網膜において双極細胞の投射野は光の刺激により動的に変化する
浅利 宏紀
(米国Harvard大学Department of Molecular and Cellular Biology)
email:浅利宏紀
DOI: 10.7875/first.author.2014.027
The projective field of retinal bipolar cells and its modulation by visual context.
Hiroki Asari, Markus Meister
Neuron, 81, 641-652 (2014)
感覚神経系の研究は“受容野”の概念を中心に発展してきた.しかし,それと相補的な概念である“投射野”についての知見はほとんど得られていない.そこで,この研究では,網膜の神経回路において視覚情報処理の主経路をなす双極細胞から神経節細胞への投射野を同定かつ解析した.その結果,ギャップ結合やアマクリン細胞を介した多シナプス性の投射により,双極細胞は比較的大きな投射野をもつことがわかった.双極細胞および神経節細胞の異なる細胞型のあいだには,光応答の特性に準じた投射の強度および頻度の勾配はあったものの,暗条件のもとでは多対多の密な投射パターンがみられた.ただし,明条件のもとではアマクリン細胞からの多様な制御をうけて投射パターンはより選択的になり,その選択性は光の刺激の違いに応じて変化した.これらの結果は,投射野の構造や機能は形態学的に予想された単シナプス性の投射によるものよりはるかに複雑で動的であることを示唆した.
網膜は平面二層構造をもつ神経回路であり(図1),光信号を電気信号へと変換し,色,形,動きなど局所的な視覚情報を実時間で並列処理する演算装置である1,2).1層目の網膜外層では視細胞により電気信号へと変換された視覚情報が双極細胞へと伝達され,水平細胞による側抑制をうけて時間的および空間的に前処理される.それにつづく網膜内層では双極細胞から神経節細胞へと視覚情報が伝達され,アマクリン細胞との相互作用により神経節細胞の発火パターンとして視覚情報が符号化される.このような初期段階の視覚情報処理は単純そうにみえてじつはきわめて複雑なものであり,たとえば,網膜においては視覚情報が20種類以上もの特徴に分解され,それぞれが別々に符号化されて脳の視覚中枢へと伝達されることが知られている3,4).
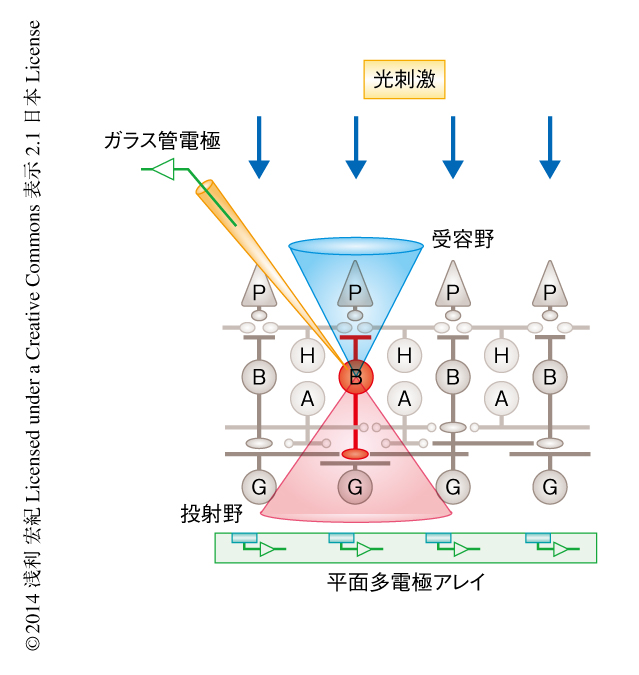
感覚系の神経回路の機能を研究するうえで重要な役割をはたしてきたのが“受容野”の概念である5).受容野とは,適切な刺激が提示されると標的となるニューロンに応答活動をひき起こすような刺激空間領域のことであり,視覚系や聴覚系など感覚系にかかわるさまざまな脳領域において同定されてきた(図1).そして,感覚ニューロン(たとえば,双極細胞)の受容野は,それよりも上流のニューロン(このときは,視細胞)の受容野の組合せにより構成される,という考え方のもと,感覚系の異なる階層における受容野の特性を比較することにより,感覚神経回路の構造や機能の研究が進んできた.
この受容野と対をなす考え方として“投射野”という概念がある6).すなわち,標的となるニューロン(たとえば,双極細胞)の活動がその下流のニューロン(このときは,神経節細胞)の活動に影響を及ぼす範囲のことをそのニューロンの投射野という(図1).さまざまなニューロンによる相互作用から複雑な脳機能が生まれるとすれば,神経回路のはたらきを解明するためには受容野と投射野の両方の知見が必要不可欠である.しかし,投射野の同定には,標的となるニューロンの活動を任意に制御したうえで,その下流にあるすべてのニューロンの応答活動を同時に記録するという実験技術的に高い壁があり,その重要性は議論されてきたものの実験データはほとんど得られていない.
そこで,この研究では,この壁をこえるべく,モデルとなる神経回路として網膜を選んだ.そして,微小ガラス管を用いた細胞内電極法により特定の双極細胞の活動を記録かつ制御する方法と,平面多点電極アレイを用いて複数の神経節細胞の活動を細胞外にて同時に記録する方法を組み合わせることにより,双極細胞から神経節細胞への投射野を同定かつ解析した(図1).なお,この研究は,さきに発表した研究の続報である7)(新着論文レビュー でも掲載).さきの論文では,個々の双極細胞からの出力信号の特性に注目することにより,双極細胞からの出力信号は多種多様な収束パターンおよび発散パターンを示すこと,および,双極細胞から神経節細胞への情報伝達はアマクリン細胞による選択的な制御をうけることを報告した.この研究では,細胞型の別にみた機能的な結合関係に注目し細胞集団レベルでの解析を試みた.
双極細胞は興奮性のニューロンであり,脱分極させるとシナプス結合している神経節細胞の発火頻度は上昇する7).しかし,投射野とは多シナプス性の間接的な情報伝達をも含めた機能的な結合関係を記述するものである.そこで,光の刺激を提示しない定常状態において双極細胞の活動が神経節細胞の活動にあたえる影響の及ぶ範囲とその極性について調べた.その結果,興奮性の投射については,その強度や頻度は距離が大きくなるほど減少することがわかった(図2a).しかし,その範囲は驚くほど広く,双極細胞の投射野はそれ自体の受容野のみならず,神経節細胞の受容野よりも平均して大きかった.一方,抑制性の投射については,興奮性の投射に比べ強度や頻度は低いものの,そのピーク値は距離が少し離れたところにあった.これは,双極細胞の投射野が受容野と同様に“中心周辺拮抗型”をしていることを示唆した.

また,薬理学的な手法を用いることにより,興奮性の投射はおもにギャップ結合により強化されており,抑制性の投射はアマクリン細胞に依存的であることがわかった.これは,ギャップ結合やアマクリン細胞を介した多シナプス性の投射が双極細胞の投射野を構成する要素として無視できないことを意味した.
網膜は視覚情報をいくつもの特徴に分けて並列処理している3).このような並列化は双極細胞の段階からはじまり,神経節細胞へとひき継がれ複雑化していく.そこで,双極細胞および神経節細胞をそれぞれ受容野の構造や光応答の特性にもとづき分類し,これらの細胞型の別に双極細胞の投射野を比較した.その結果,機能的にはどの型の双極細胞もほぼすべての型の神経節細胞へと投射していることがわかった(図2b).形態学的には,おのおのの双極細胞の型がそれぞれ異なる組合せの神経節細胞の型とシナプス結合していることが知られており2),この結果は予想外であった.しかし,暗条件のもとでは光の検出力を高めるため網膜の神経回路は信号の増幅率の高い状態にあることも知られている4).したがって,双極細胞からの多シナプス性の投射も顕在化しやすくなり,このような密な投射パターンが検出されたと考えられた.ただし,異なる細胞型のあいだの投射の強度および頻度には光応答の特性に準じた勾配がみられた.すなわち,たとえば,光が消えたときに応答するオフ型の双極細胞からの投射は,同じオフ型の神経節細胞へとむかうほうがオン型の神経節細胞へとむかう場合よりも強度が強く頻度も高かった.また,ギャップ結合を多くもつオフ型の双極細胞のほうがオン型の双極細胞よりも概して広い投射野をもち,ギャップ結合が網膜内層において横方向の信号の広がりに寄与していることを支持する結果が得られた.
暗条件のもとでは異なる型の双極細胞と神経節細胞とのあいだに多対多の機能的な結合関係がみられたが,明条件のもとではどのような投射パターンが形成されているのであろうか.この問いに答えるため,異なる光刺激を提示しながら双極細胞の投射野を測定した.その結果,明条件のもとでは暗条件のもとと比べ投射野は概して小さくなり,また,より細胞型に依存的になった(図2b).これは,明条件のもとでは双極細胞の投射パターンはより選択的になることを意味した.さらに特筆すべきは,この投射パターンの選択性が光の刺激に依存して変化したことである.すなわち,ある光刺激を提示すると抑制された投射が,また別の光刺激を提示すると増強された投射がみられることがあり,このような投射の動的な制御が細胞型に依存的にみられた.このような網膜内層における多様な情報制御機構は,光の刺激に応じて異なるアマクリン細胞が活性化することにより得られると考えられた7,8).
脳神経系における情報処理を理解するためには,解剖学的な結合関係のみならず,機能的な結合関係についての知見が欠かせない.事実,網膜において双極細胞から神経節細胞への投射野に多シナプス性の投射の寄与が少なからずあったことは,たとえば,電子顕微鏡を用いた神経回路の構造解析だけでは機能の解明にはいたらないことを示唆した.この研究では,実験技術的な理由もありモデルとなる神経回路として網膜を選んだ.しかし,最近の実験技術の発展にはめざましいものがあり,光学的な手法を用いた細胞集団の活動の同時記録や,光遺伝学的な手法によるニューロンの活動制御が可能になってきている9,10).近い将来には,ほかの脳領域においても投射野の同定および解析が進むものと予想される.
感覚神経系では階層ごとにその受容野は大きく複雑になる5).上位の階層での受容野が下位の階層での受容野の組合せにより構成されるとするならば,投射野はその“重み”を記述しているといえる.しかし,双極細胞の受容野と投射野との単純なたたみ込み演算では神経節細胞の受容野を説明できないことは,この研究が示すとおりである.投射野は状況に応じて変化するものであり,情報伝達の動的な制御のさらなる実験的な解析,そして,数理科学的な手法を用いたモデルの研究が進むことにより,感覚神経回路の機能の理解の深まることが期待される.とくに,網膜の神経回路については,異なる型の神経節細胞がどのように形成され,そのそれぞれが視覚情報のどのような特徴を選択的に抽出しているのか,という大問題を解く鍵になるのではないかと期待している.
略歴:2007年 米国Cold Spring Harbor Laboratory博士課程 修了,2008年 米国Harvard大学 ポスドク研究員を経て,2012年より米国California Institute of Technologyポスドク研究員.
研究テーマ:神経科学,神経回路,感覚情報処理.
抱負:さまざまな実験手法や数理的な手法を駆使して神経回路の構造とその機能とのあいだにある因果関係を追究し,脳神経系の動作原理にせまりたい.
© 2014 浅利 宏紀 Licensed under CC 表示 2.1 日本
(米国Harvard大学Department of Molecular and Cellular Biology)
email:浅利宏紀
DOI: 10.7875/first.author.2014.027
The projective field of retinal bipolar cells and its modulation by visual context.
Hiroki Asari, Markus Meister
Neuron, 81, 641-652 (2014)
要 約
感覚神経系の研究は“受容野”の概念を中心に発展してきた.しかし,それと相補的な概念である“投射野”についての知見はほとんど得られていない.そこで,この研究では,網膜の神経回路において視覚情報処理の主経路をなす双極細胞から神経節細胞への投射野を同定かつ解析した.その結果,ギャップ結合やアマクリン細胞を介した多シナプス性の投射により,双極細胞は比較的大きな投射野をもつことがわかった.双極細胞および神経節細胞の異なる細胞型のあいだには,光応答の特性に準じた投射の強度および頻度の勾配はあったものの,暗条件のもとでは多対多の密な投射パターンがみられた.ただし,明条件のもとではアマクリン細胞からの多様な制御をうけて投射パターンはより選択的になり,その選択性は光の刺激の違いに応じて変化した.これらの結果は,投射野の構造や機能は形態学的に予想された単シナプス性の投射によるものよりはるかに複雑で動的であることを示唆した.
はじめに
網膜は平面二層構造をもつ神経回路であり(図1),光信号を電気信号へと変換し,色,形,動きなど局所的な視覚情報を実時間で並列処理する演算装置である1,2).1層目の網膜外層では視細胞により電気信号へと変換された視覚情報が双極細胞へと伝達され,水平細胞による側抑制をうけて時間的および空間的に前処理される.それにつづく網膜内層では双極細胞から神経節細胞へと視覚情報が伝達され,アマクリン細胞との相互作用により神経節細胞の発火パターンとして視覚情報が符号化される.このような初期段階の視覚情報処理は単純そうにみえてじつはきわめて複雑なものであり,たとえば,網膜においては視覚情報が20種類以上もの特徴に分解され,それぞれが別々に符号化されて脳の視覚中枢へと伝達されることが知られている3,4).
感覚系の神経回路の機能を研究するうえで重要な役割をはたしてきたのが“受容野”の概念である5).受容野とは,適切な刺激が提示されると標的となるニューロンに応答活動をひき起こすような刺激空間領域のことであり,視覚系や聴覚系など感覚系にかかわるさまざまな脳領域において同定されてきた(図1).そして,感覚ニューロン(たとえば,双極細胞)の受容野は,それよりも上流のニューロン(このときは,視細胞)の受容野の組合せにより構成される,という考え方のもと,感覚系の異なる階層における受容野の特性を比較することにより,感覚神経回路の構造や機能の研究が進んできた.
この受容野と対をなす考え方として“投射野”という概念がある6).すなわち,標的となるニューロン(たとえば,双極細胞)の活動がその下流のニューロン(このときは,神経節細胞)の活動に影響を及ぼす範囲のことをそのニューロンの投射野という(図1).さまざまなニューロンによる相互作用から複雑な脳機能が生まれるとすれば,神経回路のはたらきを解明するためには受容野と投射野の両方の知見が必要不可欠である.しかし,投射野の同定には,標的となるニューロンの活動を任意に制御したうえで,その下流にあるすべてのニューロンの応答活動を同時に記録するという実験技術的に高い壁があり,その重要性は議論されてきたものの実験データはほとんど得られていない.
そこで,この研究では,この壁をこえるべく,モデルとなる神経回路として網膜を選んだ.そして,微小ガラス管を用いた細胞内電極法により特定の双極細胞の活動を記録かつ制御する方法と,平面多点電極アレイを用いて複数の神経節細胞の活動を細胞外にて同時に記録する方法を組み合わせることにより,双極細胞から神経節細胞への投射野を同定かつ解析した(図1).なお,この研究は,さきに発表した研究の続報である7)(新着論文レビュー でも掲載).さきの論文では,個々の双極細胞からの出力信号の特性に注目することにより,双極細胞からの出力信号は多種多様な収束パターンおよび発散パターンを示すこと,および,双極細胞から神経節細胞への情報伝達はアマクリン細胞による選択的な制御をうけることを報告した.この研究では,細胞型の別にみた機能的な結合関係に注目し細胞集団レベルでの解析を試みた.
1.双極細胞は多シナプス性の投射により比較的大きな投射野をもつ
双極細胞は興奮性のニューロンであり,脱分極させるとシナプス結合している神経節細胞の発火頻度は上昇する7).しかし,投射野とは多シナプス性の間接的な情報伝達をも含めた機能的な結合関係を記述するものである.そこで,光の刺激を提示しない定常状態において双極細胞の活動が神経節細胞の活動にあたえる影響の及ぶ範囲とその極性について調べた.その結果,興奮性の投射については,その強度や頻度は距離が大きくなるほど減少することがわかった(図2a).しかし,その範囲は驚くほど広く,双極細胞の投射野はそれ自体の受容野のみならず,神経節細胞の受容野よりも平均して大きかった.一方,抑制性の投射については,興奮性の投射に比べ強度や頻度は低いものの,そのピーク値は距離が少し離れたところにあった.これは,双極細胞の投射野が受容野と同様に“中心周辺拮抗型”をしていることを示唆した.
また,薬理学的な手法を用いることにより,興奮性の投射はおもにギャップ結合により強化されており,抑制性の投射はアマクリン細胞に依存的であることがわかった.これは,ギャップ結合やアマクリン細胞を介した多シナプス性の投射が双極細胞の投射野を構成する要素として無視できないことを意味した.
2.細胞型の別にみた双極細胞の投射野の比較
網膜は視覚情報をいくつもの特徴に分けて並列処理している3).このような並列化は双極細胞の段階からはじまり,神経節細胞へとひき継がれ複雑化していく.そこで,双極細胞および神経節細胞をそれぞれ受容野の構造や光応答の特性にもとづき分類し,これらの細胞型の別に双極細胞の投射野を比較した.その結果,機能的にはどの型の双極細胞もほぼすべての型の神経節細胞へと投射していることがわかった(図2b).形態学的には,おのおのの双極細胞の型がそれぞれ異なる組合せの神経節細胞の型とシナプス結合していることが知られており2),この結果は予想外であった.しかし,暗条件のもとでは光の検出力を高めるため網膜の神経回路は信号の増幅率の高い状態にあることも知られている4).したがって,双極細胞からの多シナプス性の投射も顕在化しやすくなり,このような密な投射パターンが検出されたと考えられた.ただし,異なる細胞型のあいだの投射の強度および頻度には光応答の特性に準じた勾配がみられた.すなわち,たとえば,光が消えたときに応答するオフ型の双極細胞からの投射は,同じオフ型の神経節細胞へとむかうほうがオン型の神経節細胞へとむかう場合よりも強度が強く頻度も高かった.また,ギャップ結合を多くもつオフ型の双極細胞のほうがオン型の双極細胞よりも概して広い投射野をもち,ギャップ結合が網膜内層において横方向の信号の広がりに寄与していることを支持する結果が得られた.
3.双極細胞の投射野は光の刺激に応じて変化する
暗条件のもとでは異なる型の双極細胞と神経節細胞とのあいだに多対多の機能的な結合関係がみられたが,明条件のもとではどのような投射パターンが形成されているのであろうか.この問いに答えるため,異なる光刺激を提示しながら双極細胞の投射野を測定した.その結果,明条件のもとでは暗条件のもとと比べ投射野は概して小さくなり,また,より細胞型に依存的になった(図2b).これは,明条件のもとでは双極細胞の投射パターンはより選択的になることを意味した.さらに特筆すべきは,この投射パターンの選択性が光の刺激に依存して変化したことである.すなわち,ある光刺激を提示すると抑制された投射が,また別の光刺激を提示すると増強された投射がみられることがあり,このような投射の動的な制御が細胞型に依存的にみられた.このような網膜内層における多様な情報制御機構は,光の刺激に応じて異なるアマクリン細胞が活性化することにより得られると考えられた7,8).
おわりに
脳神経系における情報処理を理解するためには,解剖学的な結合関係のみならず,機能的な結合関係についての知見が欠かせない.事実,網膜において双極細胞から神経節細胞への投射野に多シナプス性の投射の寄与が少なからずあったことは,たとえば,電子顕微鏡を用いた神経回路の構造解析だけでは機能の解明にはいたらないことを示唆した.この研究では,実験技術的な理由もありモデルとなる神経回路として網膜を選んだ.しかし,最近の実験技術の発展にはめざましいものがあり,光学的な手法を用いた細胞集団の活動の同時記録や,光遺伝学的な手法によるニューロンの活動制御が可能になってきている9,10).近い将来には,ほかの脳領域においても投射野の同定および解析が進むものと予想される.
感覚神経系では階層ごとにその受容野は大きく複雑になる5).上位の階層での受容野が下位の階層での受容野の組合せにより構成されるとするならば,投射野はその“重み”を記述しているといえる.しかし,双極細胞の受容野と投射野との単純なたたみ込み演算では神経節細胞の受容野を説明できないことは,この研究が示すとおりである.投射野は状況に応じて変化するものであり,情報伝達の動的な制御のさらなる実験的な解析,そして,数理科学的な手法を用いたモデルの研究が進むことにより,感覚神経回路の機能の理解の深まることが期待される.とくに,網膜の神経回路については,異なる型の神経節細胞がどのように形成され,そのそれぞれが視覚情報のどのような特徴を選択的に抽出しているのか,という大問題を解く鍵になるのではないかと期待している.
文 献
- Dowling, J. E.: The Retina: An Approachable Part of the Brain, Revised Ed. Harvard University Press, Cambridge (2012)
- Masland, R. H.: The neuronal organization of the retina. Neuron, 76, 266-280 (2012)[PubMed]
- Wassle, H.: Parallel processing in the mammalian retina. Nat. Rev. Neurosci., 5, 747-757 (2004)[PubMed]
- Gollisch, T. & Meister, M.: Eye smarter than scientists believed: neural computations in circuits of the retina. Neuron, 65, 150-164 (2010)[PubMed]
- Hubel, D. H. & Wiesel, T. N.: Brain and Visual Perception: The Story of a 25-Year Collaboration. Oxford University Press, New York (2005)
- Sejnowski, T. J.: What are the projective fields of cortical neurons? in 23 Problems in Systems Neuroscience (van Hemmen, J. L. & Sejnowski, T. J., eds.), pp.394-405, Oxford University Press, New York (2006)
- Asari, H. & Meister, M.: Divergence of visual channels in the inner retina. Nat. Neurosci., 15, 1581-1589 (2012)[PubMed] [新着論文レビュー]
- Jadzinsky, P. D. & Baccus, S. A.: Transformation of visual signals by inhibitory interneurons in retinal circuits. Annu. Rev. Neurosci., 36, 403-428 (2013)[PubMed]
- Grienberger, C. & Konnerth, A.: Imaging calcium in neurons. Neuron, 73, 862-885 (2012)[PubMed]
- Packer, A. M., Roska, B. & Hausser, M.: Targeting neurons and photons for optogenetics. Nat. Neurosci., 16, 805-815 (2013)[PubMed]
著者プロフィール
略歴:2007年 米国Cold Spring Harbor Laboratory博士課程 修了,2008年 米国Harvard大学 ポスドク研究員を経て,2012年より米国California Institute of Technologyポスドク研究員.
研究テーマ:神経科学,神経回路,感覚情報処理.
抱負:さまざまな実験手法や数理的な手法を駆使して神経回路の構造とその機能とのあいだにある因果関係を追究し,脳神経系の動作原理にせまりたい.
© 2014 浅利 宏紀 Licensed under CC 表示 2.1 日本
