致しているが劣性Smiは一致していないため,劣性Smiは劣性SP11対立遺伝子の発現を抑制できないものと考えられた.
アブラナ科植物の花粉における優劣性の発現機構に関するモデルを示す(図3).優劣の関係にあるSハプロタイプをもつヘテロ株のタペート細胞においては,優性Smiがトランスに作用し,劣性SP11対立遺伝子のプロモーター領域に特異的なde novo DNAメチル化をひき起こし,劣性SP11対立遺伝子の発現が抑制される.一方,優性SP11対立遺伝子のプロモーター領域のメチル化は起こらず,優性SP11対立遺伝子は発現する.その結果,優性/劣性ヘテロ株のタペート細胞では優性SP11タンパク質のみが生産され,花粉は優性Sハプロタイプの形質のみを示す.
おわりに
この研究により,クラスIとクラスIIのSハプロタイプのあいだに認められる優劣性の関係がSmiというsmall RNAを介してエピジェネティックに制御されていることが明らかとなった.しかし,前述のように,クラスIとクラスIIのSハプロタイプのあいだにある優劣性だけでなく,クラスIIのSハプロタイプのあいだにも直線的な優劣性が存在する.このクラスIIのSハプロタイプのあいだにある優劣性についてはSmiで説明することはできず,分子機構は未解明のまま残されている.また,同様な優劣性の関係は,今回,解析したB. rapaのみならず,ほかのアブラナ科植物やヒルガオ科植物などほかの胞子体型の自家不和合性植物において広く認められる.これらの優劣性現象にも類似の機構が関与しているのかどうか,今後,明らかにしていく必要がある.
この研究では,アブラナ科植物の自家不和合性における花粉の優劣性において,優性SP11対立遺伝子の近傍の領域に由来するsmall RNAがトランスに作用し,時期特異的および組織特異的な劣性SP11対立遺伝子のde novo DNAメチル化をひき起こして,その結果,劣性SP11対立遺伝子の発現が抑制されることを明らかにした.今回の発見は,動植物を問わず広く存在する単一対立遺伝子性の遺伝子発現制御系に新たなモデルを提唱するものであり,制御機構の解明に新たな展開をもたらすものと期待される.
文 献
- de Nettancourt, D.: Incompatibility in Angiosperms. Springer-Verlag, Berlin (1977)
- Schopfer, C. R., Nasrallah, M. E. & Nasrallah, J. B.: The male determinant of self-incompatibility in Brassica. Science, 286, 1697-1700 (1999)[PubMed]
- Takayama, S., Shiba, H., Iwano, M. et al.: The pollen determinant of self-incompatibility in Brassica campestris. Proc. Natl. Acad. Sci. USA, 97, 1920-1925 (2000)[PubMed]
- Shiba, H., Takayama, S., Iwano, M. et al.: A pollen coat protein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiol., 125, 2095-2103 (2001)[PubMed]
- Takasaki, T., Hatakeyama, K., Suzuki, G. et al.: The S receptor kinase determines self-incompatibility in Brassica stigma. Nature, 403, 913-916 (2000)[PubMed]
- Kachroo, A., Schopfer, C. R., Nasrallah, M. E. et al.: Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science, 293, 1824-1826 (2001)[PubMed]
- Takayama, S., Shimosato, H., Shiba, H. et al.: Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature, 413, 534-538 (2001)[PubMed]
- Hatakeyama, K., Watanabe, M., Takasaki, T. et al.: Dominance relationships between S-alleles in self-incompatible Brassica campestris L. Heredity, 80, 241-247 (1998)
- Shiba, H., Iwano, M., Entani, T. et al.: The dominance of alleles controlling self-incompatibility in Brassica pollen is regulated at the RNA level. Plant Cell, 14, 491-504 (2002)[PubMed]
- Kakizaki, T., Takada, Y., Ito, A. et al.: Linear dominance relationship among four class-II S haplotypes in pollen is determined by the expression of SP11 in Brassica self-incompatibility. Plant Cell Physiol., 44, 70-75 (2003)[PubMed]
- Shiba, H., Kakizaki, T., Iwano, M. et al.: Dominance relationships between self-incompatibility alleles controlled by DNA methylation. Nat. Genet., 38, 297-299 (2006)[PubMed]
著者プロフィール
略歴:2004年 東京大学大学院農学生命科学研究科博士課程 修了,同年 同 農学研究員,同年 奈良先端科学技術大学院大学バイオサイエンス研究科 ポスドクを経て,2010年より国立遺伝学研究所 助教.
研究テーマ:シロイヌナズナにおけるエピジェネティックな遺伝子発現制御機構の解明.
抱負:遊び心を忘れずに,植物のエピジェネティックな遺伝子発現制御機構の解明に取り組んでいきたい.
高山 誠司(Seiji Takayama)
略歴:1986年 東京大学大学院農学研究科博士課程 修了(農学博士),1995年 奈良先端科学技術大学院大学バイオサイエンス研究科 助教授を経て,2006年より同 教授.
研究テーマ:植物の細胞間情報伝達.
研究室URL:http://bsw3.naist.jp/takayama/index.html
© 2010 樽谷芳明・高山誠司 Licensed under CC 表示 2.1 日本
