大脳の視覚野における情報処理形態の学習による変化
牧野浩史・小宮山尚樹
(米国California大学San Diego校Department of Neurosciences)
email:牧野浩史,小宮山尚樹
DOI: 10.7875/first.author.2015.092
Learning enhances the relative impact of top-down processing in the visual cortex.
Hiroshi Makino, Takaki Komiyama
Nature Neuroscience, 18, 1116-1122 (2015)
学習とは,外界に対し脳において内部予測モデルを構築し,高次の脳領域からの予測や期待あるいは注意などの情報をつかさどるトップダウン入力を増大させるとともに,外界から得られる情報を処理するボトムアップ入力を減少させるものと考えられている.筆者らは,この理論を検証するため,視覚情報による連合学習中の1次視覚野の2/3層の興奮性ニューロン,そのボトムアップ入力のもとである4層の興奮性ニューロン,トップダウン入力である脳梁膨大後部皮質から直接に投射する興奮性の軸索の活動を2光子Ca2+イメージング法を用いて評価した.これにより,4層の活動はしだいに低下するのに対し,脳梁膨大後部皮質からの軸索の活動は上昇し2/3層への影響を強めることが明らかにされた.それとともに,抑制性ニューロンでありトップダウン入力を制御すると考えられているソマトスタチン陽性細胞の活動も学習をつうじ低下することがわかった.これらの結果から,学習がボトムアップ入力とトップダウン入力の情報処理機構のバランスに動的なシフトを促すとともに,その微小回路における神経機構も明らかにされた.
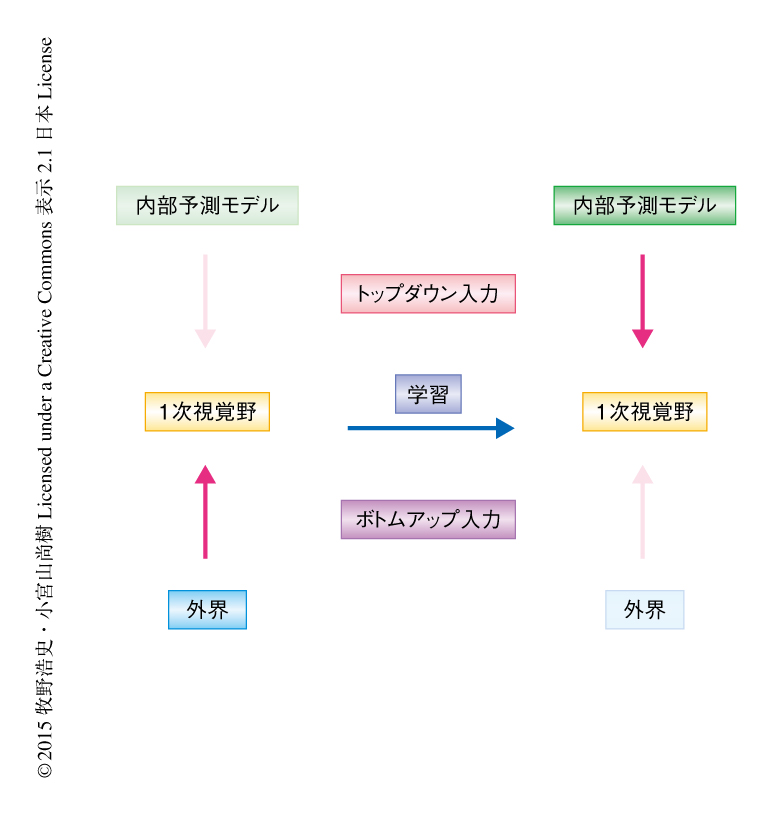
予測コーディング理論によると,脳は外界からの感覚入力をうまく説明できるような内部予測モデルを構築し,未来に起こりうる現象に対し適切な対応ができるようにすると考えられている1,2).この理論によれば,脳は階層構造をとっており,学習により得られた内部予測モデルにしたがい上の層から下の層へと予測信号(トップダウン入力)を伝達する.もし,この予測信号が実際の感覚入力と違う場合,下の層から上の層へと予測エラー信号(ボトムアップ入力)が伝達され,内部予測モデルは更新される.学習とはまさにこのような階層のあいだの相互作用により脳において内部予測モデルを徐々に最適化させる過程のことであり,脳は予測信号の正確性を最大化し,予測エラー信号を最小化させることを目的にしていると考えられる(図1).
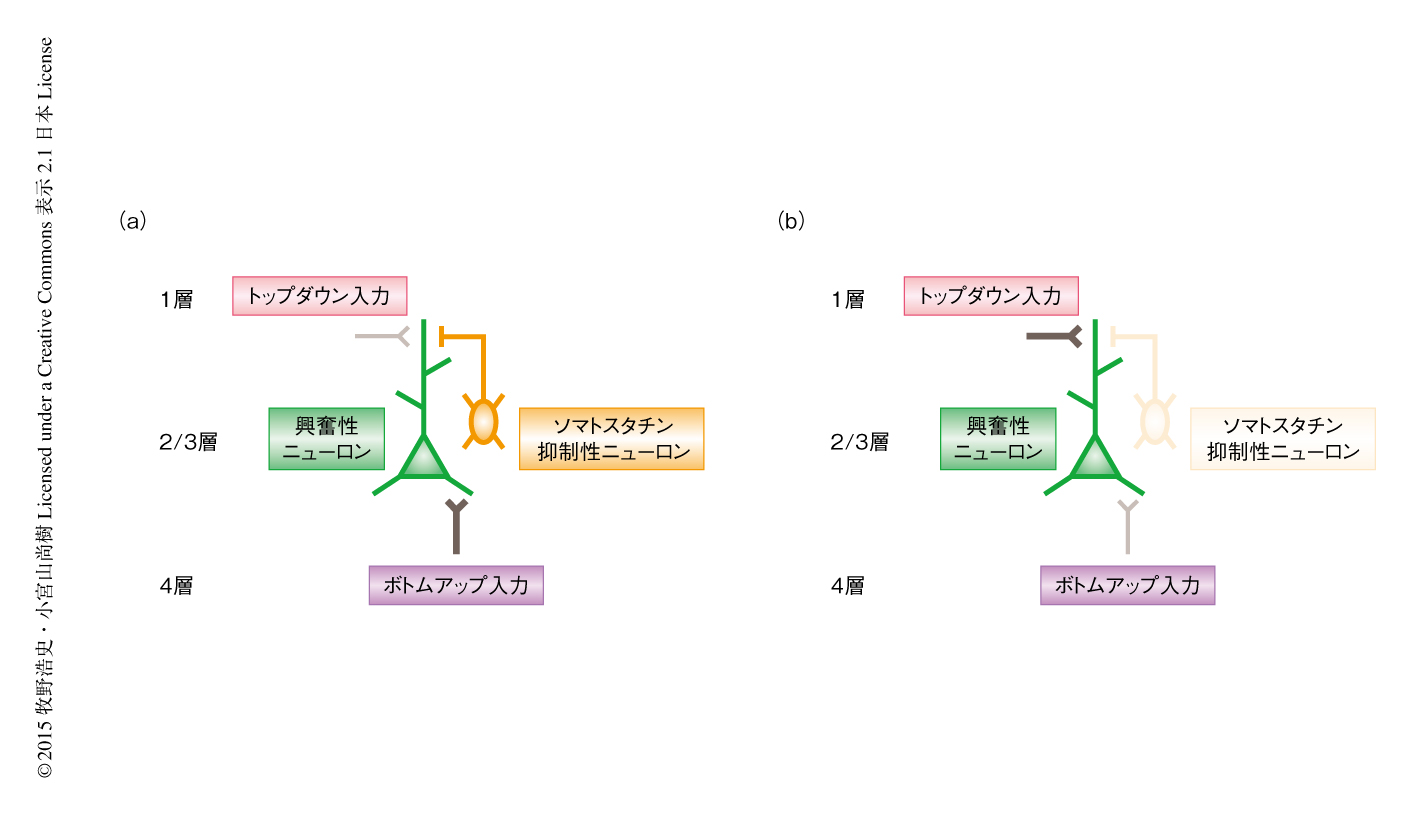
この研究は,この理論を検証するため,1次視覚野の2/3層における興奮性ニューロンに焦点をしぼり,外界から視床をつうじ伝達された4層からのボトムアップ入力と脳梁膨大後部皮質からのトップダウン入力が連合学習中にどのように変化するか評価した.4層からのボトムアップ入力は2/3層の興奮性ニューロンの細胞体の周辺に,脳梁膨大後部皮質からのトップダウン入力は1層の尖端の樹状突起に興奮性シナプスを形成する3).この空間的な分離は2つの異なる情報が同じ細胞の別々の場所において処理されていることを意味している.また,これらの情報の流れを制御すると考えられている多様な抑制性ニューロンの活動について調べ,学習による情報処理の変化の機構にもせまった.
マウスに脳を固定した状態で視覚情報による能動回避課題を4日間にわたり学習させた.この課題では,マウスは視覚情報があたえられているあいだトレッドミルを走り,あらかじめ定められたある一定の速度に達することにより微弱な電気刺激を回避する.学習をはじめてから4日間たつと成功率は約90%に到達した.先行研究により,このような視覚による能動回避学習には脳梁膨大後部皮質が必要であることが知られているが4),この研究により,脳梁膨大後部皮質とともに1次視覚野もこの課題に必要であることが明らかにされた.
2/3層の興奮性ニューロン,そのボトムアップ入力のもとである4層の興奮性ニューロン,脳梁膨大後部皮質から1次視覚野へと伝達される興奮性のトップダウン入力の活動について調べるため,アデノ随伴ウイルスを用いてCa2+指示薬であるGCaMP6 5) をそれぞれの神経回路の素子に導入した.抑制性ニューロンにおいて蛍光タンパク質tdTomatoを発現する遺伝子改変マウス6-8) を用いることにより,2/3層および4層における興奮性ニューロンと抑制性ニューロンとを区別した.また,GCaMP6を脳梁膨大後部皮質の細胞体に発現させ1次視覚野の1層へと投射する軸索を可視化することにより,トップダウン入力の活動の長期的な観察も可能になった.このような手法により,個々の神経回路の素子の活動を2光子顕微鏡によるCa2+イメージング法により測定したところ,連合学習中に4層の興奮性ニューロンの活動はしだいに低下したのに対し,脳梁膨大後部皮質からのトップダウン入力を投射する軸索の活動は上昇することがわかった(図2).このような2つの情報処理機構の非対称な変化は,学習により獲得された内部予測モデルにしたがい,トップダウン入力による予測信号の正確性は最大化されるのに対し,ボトムアップ入力による予測エラー信号は最小化されるという,さきに述べた理論と一致した.
この実験では,学習において脳梁膨大後部皮質からのトップダウン入力が増大することが示されたが,実際に,2/3層の興奮性ニューロンへの影響はどう変わるのであろうか.この問いを検証するため,2/3層の興奮性ニューロンの視覚刺激に対する時間的な反応について調べたところ,学習のまえには視覚刺激に忠実に反応したのに対し,学習のあとでは視覚刺激があたえられているあいだCa2+シグナルを徐々に上昇させ,電気刺激のタイミングにてCa2+シグナルのピークが現われた.このような反応は,先行研究により示された1次視覚野において報酬のタイミングをコードすると考えられている反応9) に酷似しており,この実験から,1次視覚野のニューロンは嫌悪刺激のタイミングもコードしている可能性が示唆された.
学習による2/3層の興奮性ニューロンの時間的な反応の変化はトップダウン入力の影響によるものなのかどうか検証するため,脳梁膨大後部皮質からのトップダウン入力を投射する軸索の活動について調べたところ,2/3層の興奮性ニューロンと同じく電気刺激を予期するような反応がみられた.対照的に,4層の興奮性ニューロンにはそのような変化はみられなかった.この結果から,2/3層の興奮性ニューロンは学習によりトップダウン入力の影響をよりうけるようになることが示唆された.
2/3層の興奮性ニューロンと脳梁膨大後部皮質からのトップダウン入力を投射する軸索との活動の変化における因果関係について調べるため,学習ののち脳梁膨大後部皮質を抑制しトップダウン入力を減少させたところ,2/3層の興奮性ニューロンは学習のまえのような視覚刺激に忠実な反応にもどった.これらの結果より,学習はトップダウン入力の影響を増大させることが明らかにされた.
近年,遺伝学の発達により大脳皮質における抑制性ニューロンの機能的,生理的,解剖学的な多様性が明らかにされた10,11).たとえば,パルブアルブミン陽性抑制性ニューロンは興奮性ニューロンの細胞体の周辺を抑制するのに対し,ソマトスタチン陽性抑制性ニューロンは軸索を1層に伸長し興奮性ニューロンの尖端の樹状突起を抑制する.この異なった種類の細胞による抑制の空間的な解離はボトムアップ入力とトップダウン入力との空間的な解離と似ており,パルブアルブミン陽性抑制性ニューロンがボトムアップ入力の,ソマトスタチン陽性抑制性ニューロンがトップダウン入力の制御にかかわることが示唆される12).これまでの実験においては,学習によりトップダウン入力の影響が増大することが示されたが,もし,ソマトスタチン陽性細胞がトップダウン入力の制御にかかわっているとするのなら,その活動は学習中に低下するのではないかと予想された.SOM-Cre遺伝子改変マウス6) を用いることによりGCaMP6をソマトスタチン陽性抑制性ニューロンに特異的に発現させ,学習中にその活動を調べたところ,ソマトスタチン陽性抑制性ニューロンの活動はしだいに低下することがわかった.このような尖端の樹状突起における脱抑制により2/3層の興奮性ニューロンはトップダウン入力の影響をよりうけるようになると考えられる(図2).
脳のおのおのの領域は多様な入力をうけており,状況に応じそれぞれの入力の比重を適切に制御することが重要である.この研究においては,1次視覚野の2/3層の興奮性ニューロンを中心に,学習による情報処理形態の変化について検証した.また,このような変化に対し,ソマトスタチン陽性抑制性ニューロンがどのような制御をしているのか明らかにされた.これらの結果から,学習におけるトップダウン入力とボトムアップ入力のバランスの動的な変化が局所の神経回路と脳の領域のあいだの入力との複雑な相互関係によりなされるものであることが示された.
予測コーディング理論によれば,内部予測モデルにしたがいトップダウン入力による予測信号が下の層へと伝達され,上の層へと伝達されるボトムアップ入力による予測エラー信号を抑制する.この研究は,これら2つの情報処理機構が学習中にどう変化するのかを明らかにしたが,今後,実際にトップダウン入力はボトムアップ入力を抑制するのか,また,その神経回路機構についての解明が期待される.
略歴:2010年 米国Cold Spring Harbor Laboratory博士課程 修了,同年より米国California大学San Diego校 博士研究員.
研究テーマ:理論と実験による神経回路の動作原理の探求.
小宮山 尚樹(Takaki Komiyama)
米国California大学San Diego校Assistant Professor.
© 2015 牧野浩史・小宮山尚樹 Licensed under CC 表示 2.1 日本
(米国California大学San Diego校Department of Neurosciences)
email:牧野浩史,小宮山尚樹
DOI: 10.7875/first.author.2015.092
Learning enhances the relative impact of top-down processing in the visual cortex.
Hiroshi Makino, Takaki Komiyama
Nature Neuroscience, 18, 1116-1122 (2015)
要 約
学習とは,外界に対し脳において内部予測モデルを構築し,高次の脳領域からの予測や期待あるいは注意などの情報をつかさどるトップダウン入力を増大させるとともに,外界から得られる情報を処理するボトムアップ入力を減少させるものと考えられている.筆者らは,この理論を検証するため,視覚情報による連合学習中の1次視覚野の2/3層の興奮性ニューロン,そのボトムアップ入力のもとである4層の興奮性ニューロン,トップダウン入力である脳梁膨大後部皮質から直接に投射する興奮性の軸索の活動を2光子Ca2+イメージング法を用いて評価した.これにより,4層の活動はしだいに低下するのに対し,脳梁膨大後部皮質からの軸索の活動は上昇し2/3層への影響を強めることが明らかにされた.それとともに,抑制性ニューロンでありトップダウン入力を制御すると考えられているソマトスタチン陽性細胞の活動も学習をつうじ低下することがわかった.これらの結果から,学習がボトムアップ入力とトップダウン入力の情報処理機構のバランスに動的なシフトを促すとともに,その微小回路における神経機構も明らかにされた.
はじめに
予測コーディング理論によると,脳は外界からの感覚入力をうまく説明できるような内部予測モデルを構築し,未来に起こりうる現象に対し適切な対応ができるようにすると考えられている1,2).この理論によれば,脳は階層構造をとっており,学習により得られた内部予測モデルにしたがい上の層から下の層へと予測信号(トップダウン入力)を伝達する.もし,この予測信号が実際の感覚入力と違う場合,下の層から上の層へと予測エラー信号(ボトムアップ入力)が伝達され,内部予測モデルは更新される.学習とはまさにこのような階層のあいだの相互作用により脳において内部予測モデルを徐々に最適化させる過程のことであり,脳は予測信号の正確性を最大化し,予測エラー信号を最小化させることを目的にしていると考えられる(図1).
この研究は,この理論を検証するため,1次視覚野の2/3層における興奮性ニューロンに焦点をしぼり,外界から視床をつうじ伝達された4層からのボトムアップ入力と脳梁膨大後部皮質からのトップダウン入力が連合学習中にどのように変化するか評価した.4層からのボトムアップ入力は2/3層の興奮性ニューロンの細胞体の周辺に,脳梁膨大後部皮質からのトップダウン入力は1層の尖端の樹状突起に興奮性シナプスを形成する3).この空間的な分離は2つの異なる情報が同じ細胞の別々の場所において処理されていることを意味している.また,これらの情報の流れを制御すると考えられている多様な抑制性ニューロンの活動について調べ,学習による情報処理の変化の機構にもせまった.
1.視覚情報による能動回避学習
マウスに脳を固定した状態で視覚情報による能動回避課題を4日間にわたり学習させた.この課題では,マウスは視覚情報があたえられているあいだトレッドミルを走り,あらかじめ定められたある一定の速度に達することにより微弱な電気刺激を回避する.学習をはじめてから4日間たつと成功率は約90%に到達した.先行研究により,このような視覚による能動回避学習には脳梁膨大後部皮質が必要であることが知られているが4),この研究により,脳梁膨大後部皮質とともに1次視覚野もこの課題に必要であることが明らかにされた.
2.ボトムアップ入力とトップダウン入力の非対称な変化
2/3層の興奮性ニューロン,そのボトムアップ入力のもとである4層の興奮性ニューロン,脳梁膨大後部皮質から1次視覚野へと伝達される興奮性のトップダウン入力の活動について調べるため,アデノ随伴ウイルスを用いてCa2+指示薬であるGCaMP6 5) をそれぞれの神経回路の素子に導入した.抑制性ニューロンにおいて蛍光タンパク質tdTomatoを発現する遺伝子改変マウス6-8) を用いることにより,2/3層および4層における興奮性ニューロンと抑制性ニューロンとを区別した.また,GCaMP6を脳梁膨大後部皮質の細胞体に発現させ1次視覚野の1層へと投射する軸索を可視化することにより,トップダウン入力の活動の長期的な観察も可能になった.このような手法により,個々の神経回路の素子の活動を2光子顕微鏡によるCa2+イメージング法により測定したところ,連合学習中に4層の興奮性ニューロンの活動はしだいに低下したのに対し,脳梁膨大後部皮質からのトップダウン入力を投射する軸索の活動は上昇することがわかった(図2).このような2つの情報処理機構の非対称な変化は,学習により獲得された内部予測モデルにしたがい,トップダウン入力による予測信号の正確性は最大化されるのに対し,ボトムアップ入力による予測エラー信号は最小化されるという,さきに述べた理論と一致した.
3.トップダウン入力の2/3層への影響の増大
この実験では,学習において脳梁膨大後部皮質からのトップダウン入力が増大することが示されたが,実際に,2/3層の興奮性ニューロンへの影響はどう変わるのであろうか.この問いを検証するため,2/3層の興奮性ニューロンの視覚刺激に対する時間的な反応について調べたところ,学習のまえには視覚刺激に忠実に反応したのに対し,学習のあとでは視覚刺激があたえられているあいだCa2+シグナルを徐々に上昇させ,電気刺激のタイミングにてCa2+シグナルのピークが現われた.このような反応は,先行研究により示された1次視覚野において報酬のタイミングをコードすると考えられている反応9) に酷似しており,この実験から,1次視覚野のニューロンは嫌悪刺激のタイミングもコードしている可能性が示唆された.
学習による2/3層の興奮性ニューロンの時間的な反応の変化はトップダウン入力の影響によるものなのかどうか検証するため,脳梁膨大後部皮質からのトップダウン入力を投射する軸索の活動について調べたところ,2/3層の興奮性ニューロンと同じく電気刺激を予期するような反応がみられた.対照的に,4層の興奮性ニューロンにはそのような変化はみられなかった.この結果から,2/3層の興奮性ニューロンは学習によりトップダウン入力の影響をよりうけるようになることが示唆された.
2/3層の興奮性ニューロンと脳梁膨大後部皮質からのトップダウン入力を投射する軸索との活動の変化における因果関係について調べるため,学習ののち脳梁膨大後部皮質を抑制しトップダウン入力を減少させたところ,2/3層の興奮性ニューロンは学習のまえのような視覚刺激に忠実な反応にもどった.これらの結果より,学習はトップダウン入力の影響を増大させることが明らかにされた.
4.学習が及ぼす抑制性ニューロンの変化
近年,遺伝学の発達により大脳皮質における抑制性ニューロンの機能的,生理的,解剖学的な多様性が明らかにされた10,11).たとえば,パルブアルブミン陽性抑制性ニューロンは興奮性ニューロンの細胞体の周辺を抑制するのに対し,ソマトスタチン陽性抑制性ニューロンは軸索を1層に伸長し興奮性ニューロンの尖端の樹状突起を抑制する.この異なった種類の細胞による抑制の空間的な解離はボトムアップ入力とトップダウン入力との空間的な解離と似ており,パルブアルブミン陽性抑制性ニューロンがボトムアップ入力の,ソマトスタチン陽性抑制性ニューロンがトップダウン入力の制御にかかわることが示唆される12).これまでの実験においては,学習によりトップダウン入力の影響が増大することが示されたが,もし,ソマトスタチン陽性細胞がトップダウン入力の制御にかかわっているとするのなら,その活動は学習中に低下するのではないかと予想された.SOM-Cre遺伝子改変マウス6) を用いることによりGCaMP6をソマトスタチン陽性抑制性ニューロンに特異的に発現させ,学習中にその活動を調べたところ,ソマトスタチン陽性抑制性ニューロンの活動はしだいに低下することがわかった.このような尖端の樹状突起における脱抑制により2/3層の興奮性ニューロンはトップダウン入力の影響をよりうけるようになると考えられる(図2).
おわりに
脳のおのおのの領域は多様な入力をうけており,状況に応じそれぞれの入力の比重を適切に制御することが重要である.この研究においては,1次視覚野の2/3層の興奮性ニューロンを中心に,学習による情報処理形態の変化について検証した.また,このような変化に対し,ソマトスタチン陽性抑制性ニューロンがどのような制御をしているのか明らかにされた.これらの結果から,学習におけるトップダウン入力とボトムアップ入力のバランスの動的な変化が局所の神経回路と脳の領域のあいだの入力との複雑な相互関係によりなされるものであることが示された.
予測コーディング理論によれば,内部予測モデルにしたがいトップダウン入力による予測信号が下の層へと伝達され,上の層へと伝達されるボトムアップ入力による予測エラー信号を抑制する.この研究は,これら2つの情報処理機構が学習中にどう変化するのかを明らかにしたが,今後,実際にトップダウン入力はボトムアップ入力を抑制するのか,また,その神経回路機構についての解明が期待される.
文 献
- Friston, K.: The free-energy principle: a unified brain theory? Nat. Rev. Neurosci., 11, 127-138 (2010)[PubMed]
- Rao, R. P. & Ballard, D. H.: Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 2, 79-87 (1999)[PubMed]
- Petreanu, L., Mao, T., Sternson, S. M. et al.: The subcellular organization of neocortical excitatory connections. Nature, 457, 1142-1145 (2009)[PubMed]
- Lukoyanov, N. V. & Lukoyanova, E. A.: Retrosplenial cortex lesions impair acquisition of active avoidance while sparing fear-based emotional memory. Behav. Brain Res., 173, 229-236 (2006)[PubMed]
- Chen, T. W., Wardill, T. J., Sun, Y. et al.: Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature, 499, 295-300 (2013)[PubMed]
- Taniguchi, H., He, M., Wu, P. et al.: A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron, 71, 995-1013 (2011)[PubMed]
- Madisen, L., Zwingman, T. A., Sunkin, S. M. et al.: A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci., 13, 133-140 (2010)[PubMed]
- Peters, A. J., Chen, S. X. & Komiyama, T.: Emergence of reproducible spatiotemporal activity during motor learning. Nature, 510, 263-267 (2014)[PubMed]
- Shuler, M. G. & Bear, M. F.: Reward timing in the primary visual cortex. Science, 311, 1606-1609 (2006)[PubMed]
- Kawaguchi, Y. & Kubota, Y.: GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cereb. Cortex, 7, 476-486 (1997)[PubMed]
- Markram, H., Toledo-Rodriguez, M., Wang, Y. et al.: Interneurons of the neocortical inhibitory system. Nat. Rev. Neurosci., 5, 793-807 (2004)[PubMed]
- Gentet, L. J., Kremer, Y., Taniguchi, H. et al.: Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex. Nat. Neurosci., 15, 607-612 (2012)[PubMed]
著者プロフィール
略歴:2010年 米国Cold Spring Harbor Laboratory博士課程 修了,同年より米国California大学San Diego校 博士研究員.
研究テーマ:理論と実験による神経回路の動作原理の探求.
小宮山 尚樹(Takaki Komiyama)
米国California大学San Diego校Assistant Professor.
© 2015 牧野浩史・小宮山尚樹 Licensed under CC 表示 2.1 日本
