ケミカルジェネティクスにより明らかとなったストリゴラクトンの植物ホルモンとしての新たな機能
土屋 雄一朗
(カナダToronto大学Department of Cell & Systems Biology)
email:土屋雄一朗
DOI: 10.7875/first.author.2010.026
A small-molecule screen identifies new functions for the plant hormone strigolactone.
Yuichiro Tsuchiya, Danielle Vidaurre, Shigeo Toh, Atsushi Hanada, Eiji Nambara, Yuji Kamiya, Shinjiro Yamaguchi, Peter McCourt
Nature Chemical Biology, 6, 741-749 (2010)
寄生植物であるストライガによる被害はアフリカの農地で拡大しており,貧しい農民に巨大な被害をあたえている.ストライガの種は宿主植物が放出する低分子化合物ストリゴラクトンを認識して発芽することが知られており,このステップを理解することでストライガ防除への道が開かれるものと信じられる.筆者らは,シロイヌナズナを使ったケミカルスクリーニングから,ストリゴラクトンの生産量を増やすコチルイミドと名づけた低分子化合物を同定した.このコチルイミドを使った耐性突然変異株のスクリーニングから,光受容体がストリゴラクトンの生産量を制御することが見い出され,発芽欠損などのさまざまな表現型はストリゴラクトンをあたえることでレスキューできることが明らかとなった.さらに,ストリゴラクトンはCOP1ユビキチンリガーゼ,転写因子HY5を経由して光誘導遺伝子の発現を制御することを発見した.ここでの発見は,ストリゴラクトンの新しい機能にとどまらず,ストライガの防除にむけさまざまな道を示したものである.
われわれが道ばたでよくみる植物は光合成を行うことで自ら栄養を生産できる.そんな植物のなかにも,自分で栄養を生産することを放棄して,まるで動物のようにほかの植物から栄養を搾取して生きるものが存在する.寄生植物であるストライガは,東欧の民話にでてくるストリゴイという吸血鬼にちなんで名づけられた植物の吸血鬼である.しかも,アフリカでは農作物への寄生による被害が拡大しており,年間で数十億ドルにものぼる被害を貧しい農民にあたえるという破壊的な吸血鬼として存在する.この問題は,われわれ植物学者が解決しなくてはならない大きな問題である.
ストライガの種は,通常は土のなかで何年も発芽せずに控えている.そして,宿主植物が近くで生育しているときにかぎり,その根から放出されるストリゴラクトンとよばれる低分子化合物を認識して発芽し,宿主植物の根に寄生しようとする.この発芽のステップを理解できればストライガの防除への道が開かれるのはわかっているが,実験室で生育するのがむずかしいストライガでは遺伝学的な手法が使えないため,ストライガがどのようにストリゴラクトンを受容して発芽するかについてはほとんどわかっていなかった.ここでは,ストリゴラクトンの宿主植物の内部での機能を低分子化合物を使ったアプローチで解明した研究を紹介する.この研究過程で判明したストリゴラクトンの新たな機能は,ストライガが自家栄養からストリゴラクトンによって発芽する寄生植物へとどのように進化したかの道筋を示すものと考えられる.
通常,ホルモンは生体内でつくられて機能する低分子化合物のことをさすが,ストリゴラクトンは植物自体にくわえて,そのまわりの生物環境にまで影響を及ぼす低分子化合物である.植物体内では枝分かれパターンを制御する植物ホルモンとして機能し,それと同時に,根から放出されることで共生菌根菌を引き寄せるアレロケミカルとしての作用もある1-3).一方,寄生植物は菌根菌を引き寄せる作用を横取りして宿主植物の検出に使う,といった構図が考えられている.このように,宿主植物でつくられるストリゴラクトンは3種類の生物で共通に使われているものの,宿主植物では枝分かれ,寄生植物では発芽,菌根菌には誘因因子,といったように,その機能には共通性がうかがえない.とくに,進化的に離れている菌根菌はまだしも,自家栄養植物から進化した寄生植物が,なぜ宿主植物ではみられないような応答をみせるのかは謎であった.
まるで突然変異株をスクリーニングするように,数万もの低分子化合物のライブラリーから目的の表現型を誘導する化合物をみつけ,それをバイオプローブとして生命現象の理解に使うケミカルジェネティクスとよばれる手法が,近年,発展してきている.継代する必要もなく,また,化合物をあたえるだけで表現型がみられるため,たとえば,致死のように突然変異株ではできないような解析をも可能とする新たな手法である.筆者らはまず,種子発芽や子葉の形態に注目してモデル植物であるシロイヌナズナを使ったケミカルスクリーニングを行った.それにより,10,000の化合物ライブラリーから子葉にのみ成長抑制作用を示す5種類の類縁化合物を発見し,これらをコチルイミドと名づけた.この名前は,子葉(コチレドン)と類縁化合物に共通に含まれる部分構造(フタルイミドまたはスクシンイミド)に由来する.
フタルイミドやスクシンイミドを部分構造としてもつ生理活性化合物は数多く知られているものの,ケミカルクラスタリング解析からは,そのなかのひとつNijmegen-1とよばれる人工ストリゴラクトンがコチルイミドともっともよく似ていると判断された.これをきっかけとして,コチルイミドとストリゴラクトンとのあいだにどういった関連があるかを調べはじめた.人工ストリゴラクトンであるGR24をシロイヌナズナに大量にあたえるとコチルイミドのように子葉の成長を抑制すること,さらに,DNAマイクロアレイによる共発現解析でGR24とコチルイミドとは高い相関を示すことが明らかとなり,この2つの化合物はよく似た生理活性をもつものと考えられた.しかし,これらの結果とはうらはらに,コチルイミドはストライガの発芽を刺激しないためストリゴラクトン活性をもっていない.似た活性をもっているにもかかわらず同じ活性を示さないという矛盾は,コチルイミドにストリゴラクトンの生産促進(あるいは,分解抑制)作用があることがイネを使った液体クロマトグラフィー-タンデム質量(LC-MS/MS)分析より判明したため無事に解決された.この結果は,GR24をコチルイミドと混ぜてあたえると,ごく低濃度で子葉の成長を抑制することによっても支持される.これらの結果をまとめると,ストリゴラクトンの内在量を増やすことは子葉の成長抑制に”十分”と考えられる.では,”必要”のほうはどうだろうか.ストリゴラクトンの生産や認識に欠損をもつmax突然変異株を使って調べたところ,予想どおり,すべてコチルイミドに耐性であった4).総括して,コチルイミドの子葉の成長抑制作用にストリゴラクトンの増加は必要十分といえた.
ストリゴラクトンの生産や認識に欠損をもつmax突然変異株がコチルイミドに耐性であるので,逆に,コチルイミド耐性の突然変異株をスクリーニングすればストリゴラクトンの生産や受容の欠損株が得られるはずである.そこで,シロイヌナズナの変異種子500,000をスクリーニングし,約240株のコチルイミド耐性株を得た.おもしろいことに,そのうちの5株はコチルイミドやGR24をあたえないとほとんど発芽しないという表現型を示した.発芽にストリゴラクトンを必要とするのは,まさにストライガと同じである.そこで,この5株に注目して解析を進めた結果,これらの突然変異株では培地に放出される寄生植物の発芽刺激物質が少ないことが明らかとなった(のちに,イネのオーソログ突然変異株を使ったLC-MS/MS分析によりストリゴラクトンの生産量が少ないことが判明した).ストリゴラクトンの生産量が少ない突然変異株では発芽欠損がみられ,ここにストリゴラクトンをあたえることで発芽するようになるということは,ストリゴラクトンがシロイヌナズナの発芽を刺激することを意味する.寄生植物以外でもストリゴラクトンが発芽を刺激するというのは特筆すべき発見である.
相補性検定からこれら5株が単一の遺伝子アリルであることがわかった.この遺伝子はストリゴラクトンの生合成にかかわる酵素をコードしているものと期待していたが,遺伝学マッピングの結果,予想外にも,光受容体であるフィトクロムの機能にかかわるHY1遺伝子において独立な突然変異が発見された5,6).
光は自家栄養植物にとってとくに重要な環境因子である.光は多くの植物で発芽を刺激し,光合成を効率よく行えるよう生育を調節する.そのため,光を認識できない突然変異株には発芽欠損やがみられたり,光を当ててももやしのように胚軸伸張がみられたりする.これまで,そういった胚軸の長いような突然変異株のスクリーニングからhy1からhy5までの突然変異株が同定され,HY1遺伝子とHY2遺伝子はフィトクロムのクロモフォア合成酵素,HY3遺伝子はフィトクロムB光受容体,HY4遺伝子はクリプトクロム光受容体,HY5遺伝子はその下流ではたらく転写因子,をそれぞれコードすることがわかっていた7).さて,hy1突然変異株の発芽欠損をストリゴラクトンが抑圧できるということは,ストリゴラクトンが光受容の下流を活性化することを示唆した.もし,これが正しいのなら,ストリゴラクトンは発芽欠損のほかにも胚軸伸張も抑圧できるはずである.この仮説を検証するため,hy1突然変異株にくわえて,フィトクロムとクリプトクロムの突然変異株,および,hy5突然変異株の胚軸伸張に対するGR24の作用を調べた.その結果,仮説どおりに,GR24はhy1突然変異株と光受容体の突然変異株の胚軸伸張を抑えることがわかり,さらに,hy5突然変異株がGR24に応答しないことが明らかとなった.このことから,ストリゴラクトンは光受容の下流,かつ,HY5遺伝子の上流に作用するものと考えられた.発現解析の結果,GR24をあたえてわずか30分足らずでHY5タンパク質の蓄積がみられ,HY5遺伝子はストリゴラクトンのシグナルを伝える重要な因子と考えられた.
COP1ユビキチンリガーゼは光受容体と転写因子HY5とのあいだに存在する負の制御因子である.暗所ではCOP1ユビキチンリガーゼがHY5に結合して分解を促進する.一方,光を当てると光受容体がCOP1ユビキチンリガーゼを核から排出することでHY5が核に蓄積し,光誘導性遺伝子が発現する8).そのため,cop1突然変異株では暗所でも光を当てたときように胚軸伸張が起こらず,HY5が蓄積し光誘導遺伝子の発現がみられる.ストリゴラクトンの作用点が光受容とHY5遺伝子とのあいだにあり,HY5タンパク質の蓄積を促進するなら,ストリゴラクトンがCOP1ユビキチンリガーゼの機能を阻害する可能性が考えられる.この仮説どおり,cop1突然変異株(漏出性アリル)はGR24に高感受性を示し,COP1過剰発現体でみられる胚軸伸張はGR24をあたえることで抑圧された.さらに,暗所で育てた植物にGR24をあたえるとCOP1ユビキチンリガーゼが核から排出されることがGUSレポーターコンストラクトを使った実験から明らかとなった.
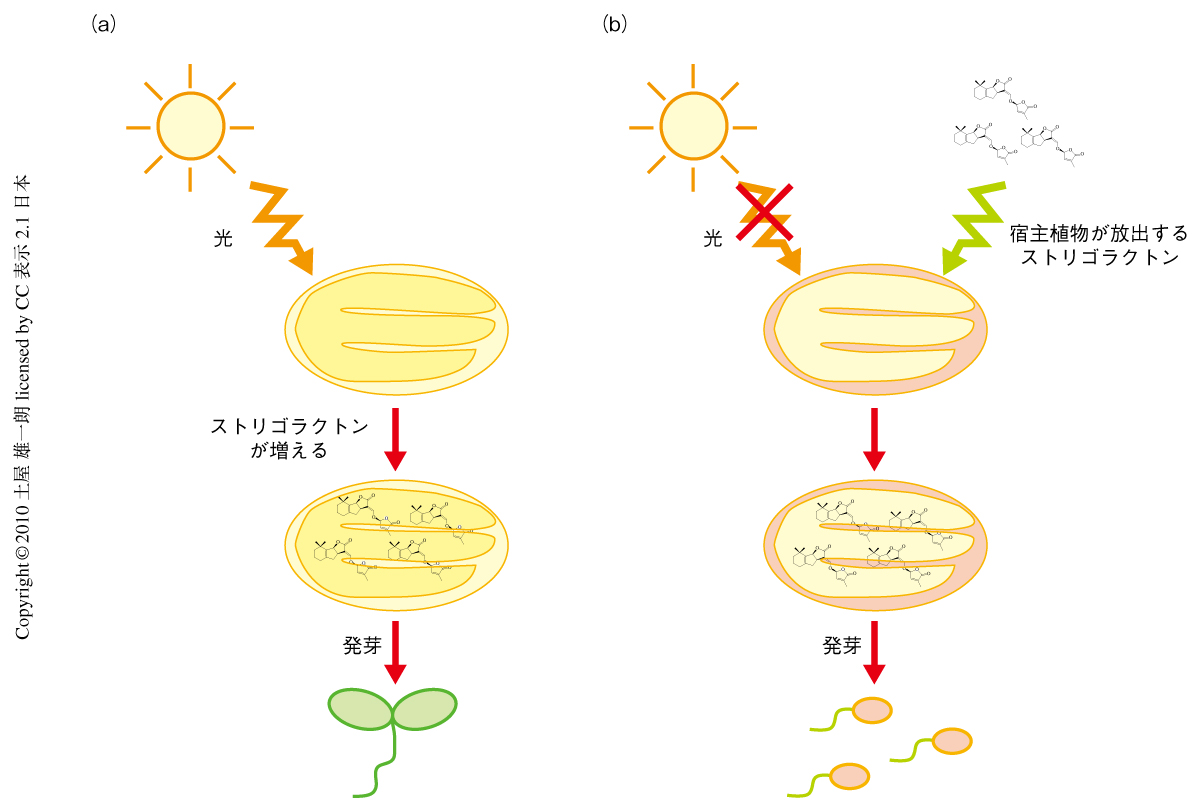
ストリゴラクトンと光はどちらもCOP1ユビキチンリガーゼを核から排出する.ならば,ストリゴラクトンは光の代わりをできるのではないだろうか? そこで,暗所にてGR24をあたえて生育させてみると,光を当てて育てたように胚軸の伸張が起こらず,HY5タンパク質の蓄積がみられ,光誘導遺伝子の発現が確認された.この研究を総括して,種子発芽の促進,胚軸伸張の抑制,光誘導遺伝子の発現など,調べたすべてにおいてストリゴラクトンには光と同じ作用があることが判明した(図1).

この研究では,コチルイミドの発見から一連の解析をへて,ストリゴラクトンの植物体内での新たな役割の発見にいたった.ここで発見された光シグナル伝達にかかわる因子やコチルイミドは,ストライガの防除にむけ新しい可能性を示したことだろう.たとえば,コチルイミドをストライガの宿主とはならない植物にあたえることでストリゴラクトンを過剰に生産させストライガの自殺発芽を誘導したり,光シグナル伝達遺伝子を使ったストライガ耐性作物の育種へ応用したりすることが考えられる.また,ストリゴラクトンが発芽を誘導する分子機構がシロイヌナズナとストライガとのあいだで保存されているなら,シロイヌナズナを使って新たな遺伝子や低分子化合物を発見することも可能である.
ストライガのような寄生植物は必ずしも光に応答する必要はない.ひとたび寄生する能力を獲得してしまうと,このような選択圧のゆるい環境ではhy1突然変異株のように光応答が退化してしまうのだろう.すると,hy1突然変異株のようにストリゴラクトンをつくれなくなり,発芽するためにはほかの植物が放出するストリゴラクトンに依存しなくてはならない.ストライガはこのようにして進化してきたのかもしれない(図2).
略歴:2002年 北海道大学大学院農学研究科 修了,同年 カナダToronto大学 ポスドク研究員,2006年 理化学研究所植物科学研究センター 基礎科学特別研究員を経て,2009年よりカナダToronto大学 ポスドク研究員.
研究テーマ:ストリゴラクトンのシグナル伝達メカニズムの解析.
考察:研究におけるギャンブルとリスクマネージメント.
© 2010 土屋 雄一朗 Licensed under CC 表示 2.1 日本
(カナダToronto大学Department of Cell & Systems Biology)
email:土屋雄一朗
DOI: 10.7875/first.author.2010.026
A small-molecule screen identifies new functions for the plant hormone strigolactone.
Yuichiro Tsuchiya, Danielle Vidaurre, Shigeo Toh, Atsushi Hanada, Eiji Nambara, Yuji Kamiya, Shinjiro Yamaguchi, Peter McCourt
Nature Chemical Biology, 6, 741-749 (2010)
要 約
寄生植物であるストライガによる被害はアフリカの農地で拡大しており,貧しい農民に巨大な被害をあたえている.ストライガの種は宿主植物が放出する低分子化合物ストリゴラクトンを認識して発芽することが知られており,このステップを理解することでストライガ防除への道が開かれるものと信じられる.筆者らは,シロイヌナズナを使ったケミカルスクリーニングから,ストリゴラクトンの生産量を増やすコチルイミドと名づけた低分子化合物を同定した.このコチルイミドを使った耐性突然変異株のスクリーニングから,光受容体がストリゴラクトンの生産量を制御することが見い出され,発芽欠損などのさまざまな表現型はストリゴラクトンをあたえることでレスキューできることが明らかとなった.さらに,ストリゴラクトンはCOP1ユビキチンリガーゼ,転写因子HY5を経由して光誘導遺伝子の発現を制御することを発見した.ここでの発見は,ストリゴラクトンの新しい機能にとどまらず,ストライガの防除にむけさまざまな道を示したものである.
はじめに
われわれが道ばたでよくみる植物は光合成を行うことで自ら栄養を生産できる.そんな植物のなかにも,自分で栄養を生産することを放棄して,まるで動物のようにほかの植物から栄養を搾取して生きるものが存在する.寄生植物であるストライガは,東欧の民話にでてくるストリゴイという吸血鬼にちなんで名づけられた植物の吸血鬼である.しかも,アフリカでは農作物への寄生による被害が拡大しており,年間で数十億ドルにものぼる被害を貧しい農民にあたえるという破壊的な吸血鬼として存在する.この問題は,われわれ植物学者が解決しなくてはならない大きな問題である.
ストライガの種は,通常は土のなかで何年も発芽せずに控えている.そして,宿主植物が近くで生育しているときにかぎり,その根から放出されるストリゴラクトンとよばれる低分子化合物を認識して発芽し,宿主植物の根に寄生しようとする.この発芽のステップを理解できればストライガの防除への道が開かれるのはわかっているが,実験室で生育するのがむずかしいストライガでは遺伝学的な手法が使えないため,ストライガがどのようにストリゴラクトンを受容して発芽するかについてはほとんどわかっていなかった.ここでは,ストリゴラクトンの宿主植物の内部での機能を低分子化合物を使ったアプローチで解明した研究を紹介する.この研究過程で判明したストリゴラクトンの新たな機能は,ストライガが自家栄養からストリゴラクトンによって発芽する寄生植物へとどのように進化したかの道筋を示すものと考えられる.
1.ストリゴラクトンとは
通常,ホルモンは生体内でつくられて機能する低分子化合物のことをさすが,ストリゴラクトンは植物自体にくわえて,そのまわりの生物環境にまで影響を及ぼす低分子化合物である.植物体内では枝分かれパターンを制御する植物ホルモンとして機能し,それと同時に,根から放出されることで共生菌根菌を引き寄せるアレロケミカルとしての作用もある1-3).一方,寄生植物は菌根菌を引き寄せる作用を横取りして宿主植物の検出に使う,といった構図が考えられている.このように,宿主植物でつくられるストリゴラクトンは3種類の生物で共通に使われているものの,宿主植物では枝分かれ,寄生植物では発芽,菌根菌には誘因因子,といったように,その機能には共通性がうかがえない.とくに,進化的に離れている菌根菌はまだしも,自家栄養植物から進化した寄生植物が,なぜ宿主植物ではみられないような応答をみせるのかは謎であった.
2.コチルイミドの発見
まるで突然変異株をスクリーニングするように,数万もの低分子化合物のライブラリーから目的の表現型を誘導する化合物をみつけ,それをバイオプローブとして生命現象の理解に使うケミカルジェネティクスとよばれる手法が,近年,発展してきている.継代する必要もなく,また,化合物をあたえるだけで表現型がみられるため,たとえば,致死のように突然変異株ではできないような解析をも可能とする新たな手法である.筆者らはまず,種子発芽や子葉の形態に注目してモデル植物であるシロイヌナズナを使ったケミカルスクリーニングを行った.それにより,10,000の化合物ライブラリーから子葉にのみ成長抑制作用を示す5種類の類縁化合物を発見し,これらをコチルイミドと名づけた.この名前は,子葉(コチレドン)と類縁化合物に共通に含まれる部分構造(フタルイミドまたはスクシンイミド)に由来する.
3.コチルイミドはストリゴラクトンの生産量を増やす
フタルイミドやスクシンイミドを部分構造としてもつ生理活性化合物は数多く知られているものの,ケミカルクラスタリング解析からは,そのなかのひとつNijmegen-1とよばれる人工ストリゴラクトンがコチルイミドともっともよく似ていると判断された.これをきっかけとして,コチルイミドとストリゴラクトンとのあいだにどういった関連があるかを調べはじめた.人工ストリゴラクトンであるGR24をシロイヌナズナに大量にあたえるとコチルイミドのように子葉の成長を抑制すること,さらに,DNAマイクロアレイによる共発現解析でGR24とコチルイミドとは高い相関を示すことが明らかとなり,この2つの化合物はよく似た生理活性をもつものと考えられた.しかし,これらの結果とはうらはらに,コチルイミドはストライガの発芽を刺激しないためストリゴラクトン活性をもっていない.似た活性をもっているにもかかわらず同じ活性を示さないという矛盾は,コチルイミドにストリゴラクトンの生産促進(あるいは,分解抑制)作用があることがイネを使った液体クロマトグラフィー-タンデム質量(LC-MS/MS)分析より判明したため無事に解決された.この結果は,GR24をコチルイミドと混ぜてあたえると,ごく低濃度で子葉の成長を抑制することによっても支持される.これらの結果をまとめると,ストリゴラクトンの内在量を増やすことは子葉の成長抑制に”十分”と考えられる.では,”必要”のほうはどうだろうか.ストリゴラクトンの生産や認識に欠損をもつmax突然変異株を使って調べたところ,予想どおり,すべてコチルイミドに耐性であった4).総括して,コチルイミドの子葉の成長抑制作用にストリゴラクトンの増加は必要十分といえた.
4.コチルイミドを使ったストリゴラクトンの生産量が少ない突然変異株の同定
ストリゴラクトンの生産や認識に欠損をもつmax突然変異株がコチルイミドに耐性であるので,逆に,コチルイミド耐性の突然変異株をスクリーニングすればストリゴラクトンの生産や受容の欠損株が得られるはずである.そこで,シロイヌナズナの変異種子500,000をスクリーニングし,約240株のコチルイミド耐性株を得た.おもしろいことに,そのうちの5株はコチルイミドやGR24をあたえないとほとんど発芽しないという表現型を示した.発芽にストリゴラクトンを必要とするのは,まさにストライガと同じである.そこで,この5株に注目して解析を進めた結果,これらの突然変異株では培地に放出される寄生植物の発芽刺激物質が少ないことが明らかとなった(のちに,イネのオーソログ突然変異株を使ったLC-MS/MS分析によりストリゴラクトンの生産量が少ないことが判明した).ストリゴラクトンの生産量が少ない突然変異株では発芽欠損がみられ,ここにストリゴラクトンをあたえることで発芽するようになるということは,ストリゴラクトンがシロイヌナズナの発芽を刺激することを意味する.寄生植物以外でもストリゴラクトンが発芽を刺激するというのは特筆すべき発見である.
相補性検定からこれら5株が単一の遺伝子アリルであることがわかった.この遺伝子はストリゴラクトンの生合成にかかわる酵素をコードしているものと期待していたが,遺伝学マッピングの結果,予想外にも,光受容体であるフィトクロムの機能にかかわるHY1遺伝子において独立な突然変異が発見された5,6).
5.ストリゴラクトンは光シグナルを伝達する転写因子HY5の蓄積を促進する
光は自家栄養植物にとってとくに重要な環境因子である.光は多くの植物で発芽を刺激し,光合成を効率よく行えるよう生育を調節する.そのため,光を認識できない突然変異株には発芽欠損やがみられたり,光を当ててももやしのように胚軸伸張がみられたりする.これまで,そういった胚軸の長いような突然変異株のスクリーニングからhy1からhy5までの突然変異株が同定され,HY1遺伝子とHY2遺伝子はフィトクロムのクロモフォア合成酵素,HY3遺伝子はフィトクロムB光受容体,HY4遺伝子はクリプトクロム光受容体,HY5遺伝子はその下流ではたらく転写因子,をそれぞれコードすることがわかっていた7).さて,hy1突然変異株の発芽欠損をストリゴラクトンが抑圧できるということは,ストリゴラクトンが光受容の下流を活性化することを示唆した.もし,これが正しいのなら,ストリゴラクトンは発芽欠損のほかにも胚軸伸張も抑圧できるはずである.この仮説を検証するため,hy1突然変異株にくわえて,フィトクロムとクリプトクロムの突然変異株,および,hy5突然変異株の胚軸伸張に対するGR24の作用を調べた.その結果,仮説どおりに,GR24はhy1突然変異株と光受容体の突然変異株の胚軸伸張を抑えることがわかり,さらに,hy5突然変異株がGR24に応答しないことが明らかとなった.このことから,ストリゴラクトンは光受容の下流,かつ,HY5遺伝子の上流に作用するものと考えられた.発現解析の結果,GR24をあたえてわずか30分足らずでHY5タンパク質の蓄積がみられ,HY5遺伝子はストリゴラクトンのシグナルを伝える重要な因子と考えられた.
6.ストリゴラクトンはCOP1ユビキチンリガーゼを核から排出する
COP1ユビキチンリガーゼは光受容体と転写因子HY5とのあいだに存在する負の制御因子である.暗所ではCOP1ユビキチンリガーゼがHY5に結合して分解を促進する.一方,光を当てると光受容体がCOP1ユビキチンリガーゼを核から排出することでHY5が核に蓄積し,光誘導性遺伝子が発現する8).そのため,cop1突然変異株では暗所でも光を当てたときように胚軸伸張が起こらず,HY5が蓄積し光誘導遺伝子の発現がみられる.ストリゴラクトンの作用点が光受容とHY5遺伝子とのあいだにあり,HY5タンパク質の蓄積を促進するなら,ストリゴラクトンがCOP1ユビキチンリガーゼの機能を阻害する可能性が考えられる.この仮説どおり,cop1突然変異株(漏出性アリル)はGR24に高感受性を示し,COP1過剰発現体でみられる胚軸伸張はGR24をあたえることで抑圧された.さらに,暗所で育てた植物にGR24をあたえるとCOP1ユビキチンリガーゼが核から排出されることがGUSレポーターコンストラクトを使った実験から明らかとなった.
7.ストリゴラクトンには光と同じはたらきがある
ストリゴラクトンと光はどちらもCOP1ユビキチンリガーゼを核から排出する.ならば,ストリゴラクトンは光の代わりをできるのではないだろうか? そこで,暗所にてGR24をあたえて生育させてみると,光を当てて育てたように胚軸の伸張が起こらず,HY5タンパク質の蓄積がみられ,光誘導遺伝子の発現が確認された.この研究を総括して,種子発芽の促進,胚軸伸張の抑制,光誘導遺伝子の発現など,調べたすべてにおいてストリゴラクトンには光と同じ作用があることが判明した(図1).
おわりに
この研究では,コチルイミドの発見から一連の解析をへて,ストリゴラクトンの植物体内での新たな役割の発見にいたった.ここで発見された光シグナル伝達にかかわる因子やコチルイミドは,ストライガの防除にむけ新しい可能性を示したことだろう.たとえば,コチルイミドをストライガの宿主とはならない植物にあたえることでストリゴラクトンを過剰に生産させストライガの自殺発芽を誘導したり,光シグナル伝達遺伝子を使ったストライガ耐性作物の育種へ応用したりすることが考えられる.また,ストリゴラクトンが発芽を誘導する分子機構がシロイヌナズナとストライガとのあいだで保存されているなら,シロイヌナズナを使って新たな遺伝子や低分子化合物を発見することも可能である.
ストライガのような寄生植物は必ずしも光に応答する必要はない.ひとたび寄生する能力を獲得してしまうと,このような選択圧のゆるい環境ではhy1突然変異株のように光応答が退化してしまうのだろう.すると,hy1突然変異株のようにストリゴラクトンをつくれなくなり,発芽するためにはほかの植物が放出するストリゴラクトンに依存しなくてはならない.ストライガはこのようにして進化してきたのかもしれない(図2).
文 献
- Umehara, M., Hanada, A., Yoshida, S. et al.: Inhibition of shoot branching by new terpenoid plant hormones. Nature, 455, 195-200 (2008)[PubMed]
- Gomez-Roldan, V., Fermas, S., Brewer, P. B. et al.: Strigolactone inhibition of shoot branching. Nature, 455, 189-194 (2008)[PubMed]
- Akiyama, K., Matsuzaki, K. & Hayashi, H.: Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature, 435, 824-827 (2005)[PubMed]
- Mouchel, C. F. & Leyser, O.: Novel phytohormones involved in long-range signaling. Curr. Opin. Plant Biol., 10, 473-476 (2007)[PubMed]
- Davis, S. J., Kurepa, J. & Vierstra, R. D.: The Arabidopsis thaliana HY1 locus, required for phytochrome-chromophore biosynthesis, encodes a protein related to heme oxygenases. Proc. Natl. Acad. Sci. USA, 96, 6541-6546 (1999)[PubMed]
- Muramoto, T., Kohchi, T., Yokota, A. et al.: The Arabidopsis photomorphogenic mutant hy1 is deficient in phytochrome chromophore biosynthesis as a result of a mutation in a plastid heme oxygenase. Plant Cell, 11, 335-348 (1999)[PubMed]
- Chen, M., Chory, J. & Fankhauser, C.: Light signal transduction in higher plants. Annu. Rev. Genet., 38, 87-117 (2004)[PubMed]
- Holm, M. & Deng, X. W.: Structural organization and interactions of COP1, a light-regulated developmental switch. Plant Mol. Biol., 41, 151-158 (1999)[PubMed]
著者プロフィール
略歴:2002年 北海道大学大学院農学研究科 修了,同年 カナダToronto大学 ポスドク研究員,2006年 理化学研究所植物科学研究センター 基礎科学特別研究員を経て,2009年よりカナダToronto大学 ポスドク研究員.
研究テーマ:ストリゴラクトンのシグナル伝達メカニズムの解析.
考察:研究におけるギャンブルとリスクマネージメント.
© 2010 土屋 雄一朗 Licensed under CC 表示 2.1 日本
