シロオビアゲハのベイツ型擬態の分子機構
西川英輝・藤原晴彦
(東京大学大学院新領域創成科学研究科 先端生命科学専攻遺伝システム革新学分野)
email:藤原晴彦
DOI: 10.7875/first.author.2015.033
A genetic mechanism for female-limited Batesian mimicry in Papilio butterfly.
Hideki Nishikawa, Takuro Iijima, Rei Kajitani, Junichi Yamaguchi, Toshiya Ando, Yutaka Suzuki, Sumio Sugano, Asao Fujiyama, Shunichi Kosugi, Hideki Hirakawa, Satoshi Tabata, Katsuhisa Ozaki, Hiroya Morimoto, Kunio Ihara, Madoka Obara, Hiroshi Hori, Takehiko Itoh, Haruhiko Fujiwara
Nature Genetics, 47, 405-409 (2015)
シロオビアゲハは一部のメスのみが毒をもつチョウベニモンアゲハに擬態する.その擬態型の形質はH遺伝子座に起因することが知られていた.シロオビアゲハの全ゲノム塩基配列解析から,擬態型メスの第25染色体にあるH遺伝子座において逆位の生じていることが判明した.H遺伝子座においては性分化を制御するdsx遺伝子とほか2つの遺伝子が超遺伝子構造をとり,この3つの遺伝子は擬態型メスの翅において高く発現していた.擬態型の染色体のH遺伝子座にコードされるDsx(H) のアミノ酸配列は非擬態型の染色体のh遺伝子座にコードされるDsx(h) とは15か所ほど異なっており,これらの変異が擬態型の形質をひき起こすと考えられた.新規の遺伝子導入法を用いて,dsx(H) 遺伝子が擬態型の紋様の形成を誘導し同時に非擬態型の紋様の形成を抑制すること,dsx(h) 遺伝子は紋様の形成に関与しないことが示された.逆位領域では相同組換えが抑制され,H遺伝子座の超遺伝子構造は長期にわたり安定に維持されてきたと考えられた.
ダーウィンと同じ時代の生物学者ベイツは,南米で採集したチョウを分類する過程において,無毒なチョウの翅の紋様が系統的には離れた毒をもつチョウと類似したケースが多数あることに気づいた.鳥などの捕食者は,毒をもつチョウのまずい味と毒々しい紋様とを連合学習して記憶し,毒をもつチョウだけでなくそれと似た紋様をもつチョウを避けるようになると考えられる.後年,このような擬態は総称してベイツ型擬態とよばれるようになったが,有毒なサンゴヘビに似せた無毒なミルクヘビなど,動物の全般において多数の例が知られている.
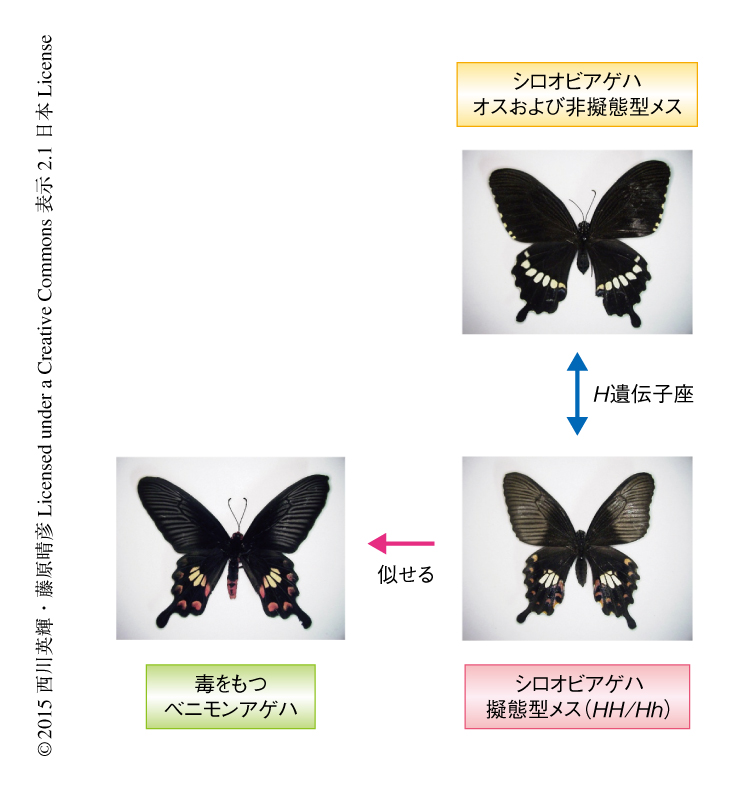
わが国では沖縄など南西諸島に生息するシロオビアゲハは,その名のとおり,後翅に白い帯状の斑紋があるのが普通だが,一部のメスでは後翅の辺縁部に赤いスポット紋様が点在し,また,黄白色の斑紋が中心部にかたよって存在する1)(図1).これは,同じ地域に生息する毒をもつベニモンアゲハに擬態しているといわれている.なぜメスだけが擬態するのか,また,なぜ擬態型と非擬態型の2種類のメスが存在するのかなど,多くの謎が生態学者や動物行動学者の興味をひいてきた.メスのみに限定されたベイツ型擬態はアゲハチョウ科のほかのチョウにもよくみられるが,シロオビアゲハが注目されてきたのは,擬態型メスを生じさせる原因遺伝子が常染色体のH遺伝子座に由来することが遺伝学的に示されていたからである2).擬態型の形質は非擬態型の形質に対し優性で,擬態型メスの遺伝子型はHHあるいはHh,非擬態型メスの遺伝子型はhhである.しかし,オスはすべて非擬態型になる.
古くから,シロオビアゲハの擬態型の原因遺伝子座はじつは単一の遺伝子ではなく,複数の遺伝子が隣接して関与するという仮説が提唱されていた.このような染色体のユニットは超遺伝子(supergene)とよばれ,多岐にわたる複雑な適応形質の原因になっているとみられるが3),これまで,明確にその遺伝子あるいは染色体の構造が示されたのはミューラー型擬態(系統的に離れた毒のある生物が互いに紋様を似せる擬態)をするヘリコニウスという毒をもつチョウだけであった4).しかし,ヘリコニウスの超遺伝子における18個あるそれぞれの遺伝子の機能はわかっていない.一方,シロオビアゲハに関しては,2014年にdoublesex(dsx)遺伝子という性分化を制御する遺伝子がH遺伝子座の超遺伝子の実体であるという報告があったが,その詳細な構造やdsx遺伝子が擬態型の形質にどのように関与するかは不明瞭だった5).
筆者らは,ゲノム塩基配列の解読と機能の解析から,H遺伝子座にはdsx遺伝子のほか2つの遺伝子が含まれ,さらに擬態型の染色体のH遺伝子座に由来するdsx遺伝子のみが擬態型の形質を誘導しうることを示し,超遺伝子の実体および機能をはじめて明らかにした.
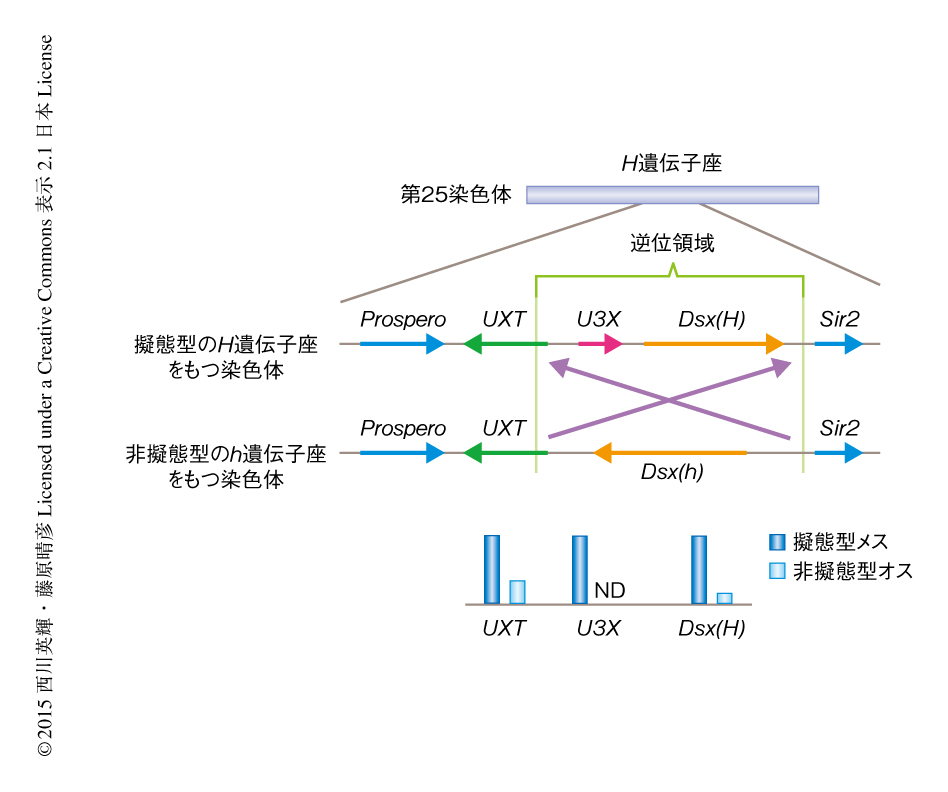
筆者らは,10年近くまえからシロオビアゲハのH遺伝子座を同定しようと試みてきた.シロオビアゲハ亜種のあいだの掛け合わせ実験から,DNA多型を利用して第25染色体の1 Mb以内の領域にH遺伝子座のあることを見い出していたが,さらに今回,この領域にある6個の遺伝子における複数のSNP(1塩基多型)と擬態型の表現型との連関を調べたところ,dsx遺伝子を中心とした200 kb程度の領域に擬態型の原因のあることが示された(図2).
一方,これとはまったく別の角度の解析から,同じ領域がH遺伝子座である可能性が示された.擬態の背景となる遺伝子や染色体の構造を調べるため,シロオビアゲハ,および,参照のためベイツ型擬態をしないナミアゲハの全ゲノム塩基配列を解読した.シロオビアゲハのゲノムは約227 Mb,ナミアゲハのゲノムは約244 Mbであった.シロオビアゲハのゲノム情報を解析したところ,100 kb以上にわたり父方の染色体の配列と母方の染色体の配列が10%以上も異なる領域が15か所みつかった.ゲノム解析に用いたDNAは遺伝子型Hhの擬態型メスの1匹の幼虫から分離したもので,14か所は性染色体に存在した.しかし,1か所だけは常染色体にマップされ,そこは連鎖解析の結果と同じ第25染色体のdsx遺伝子の近傍だったのである.これらの結果から,擬態型のH遺伝子座と非擬態型のh遺伝子座とで構造の大きく異なる2種類の第25染色体が存在する可能性が示唆された.
この領域の構造を詳細に調べてみると,H遺伝子座とh遺伝子座では染色体においてdsx遺伝子の向きが完全に逆になっていた(図2).dsx遺伝子の近傍の遺伝子もあわせて鱗翅目の昆虫のあいだでシンテニーを比較すると,dsx遺伝子の向きはすべてh遺伝子座と同じであった.つまり,h遺伝子座をもつ染色体が本来の染色体で,擬態型のH遺伝子座をもつ染色体はそこから派生したと考えられた.興味深いことに,H遺伝子座とh遺伝子座とのあいだで逆位領域の構造を比較すると,逆向きだけでなく,同じ方向で比較しても配列の相同性はきわめて低かった.つまり,逆位の起こったのちに変異が大量に蓄積し,逆位によりその構造が維持されてきたと考えられた.逆位がいつ起こったのかを類推するため,さらにこの領域を,h遺伝子座をもつシロオビアゲハの染色体とナミアゲハの染色体とで比較すると,H遺伝子座とh遺伝子座の相同性のレベルとそれほど変わらなかった.シロオビアゲハとナミアゲハの分岐年代は約4000万年前と推測されており6),これらの結果から,H遺伝子座の逆位はそれに近い古い年代に生じた可能性が示唆された.
約130 kbに及ぶH遺伝子座の逆位領域を詳細に調べると,dsx遺伝子が丸ごと入っていたほか,2つの遺伝子がみつかった(図2).ひとつは3つのエキソンからなる非コードRNAをコードする遺伝子で,U3X遺伝子と命名した.もうひとつは動物の全般において広く保存されている転写因子UXTをコードする遺伝子であった7).U3X遺伝子はh遺伝子座をもつ染色体には存在せず,また,ほかのゲノム領域にも相同な配列は同定されなかったので,どのように出現したのかは不明であった.一方,逆位の左側の境目はちょうどUXT遺伝子の5’側非翻訳領域の内部に存在しており,UXT遺伝子に対してはH遺伝子座とh遺伝子座とでまったく異なる転写制御がはたらいていると予想された.実際に,UXT遺伝子のmRNAを擬態型メスと非擬態型メスの翅とで比較すると,蛹期初期あるいは蛹期後期のいずれでも擬態型メスにおいて数倍も高く発現していた(図2).また,当然ではあるが,U3X遺伝子は擬態型メスでのみ発現し,非擬態型メスでは発現がみられなかった.UXT遺伝子もU3X遺伝子も擬態型メスの翅において強く発現していたことから,UXT遺伝子およびU3X遺伝子が擬態型の形質に関与している可能性が考えられた.H遺伝子座における逆位の構造とその内部の遺伝子の発現からみると,dsx遺伝子,UXT遺伝子,U3X遺伝子が超遺伝子を形成してベイツ型擬態をひき起こしていると考えられた.これらの領域はH遺伝子座とh遺伝子座とのあいだに逆位が存在するため相同染色体のあいだの組換えが抑制され,その結果,野生集団においてH遺伝子座の超遺伝子の構造は長期にわたり維持されてきたと考えられた.
メスに限定されたベイツ型擬態に性分化を制御するdsx遺伝子が関与するのは当然のように思われるが,dsx遺伝子はどのようにはたらいているのだろうか? Dsxは昆虫から脊椎動物にいたるまで高度に保存された転写因子である.dsx遺伝子からは選択的スプライシングによりメス型mRNAおよびオス型mRNAという異なるアイソフォームが生じ,それぞれがメスおよびオスへの分化を誘導すると考えられる.擬態型メスと非擬態型メスの翅においてdsx遺伝子の発現を調べてみると,擬態型のH遺伝子座に由来するdsx(H) 遺伝子が擬態型メスにおいて強く発現していることがわかった(図2).また,遺伝子型Hhの擬態型メスでは蛹期初期においてdsx(H) 遺伝子がdsx(h) 遺伝子に対し優先的に発現していた.一方で,終齢幼虫後期および蛹期後期においてはdsx(H) 遺伝子よりdsx(h) 遺伝子の発現のほうが高かった.以上の結果から,蛹期初期においてdsx(H) 遺伝子が擬態型の形質の誘導にはたらくと推測された.
dsx(H) 遺伝子とdsx(h) 遺伝子では産物であるタンパク質の機能が異なるのだろうか? 興味深いことに,Dsx(H) とDsx(h) のあいだでは少なくとも15か所のアミノ酸変異がみられた.とくに,5つのアミノ酸残基は鱗翅目の昆虫の全般において保存されているにもかかわらずDsx(H) のみで変異しており,これらのアミノ酸残基がベイツ型擬態の形成に関与している可能性が考えられた.一方,なぜメスにだけ擬態が生じるのかという疑問に答えるため,遺伝子型Hhのオスの翅におけるdsx遺伝子の発現を調べた.驚いたことに,オスの翅ではdsx(h) 遺伝子に比べdsx(H) 遺伝子はほとんど発現してないことが判明した.オスではたとえH遺伝子座をもっていても擬態に関与するdsx(H) 遺伝子は発現せず,擬態型の形質は生じないと考えられた.
以上の結果から,第25染色体において逆位が生じたのち,H遺伝子座に多数の変異が導入され,dsx遺伝子の機能に変化をもたらしただけでなく,オスおよびメスの翅におけるdsx遺伝子の発現パターンが異なるようになったことが示唆された.H遺伝子座は進化の過程でベイツ型擬態を生じさせるように最適化されたと考えられた.
この研究の大きな成果は,dsx遺伝子の機能を実証することに成功したことにある.チョウの翅の紋様を研究している研究グループは世界中に多数あるが,これまで,それを実用的に解析できる系はなかった.その理由のひとつは,鱗翅目の昆虫では2本鎖RNAなど核酸の細胞への取り込みが注射だけでは十分に起こらず,RNAi法などによる機能の解析が困難であったためである.以前に筆者らは,エレクトロポレーション法を利用して個体においてsiRNAや強制発現プラスミドを効率的に導入する系を開発した8,9).今回,蛹の翅に損傷をあたえないよう間接的に電極を接触させるようにくふうして,翅の紋様の解析のために改良した系の開発に成功した.
この新規の遺伝子導入法を用いて,擬態型メスの翅にdsx(H) 遺伝子のみをノックダウンするsiRNAを導入したところ,本来は擬態型になる翅が導入した部分においてのみ非擬態型になった.擬態型メスの後翅に特徴的な赤いスポットや中心部にかたよった黄白色の斑紋の形成が抑制され,同時に,非擬態型メスに特徴的な帯状の白いバンド紋様がみられるようになったのである.重要なことは,擬態型の抑制と非擬態型の誘導とが同時に観察されたことであった.また,蛹化した直後にsiRNAを導入してもこのような変化が起こったことから,少なくとも蛹になった時点でもdsx(H) 遺伝子は擬態型の形質に関与していると考えられた.一方で,dsx(h) 遺伝子に対するsiRNAを導入してもなんの変化も起こらなかった.さらに,dsx(H) 遺伝子に対するsiRNAとdsx(h) 遺伝子に対するsiRNAとを同時に導入した場合は,dsx(H) 遺伝子に対するsiRNAを単独で導入した場合と同じ結果になった.
遺伝子発現の解析とも考えあわせると,これらの結果から,dsx(H) 遺伝子は蛹期初期にメスの翅においてのみ擬態型の紋様の形成を誘導し,同時に,非擬態型の紋様の形成を抑制するはたらきをもつことが示唆された.一方,dsx(h) 遺伝子は翅の紋様の形成には関与していないと考えられた.翅の辺縁部にあるスポット紋様はほかのチョウにもよくみられることから,紋様パターンそのものはdsx(H) 遺伝子のほかの遺伝子により決められ,dsx(H) 遺伝子は擬態型の紋様への着色のスイッチを入れ,同時に,非擬態型の紋様の着色のスイッチを切る役割をはたしていると考えられた.チョウの翅の紋様を変更させる実用的な技術はこれまで知られておらず,今後,このような解析手法は世界中の研究者に使われるようになると期待される.
今回の研究において,シロオビアゲハの第25染色体のH遺伝子座にはdsx遺伝子のほかU3X遺伝子およびUXT遺伝子が含まれ,逆位によりこれらの遺伝子が進化的に固定されてきたことが示された.今後,さらなる機能の解析が必要であるが,U3X遺伝子およびUXT遺伝子も,dsx遺伝子の発現の制御や紋様の形成に関与している可能性が考えられる.古くから,シロオビアゲハのベイツ型擬態の原因は超遺伝子であるとの仮説が提唱されていた10).今回,原因遺伝子および超遺伝子構造がようやく解明されたわけだが,たんにシロオビアゲハの擬態にとどまらず,より広範な超遺伝子の研究のモデルになりうることが期待される.とくに今回,全ゲノム塩基配列解析から逆位の領域を同定した手法は,今後,ほかの生物種においても利用できる可能性が高い.アリの社会性,鳥の羽毛の多型,植物のめしべの形成など,多岐にわたる複雑な適応形質の原因は超遺伝子であるとの報告があいついでいる3).進化遺伝学において古くて重要な概念である超遺伝子についてはほとんど未解明の状態にあったが,今回の研究はその実体の解明のスタート地点として重要な位置づけにあると考えられる.今回の研究の結果が起爆剤となり,ほかの興味深い生物現象の遺伝的な基盤および進化のプロセスが明瞭になっていくだろう.
略歴:2014年 東京大学大学院新領域創成科学研究科 修了,同年より同 特任研究員.
研究テーマ:シロオビアゲハのベイツ型擬態の分子機構.
抱負:今回の知見をもとに,ベイツ型擬態を示すほかの生物種に関しても,その分子機構を明らかにしていきたい.
藤原 晴彦(Haruhiko Fujiwara)
東京大学大学院新領域創成科学研究科 教授.
研究室URL:http://www.idensystem.k.u-tokyo.ac.jp/index.htm
© 2015 西川英輝・藤原晴彦 Licensed under CC 表示 2.1 日本
(東京大学大学院新領域創成科学研究科 先端生命科学専攻遺伝システム革新学分野)
email:藤原晴彦
DOI: 10.7875/first.author.2015.033
A genetic mechanism for female-limited Batesian mimicry in Papilio butterfly.
Hideki Nishikawa, Takuro Iijima, Rei Kajitani, Junichi Yamaguchi, Toshiya Ando, Yutaka Suzuki, Sumio Sugano, Asao Fujiyama, Shunichi Kosugi, Hideki Hirakawa, Satoshi Tabata, Katsuhisa Ozaki, Hiroya Morimoto, Kunio Ihara, Madoka Obara, Hiroshi Hori, Takehiko Itoh, Haruhiko Fujiwara
Nature Genetics, 47, 405-409 (2015)
要 約
シロオビアゲハは一部のメスのみが毒をもつチョウベニモンアゲハに擬態する.その擬態型の形質はH遺伝子座に起因することが知られていた.シロオビアゲハの全ゲノム塩基配列解析から,擬態型メスの第25染色体にあるH遺伝子座において逆位の生じていることが判明した.H遺伝子座においては性分化を制御するdsx遺伝子とほか2つの遺伝子が超遺伝子構造をとり,この3つの遺伝子は擬態型メスの翅において高く発現していた.擬態型の染色体のH遺伝子座にコードされるDsx(H) のアミノ酸配列は非擬態型の染色体のh遺伝子座にコードされるDsx(h) とは15か所ほど異なっており,これらの変異が擬態型の形質をひき起こすと考えられた.新規の遺伝子導入法を用いて,dsx(H) 遺伝子が擬態型の紋様の形成を誘導し同時に非擬態型の紋様の形成を抑制すること,dsx(h) 遺伝子は紋様の形成に関与しないことが示された.逆位領域では相同組換えが抑制され,H遺伝子座の超遺伝子構造は長期にわたり安定に維持されてきたと考えられた.
はじめに
ダーウィンと同じ時代の生物学者ベイツは,南米で採集したチョウを分類する過程において,無毒なチョウの翅の紋様が系統的には離れた毒をもつチョウと類似したケースが多数あることに気づいた.鳥などの捕食者は,毒をもつチョウのまずい味と毒々しい紋様とを連合学習して記憶し,毒をもつチョウだけでなくそれと似た紋様をもつチョウを避けるようになると考えられる.後年,このような擬態は総称してベイツ型擬態とよばれるようになったが,有毒なサンゴヘビに似せた無毒なミルクヘビなど,動物の全般において多数の例が知られている.
わが国では沖縄など南西諸島に生息するシロオビアゲハは,その名のとおり,後翅に白い帯状の斑紋があるのが普通だが,一部のメスでは後翅の辺縁部に赤いスポット紋様が点在し,また,黄白色の斑紋が中心部にかたよって存在する1)(図1).これは,同じ地域に生息する毒をもつベニモンアゲハに擬態しているといわれている.なぜメスだけが擬態するのか,また,なぜ擬態型と非擬態型の2種類のメスが存在するのかなど,多くの謎が生態学者や動物行動学者の興味をひいてきた.メスのみに限定されたベイツ型擬態はアゲハチョウ科のほかのチョウにもよくみられるが,シロオビアゲハが注目されてきたのは,擬態型メスを生じさせる原因遺伝子が常染色体のH遺伝子座に由来することが遺伝学的に示されていたからである2).擬態型の形質は非擬態型の形質に対し優性で,擬態型メスの遺伝子型はHHあるいはHh,非擬態型メスの遺伝子型はhhである.しかし,オスはすべて非擬態型になる.
古くから,シロオビアゲハの擬態型の原因遺伝子座はじつは単一の遺伝子ではなく,複数の遺伝子が隣接して関与するという仮説が提唱されていた.このような染色体のユニットは超遺伝子(supergene)とよばれ,多岐にわたる複雑な適応形質の原因になっているとみられるが3),これまで,明確にその遺伝子あるいは染色体の構造が示されたのはミューラー型擬態(系統的に離れた毒のある生物が互いに紋様を似せる擬態)をするヘリコニウスという毒をもつチョウだけであった4).しかし,ヘリコニウスの超遺伝子における18個あるそれぞれの遺伝子の機能はわかっていない.一方,シロオビアゲハに関しては,2014年にdoublesex(dsx)遺伝子という性分化を制御する遺伝子がH遺伝子座の超遺伝子の実体であるという報告があったが,その詳細な構造やdsx遺伝子が擬態型の形質にどのように関与するかは不明瞭だった5).
筆者らは,ゲノム塩基配列の解読と機能の解析から,H遺伝子座にはdsx遺伝子のほか2つの遺伝子が含まれ,さらに擬態型の染色体のH遺伝子座に由来するdsx遺伝子のみが擬態型の形質を誘導しうることを示し,超遺伝子の実体および機能をはじめて明らかにした.
1.ベイツ型擬態の原因遺伝子座であるH遺伝子座の同定
筆者らは,10年近くまえからシロオビアゲハのH遺伝子座を同定しようと試みてきた.シロオビアゲハ亜種のあいだの掛け合わせ実験から,DNA多型を利用して第25染色体の1 Mb以内の領域にH遺伝子座のあることを見い出していたが,さらに今回,この領域にある6個の遺伝子における複数のSNP(1塩基多型)と擬態型の表現型との連関を調べたところ,dsx遺伝子を中心とした200 kb程度の領域に擬態型の原因のあることが示された(図2).
一方,これとはまったく別の角度の解析から,同じ領域がH遺伝子座である可能性が示された.擬態の背景となる遺伝子や染色体の構造を調べるため,シロオビアゲハ,および,参照のためベイツ型擬態をしないナミアゲハの全ゲノム塩基配列を解読した.シロオビアゲハのゲノムは約227 Mb,ナミアゲハのゲノムは約244 Mbであった.シロオビアゲハのゲノム情報を解析したところ,100 kb以上にわたり父方の染色体の配列と母方の染色体の配列が10%以上も異なる領域が15か所みつかった.ゲノム解析に用いたDNAは遺伝子型Hhの擬態型メスの1匹の幼虫から分離したもので,14か所は性染色体に存在した.しかし,1か所だけは常染色体にマップされ,そこは連鎖解析の結果と同じ第25染色体のdsx遺伝子の近傍だったのである.これらの結果から,擬態型のH遺伝子座と非擬態型のh遺伝子座とで構造の大きく異なる2種類の第25染色体が存在する可能性が示唆された.
2.H遺伝子座の逆位の起源
この領域の構造を詳細に調べてみると,H遺伝子座とh遺伝子座では染色体においてdsx遺伝子の向きが完全に逆になっていた(図2).dsx遺伝子の近傍の遺伝子もあわせて鱗翅目の昆虫のあいだでシンテニーを比較すると,dsx遺伝子の向きはすべてh遺伝子座と同じであった.つまり,h遺伝子座をもつ染色体が本来の染色体で,擬態型のH遺伝子座をもつ染色体はそこから派生したと考えられた.興味深いことに,H遺伝子座とh遺伝子座とのあいだで逆位領域の構造を比較すると,逆向きだけでなく,同じ方向で比較しても配列の相同性はきわめて低かった.つまり,逆位の起こったのちに変異が大量に蓄積し,逆位によりその構造が維持されてきたと考えられた.逆位がいつ起こったのかを類推するため,さらにこの領域を,h遺伝子座をもつシロオビアゲハの染色体とナミアゲハの染色体とで比較すると,H遺伝子座とh遺伝子座の相同性のレベルとそれほど変わらなかった.シロオビアゲハとナミアゲハの分岐年代は約4000万年前と推測されており6),これらの結果から,H遺伝子座の逆位はそれに近い古い年代に生じた可能性が示唆された.
3.H遺伝子座の超遺伝子としての構造
約130 kbに及ぶH遺伝子座の逆位領域を詳細に調べると,dsx遺伝子が丸ごと入っていたほか,2つの遺伝子がみつかった(図2).ひとつは3つのエキソンからなる非コードRNAをコードする遺伝子で,U3X遺伝子と命名した.もうひとつは動物の全般において広く保存されている転写因子UXTをコードする遺伝子であった7).U3X遺伝子はh遺伝子座をもつ染色体には存在せず,また,ほかのゲノム領域にも相同な配列は同定されなかったので,どのように出現したのかは不明であった.一方,逆位の左側の境目はちょうどUXT遺伝子の5’側非翻訳領域の内部に存在しており,UXT遺伝子に対してはH遺伝子座とh遺伝子座とでまったく異なる転写制御がはたらいていると予想された.実際に,UXT遺伝子のmRNAを擬態型メスと非擬態型メスの翅とで比較すると,蛹期初期あるいは蛹期後期のいずれでも擬態型メスにおいて数倍も高く発現していた(図2).また,当然ではあるが,U3X遺伝子は擬態型メスでのみ発現し,非擬態型メスでは発現がみられなかった.UXT遺伝子もU3X遺伝子も擬態型メスの翅において強く発現していたことから,UXT遺伝子およびU3X遺伝子が擬態型の形質に関与している可能性が考えられた.H遺伝子座における逆位の構造とその内部の遺伝子の発現からみると,dsx遺伝子,UXT遺伝子,U3X遺伝子が超遺伝子を形成してベイツ型擬態をひき起こしていると考えられた.これらの領域はH遺伝子座とh遺伝子座とのあいだに逆位が存在するため相同染色体のあいだの組換えが抑制され,その結果,野生集団においてH遺伝子座の超遺伝子の構造は長期にわたり維持されてきたと考えられた.
4.擬態型のH遺伝子座に由来するdsx遺伝子の構造と発現
メスに限定されたベイツ型擬態に性分化を制御するdsx遺伝子が関与するのは当然のように思われるが,dsx遺伝子はどのようにはたらいているのだろうか? Dsxは昆虫から脊椎動物にいたるまで高度に保存された転写因子である.dsx遺伝子からは選択的スプライシングによりメス型mRNAおよびオス型mRNAという異なるアイソフォームが生じ,それぞれがメスおよびオスへの分化を誘導すると考えられる.擬態型メスと非擬態型メスの翅においてdsx遺伝子の発現を調べてみると,擬態型のH遺伝子座に由来するdsx(H) 遺伝子が擬態型メスにおいて強く発現していることがわかった(図2).また,遺伝子型Hhの擬態型メスでは蛹期初期においてdsx(H) 遺伝子がdsx(h) 遺伝子に対し優先的に発現していた.一方で,終齢幼虫後期および蛹期後期においてはdsx(H) 遺伝子よりdsx(h) 遺伝子の発現のほうが高かった.以上の結果から,蛹期初期においてdsx(H) 遺伝子が擬態型の形質の誘導にはたらくと推測された.
dsx(H) 遺伝子とdsx(h) 遺伝子では産物であるタンパク質の機能が異なるのだろうか? 興味深いことに,Dsx(H) とDsx(h) のあいだでは少なくとも15か所のアミノ酸変異がみられた.とくに,5つのアミノ酸残基は鱗翅目の昆虫の全般において保存されているにもかかわらずDsx(H) のみで変異しており,これらのアミノ酸残基がベイツ型擬態の形成に関与している可能性が考えられた.一方,なぜメスにだけ擬態が生じるのかという疑問に答えるため,遺伝子型Hhのオスの翅におけるdsx遺伝子の発現を調べた.驚いたことに,オスの翅ではdsx(h) 遺伝子に比べdsx(H) 遺伝子はほとんど発現してないことが判明した.オスではたとえH遺伝子座をもっていても擬態に関与するdsx(H) 遺伝子は発現せず,擬態型の形質は生じないと考えられた.
以上の結果から,第25染色体において逆位が生じたのち,H遺伝子座に多数の変異が導入され,dsx遺伝子の機能に変化をもたらしただけでなく,オスおよびメスの翅におけるdsx遺伝子の発現パターンが異なるようになったことが示唆された.H遺伝子座は進化の過程でベイツ型擬態を生じさせるように最適化されたと考えられた.
5.新規の遺伝子導入法を用いたdsx遺伝子の機能の解析
この研究の大きな成果は,dsx遺伝子の機能を実証することに成功したことにある.チョウの翅の紋様を研究している研究グループは世界中に多数あるが,これまで,それを実用的に解析できる系はなかった.その理由のひとつは,鱗翅目の昆虫では2本鎖RNAなど核酸の細胞への取り込みが注射だけでは十分に起こらず,RNAi法などによる機能の解析が困難であったためである.以前に筆者らは,エレクトロポレーション法を利用して個体においてsiRNAや強制発現プラスミドを効率的に導入する系を開発した8,9).今回,蛹の翅に損傷をあたえないよう間接的に電極を接触させるようにくふうして,翅の紋様の解析のために改良した系の開発に成功した.
この新規の遺伝子導入法を用いて,擬態型メスの翅にdsx(H) 遺伝子のみをノックダウンするsiRNAを導入したところ,本来は擬態型になる翅が導入した部分においてのみ非擬態型になった.擬態型メスの後翅に特徴的な赤いスポットや中心部にかたよった黄白色の斑紋の形成が抑制され,同時に,非擬態型メスに特徴的な帯状の白いバンド紋様がみられるようになったのである.重要なことは,擬態型の抑制と非擬態型の誘導とが同時に観察されたことであった.また,蛹化した直後にsiRNAを導入してもこのような変化が起こったことから,少なくとも蛹になった時点でもdsx(H) 遺伝子は擬態型の形質に関与していると考えられた.一方で,dsx(h) 遺伝子に対するsiRNAを導入してもなんの変化も起こらなかった.さらに,dsx(H) 遺伝子に対するsiRNAとdsx(h) 遺伝子に対するsiRNAとを同時に導入した場合は,dsx(H) 遺伝子に対するsiRNAを単独で導入した場合と同じ結果になった.
遺伝子発現の解析とも考えあわせると,これらの結果から,dsx(H) 遺伝子は蛹期初期にメスの翅においてのみ擬態型の紋様の形成を誘導し,同時に,非擬態型の紋様の形成を抑制するはたらきをもつことが示唆された.一方,dsx(h) 遺伝子は翅の紋様の形成には関与していないと考えられた.翅の辺縁部にあるスポット紋様はほかのチョウにもよくみられることから,紋様パターンそのものはdsx(H) 遺伝子のほかの遺伝子により決められ,dsx(H) 遺伝子は擬態型の紋様への着色のスイッチを入れ,同時に,非擬態型の紋様の着色のスイッチを切る役割をはたしていると考えられた.チョウの翅の紋様を変更させる実用的な技術はこれまで知られておらず,今後,このような解析手法は世界中の研究者に使われるようになると期待される.
おわりに
今回の研究において,シロオビアゲハの第25染色体のH遺伝子座にはdsx遺伝子のほかU3X遺伝子およびUXT遺伝子が含まれ,逆位によりこれらの遺伝子が進化的に固定されてきたことが示された.今後,さらなる機能の解析が必要であるが,U3X遺伝子およびUXT遺伝子も,dsx遺伝子の発現の制御や紋様の形成に関与している可能性が考えられる.古くから,シロオビアゲハのベイツ型擬態の原因は超遺伝子であるとの仮説が提唱されていた10).今回,原因遺伝子および超遺伝子構造がようやく解明されたわけだが,たんにシロオビアゲハの擬態にとどまらず,より広範な超遺伝子の研究のモデルになりうることが期待される.とくに今回,全ゲノム塩基配列解析から逆位の領域を同定した手法は,今後,ほかの生物種においても利用できる可能性が高い.アリの社会性,鳥の羽毛の多型,植物のめしべの形成など,多岐にわたる複雑な適応形質の原因は超遺伝子であるとの報告があいついでいる3).進化遺伝学において古くて重要な概念である超遺伝子についてはほとんど未解明の状態にあったが,今回の研究はその実体の解明のスタート地点として重要な位置づけにあると考えられる.今回の研究の結果が起爆剤となり,ほかの興味深い生物現象の遺伝的な基盤および進化のプロセスが明瞭になっていくだろう.
文 献
- Nishikawa, H., Iga, M., Yamaguchi, J. et al.: Molecular basis of wing coloration in a Batesian mimic butterfly, Papilio polytes. Sci. Rep., 3, 3184 (2013)[PubMed]
- Clarke, C. A. & Sheppard, P. M.: The genetics of the mimetic butterfly Papilio polytes L. Phil. Trans. R. Soc. Lond. B., 263, 431-458 (1972)[PubMed]
- Schwander, T., Libbrecht, R. & Keller, L.: Supergenes and complex phenotypes. Curr. Biol., 24, R288-R294 (2014)[PubMed]
- Joron, M., Frezal, L., Jones, R. T. et al.: Chromosomal rearrangements maintain a polymorphic supergene controlling butterfly mimicry. Nature, 477, 203-208 (2011)[PubMed]
- Kunte, K., Zhang, W., Tenger-Trolander, A. et al.: doublesex is a mimicry supergene. Nature, 507, 229-232 (2014)[PubMed]
- Zakharov, E. V., Caterino, M. S. & Sperling, F. A.: Molecular phylogeny, historical biogeography, and divergence time estimates for swallowtail butterflies of the genus Papilio (Lepidoptera: Papilionidae). Syst. Biol., 53, 193-215 (2004)[PubMed]
- Schroer, A., Schneider, S., Ropers, H. et al.: Cloning and characterization of UXT, a novel gene in human Xp11, which is widely and abundantly expressed in tumor tissue. Genomics, 56, 340-343 (1999)[PubMed]
- Ando, T. & Fujiwara, H.: Electroporation-mediated somatic transgenesis for rapid functional analysis in insects. Development, 140, 454-458 (2013)[PubMed]
- Yamaguchi, J., Mizoguchi, T. & Fujiwara, H.: siRNAs induce efficient RNAi response in Bombyx mori embryos. PLoS One, 6, e25469 (2011)[PubMed]
- Fisher, R. A.: The Genetic Theory of Natural Selection. Clarendon Press, Oxford (1930)
著者プロフィール
略歴:2014年 東京大学大学院新領域創成科学研究科 修了,同年より同 特任研究員.
研究テーマ:シロオビアゲハのベイツ型擬態の分子機構.
抱負:今回の知見をもとに,ベイツ型擬態を示すほかの生物種に関しても,その分子機構を明らかにしていきたい.
藤原 晴彦(Haruhiko Fujiwara)
東京大学大学院新領域創成科学研究科 教授.
研究室URL:http://www.idensystem.k.u-tokyo.ac.jp/index.htm
© 2015 西川英輝・藤原晴彦 Licensed under CC 表示 2.1 日本
