メダカの雌が見知った雄を配偶者として選択する行動の神経基盤
奥山輝大・竹内秀明
(東京大学大学院理学系研究科 生物科学専攻細胞生理化学研究室)
email:奥山輝大,竹内秀明
DOI: 10.7875/first.author.2014.017
A neural mechanism underlying mating preferences for familiar individuals in medaka fish.
Teruhiro Okuyama, Saori Yokoi, Hideki Abe, Yasuko Isoe, Yuji Suehiro, Haruka Imada, Minoru Tanaka, Takashi Kawasaki, Shunsuke Yuba, Yoshihito Taniguchi, Yasuhiro Kamei, Kataaki Okubo, Atsuko Shimada, Kiyoshi Naruse, Hiroyuki Takeda, Yoshitaka Oka, Takeo Kubo, Hideaki Takeuchi
Science, 343, 91-94 (2014)
社会的な親密性は多くの動物において配偶者を選択する行動に影響をあたえる.しかしながら,その神経基盤については未知な部分が多かった.筆者らは,メダカの雌が配偶者を選択する際,そのまえに視覚的に認識していた見知った雄を記憶していて,その求愛を積極的に受け入れることを見い出した.さらに,配偶者の選択行動に異常を示す変異体を同定し解析した結果,視床下部の外にて神経を修飾する機能をもつと考えられていた終神経の性腺刺激ホルモン放出ホルモン3ニューロンが,選好性を決定するのに中心的な役割を担っていることを明らかにした.終神経の性腺刺激ホルモン放出ホルモン3ニューロンからの性腺刺激ホルモン放出ホルモン3ペプチドの放出を介した自己活性化が,配偶者の選択行動において拒絶から受け入れへのスイッチを担うことが示唆され,この研究により,配偶者の選択行動における意思決定の神経基盤がはじめて明らかになった.
雌が種に特異的な価値基準をもとに配偶の相手を選り好む現象はダーウィンにより“配偶者選択行動”と名づけられ,形態進化の強い駆動力になると考えられている.近年になり,形態的な特徴だけでなく,社会的な親密性も配偶者の選択行動に影響をあたえることがわかってきた1).たとえば,グッピーは新奇な個体を配偶の相手として選択する傾向にあり,一方,一夫一妻制をいとなむプレーリーハタネズミは生活をともにするパートナーを交尾の相手として選択する.さまざまな動物が配偶者の選択行動を示すが,モデル生物を用いた分子遺伝学的な研究はほとんどなく,その神経基盤および分子基盤は明らかになっていない.そこで,この研究では,分子遺伝学におけるモデル生物であるメダカに着目した.メダカは最先端の遺伝学的な手法の利用が可能であり,性周期がわずか1日であるため,毎朝,おのおののペアの配偶行動を解析することができる.この研究では,メダカの雌は見知った雄を配偶の相手として好む傾向にあり,終神経の性腺刺激ホルモン放出ホルモン3ニューロンが配偶の相手を選択する神経基盤において中心的な役割をもつことを示した.
メダカの配偶行動は遺伝的に規定された行動ステップから構成される.雄が雌に対し特徴的な求愛行動を行うと,雌はその求愛を拒絶し,再度,雄が求愛するというやりとりを何回かくり返したのち,数分以内に雌は雄を受け入れて放卵および放精にいたる2).そこで,メダカにおいて配偶の相手との親密性が配偶行動に影響をあたえるかどうかを検討した.前夜から,互いの姿が見えるように隔離したペアと,不透明な壁を用いて見えないように隔離したペアを準備し,翌朝,隔離を解除したのちの配偶行動を観察した.その結果,互いの姿が見えるよう隔離したペアでは,互いの姿が見えないよう隔離したペアと比べ,雌の求愛を受け入れるまでの時間が有意に減少した.また,雄の求愛行動の頻度や求愛を開始するまでの時間に変化はなかった.このことから,おのおののペアにおける親密性は,雄の求愛行動には影響をあたえないが,雌の求愛の受け入れを亢進することがわかった.
さらに,前夜からそばにいた特定の雄に対してのみ,雌の求愛の受け入れの亢進が起こる可能性について検討した.雌に配偶の相手とは異なる雄を見せた場合,互いを見ていた2つのペアを配偶行動の直前に入れ替えた場合,雄に配偶の相手とは異なる雌を見せた場合を比較した.その結果,これらの条件では雌の求愛を受け入れるまでの時間は,互いの姿が見えないよう隔離した場合と同様に長く,雌の求愛の受け入れの亢進は起こらないことがわかった.よって,雌は視覚的に個体を識別し,見知った(視認していた)雄を特異的に配偶の相手として受け入れるというルールがあることが示唆された.さらに,眼球を外科的に切除した雌の個体についてこの配偶行動アッセイを行ったところ,配偶の相手との親密性は雌の求愛の受け入れに影響をあたえなかったため,雌の求愛の受け入れの亢進は視覚情報に依存することが示された.
この配偶行動アッセイ系を用いて,N-エチル-N-ニトロソウレア(ENU)の処理により得た変異体を対象として,雌の求愛の受け入れの亢進に異常を示す変異体を探索したところ,見ていなかった雄に対しても求愛の受け入れの亢進を示す変異体としてCXCR7変異体およびCXCR4変異体を同定した.CXCR7およびCXCR4は共通のリガンドCXCL12/SDFに対する受容体であり,メダカのほかマウスやゼブラフィッシュにおいても細胞移動などの形態形成に協調的に関与することが報告されている3-6).CXCR7遺伝子を特異的にノックダウンするモルフォリノアンチセンスオリゴを顕微注入したところ,変異体と同じ行動異常がみられた.モルフォリノアンチセンスオリゴは顕微注入ののち約1週間で分解されてしまうため,胚発生期におけるCXCR7遺伝子の発現が正常な配偶行動を行うために必要であることが示された.また,これら変異体を用いて,視運動反応試験,群れ行動試験,鏡へのアプローチ試験を行ったところ,視覚能力,運動能力,社会性行動などには異常がないことが確認された.
ゼブラフィッシュやメダカでは胚発生期においてCXCRシグナルが始原生殖細胞の生殖巣への細胞移動に必要であるため3,4),メダカのCXCR7変異体およびCXCR4変異体の雌では生殖巣において始原生殖細胞が減少している.そこで,CXCR7変異体およびCXCR4変異体における行動異常が生殖巣における始原生殖細胞の減少によるのかどうかを検証した.野生型の始原生殖細胞をCXCR7変異体へ細胞移植することにより,CXCR7変異体において始原生殖細胞の数を増加させたが,表現型の回復はみられなかった.一方,始原生殖細胞の生存に必須なDead-End遺伝子をノックダウンするモルフォリノアンチセンスオリゴを顕微注入することにより始原生殖細胞の数を減少させたが,野生型と同様の行動異常を示した.以上より,CXCR7変異体およびCXCR4変異体における行動異常は,生殖巣における始原生殖細胞の減少に依存するものではないことが示された.
マウスおよびゼブラフィッシュではCXCRシグナルが性腺刺激ホルモン放出ホルモン(gonadotropin releasing hormone:GnRH)ニューロンの発生に関与しているため5,6),CXCR7変異体あるいはCXCR4変異体における行動異常は性腺刺激ホルモン放出ホルモンニューロンの発生の異常を原因とする可能性について検討した.メダカには性腺刺激ホルモン放出ホルモンをコードする3つのパラログ遺伝子が存在し,視索前野のGnRH1ニューロンにgnrh1遺伝子,中脳のGnRH2ニューロンにgnrh2遺伝子,終神経のGnRH3ニューロンにgnrh3遺伝子が発現している7-9).それぞれの性腺刺激ホルモン放出ホルモンニューロンを可視化できるトランスジェニック系統に対し,CXCR7遺伝子あるいはCXCR4遺伝子をノックダウンするモルフォリノアンチセンスオリゴを顕微注入したところ,視索前野GnRH1ニューロンおよび中脳GnRH2ニューロンの発生は正常であった一方,終神経GnRH3ニューロンの細胞移動に異常が生じ異所的な軸索投射が生じた.また,遺伝学的にCXCR7を欠損させた系統では同様に終神経GnRH3ニューロンの形態形成に異常が観察された.以上より,終神経GnRH3ニューロンの細胞移動にCXCR7シグナルおよびCXCR4シグナルが必要であることが示された.
これまでの性腺刺激ホルモン放出ホルモンニューロンの研究においては,脳下垂体へと投射し黄体形成ホルモンあるいは卵胞形成ホルモンの放出の制御することにより性成熟や性周期を制御する視索前野GnRH1ニューロンのみが注目されていた.一方で,終神経GnRH3ニューロンは網膜あるいは嗅球から小脳にいたるまで広範囲に軸索を投射するという特徴的な形態をもち,性的な動機づけにかかわると予想されてはいたが,これまで性行動との関連について直接的な証拠はなかった8).そこで,終神経GnRH3ニューロンが雌の求愛の受け入れの制御に関与している可能性を直接的に検討する目的で,レーザーアブレーション法により終神経GnRH3ニューロンを特異的に破壊した.その結果,終神経GnRH3ニューロンを破壊した雌は見ていなかった雄を短時間で受け入れ,これはCXCR7変異体あるいはCXCR4変異体の表現型と同様であった.したがって,終神経GnRH3ニューロンは見ていなかった雄の短時間での受け入れを抑制していることが明らかになった.
雄との親密性が雌の脳における終神経GnRH3ニューロンの神経活動に影響をあたえるかどうか検討した.雌の終神経GnRH3ニューロンの神経活動を,雄を見ていたときと見ていなかったときとで比較した.全脳を摘出したのちに終神経GnRH3ニューロンの神経活動を電気生理学的に記録することが可能であり,この状態では終神経GnRH3ニューロンは一定のリズムで自発的に発火する.ここでは,雄との親密性によりこの自発的な発火の頻度が変化するかどうか検討した.その結果,雄を見ていなかったときは自発発火の頻度は約2 Hzであったのに対し,雄を見ていたときには頻度は4~5 Hzにまで有意に上昇することを見い出した.また,性成熟まえの雌や性成熟してない老齢の雌ではこの自発発火の頻度の上昇は観察されず,頻度は2 Hz以下であった.以上より,自発発火の頻度と雌の求愛の受け入れの程度とのあいだに正の相関関係のあることが示唆された.
終神経GnRH3ニューロンから放出されるGnRH3ペプチドの機能を調べる目的で,TILLING(targeting induced local lesions in genomes)法10) を用いて,10アミノ残基からなるGnRH3ペプチドにおいてSerがProへと置換した変異を同定した.変異を起こしたSerは性腺刺激ホルモン放出ホルモンをコードするパラログ遺伝子のあいだ,および,種間において進化的に保存されている残基であったため,このgnrh3変異体における機能の喪失が期待された.そこで,性腺刺激ホルモン放出ホルモン受容体を発現させた培養細胞において,ルシフェラーゼ法を用いてGnRH受容体1,GnRH受容体2,GnRH受容体3のそれぞれの応答性を調べたところ,いずれも有意に低下していることが示された.配偶行動アッセイを行ったところ,終神経GnRH3ニューロンを破壊した雌とは逆に,このgnrh3変異体の雌は見知った雄を短時間で受け入れることはなかった.よって,GnRH3ペプチドは拒絶から受け入れへのスイッチに必要であることが示唆された.さらに,gnrh3変異体の雌において規則的な自発発火のパターンを記録したところ,発火の頻度は2 Hz以下であり,親密性による頻度の上昇もみられなかった.以上の結果より,GnRH3ペプチドは終神経GnRH3ニューロンにおける視認に依存的な自発発火の頻度の上昇に必要であることが示された.
雌の配偶者の選択行動に親密性が影響をあたえるかどうかを調べる目的で,1匹の雌と2匹の雄を用いて配偶行動アッセイを行った.一方の雄を野生型,もう一方の雄にはGFP遺伝子をホモにもつトランスジェニック系統を用い,子のGFP蛍光を指標として父親を検定した.雌が両方の雄を見ていた場合,雌がトランスジェニック系統の雄を配偶の相手として選択する割合は約5割であった.しかし,トランスジェニック系統の雄を見せて野生型の雄を見せない場合には,トランスジェニック系統の雄を配偶の相手として選択する割合は約9割にまで上昇した.以上より,雌との親密性により雄の適応度は上昇することがわかった.終神経GnRH3ニューロンが配偶者の選択行動に関与するかどうかを検証するため,この1匹の雌と2匹の雄による配偶行動アッセイをCXCR7変異体の雌やgnrh3変異体の雌を用いて行った.その結果,CXCR7変異体の雌もgnrh3変異体の雌も,親密性は配偶の相手の選択には影響をあたえないことがわかった.この結果は,終神経GnRH3ニューロンが配偶者の選択において必須であることを示唆するものであった.
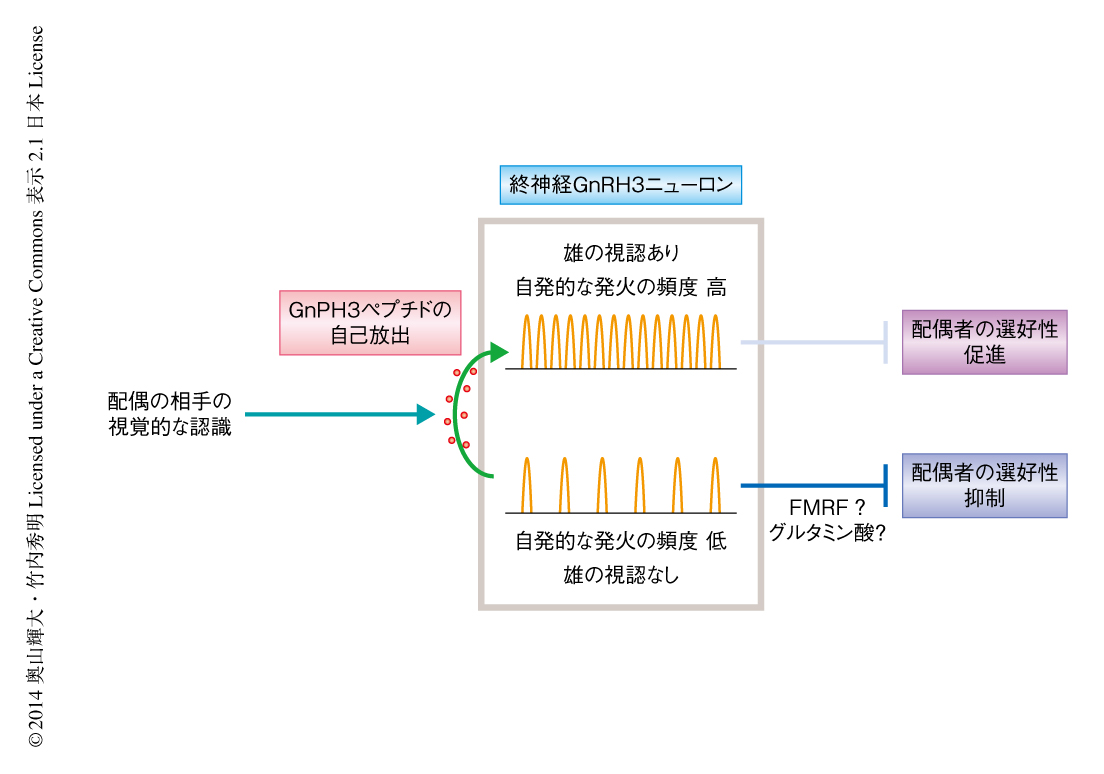
この研究では,個体の識別を介した配偶者の選択行動における意思決定の神経基盤の一端をはじめて解明した.終神経GnRH3ニューロンは求愛の受け入れと拒絶の両方を二相的に制御しており,拒絶から受け入れへのスイッチにはGnRH3ペプチドが必要であることが示された.終神経GnRH3ニューロンは興奮が上昇することによりGnRH3ペプチドを放出することが示唆されているが,その方法として,自己放出経路と,ほかのニューロンの興奮性を変化させるための漏出分泌経路の2つが存在する9,11).魚類の一種ドワーフグラミーにおいてGnRH3ペプチドの暴露により終神経GnRH3ニューロン自体の興奮が上昇することが報告されており12),ペプチドの放出と興奮性の上昇とが正のフィードバックループをつくりだす.gnrh3変異体の雌では見ていた雄に対しても受け入れは亢進せず,終神経GnRH3ニューロンの自発的な発火の頻度は低いままであった.この結果から,雄との親密性が高くなると終神経GnRH3ニューロンの自発的な発火の頻度が徐々に上昇し,ある閾値をこえた時点で正のフィードバックループが機能しはじめ,拒絶から受け入れへのスイッチがオンになるというモデルを提唱した(図1).また,自発的な発火の頻度が低い状態において終神経GnRH3ニューロンが雌の求愛の受け入れを抑制する機構にGnRH3ペプチドは必要がないことが示された.終神経GnRH3ニューロンはグルタミン酸作動性ニューロンであり13),FMRFペプチドを共発現することから,グルタミン酸あるいはFMRFペプチドが雌の求愛の受け入れの抑制に関与しているのかもしれない.終神経GnRH3ニューロンと相同な機能をもつニューロンは哺乳類ではまだ同定されていない.今後,ほかの動物において同様の機能をもつ神経を検索することにより,メダカの研究からヒトを含む脊椎動物において保存された配偶者の選択行動の神経基盤が解明されることが期待される.
略歴:2011年 東京大学大学院理学系研究科 修了,同年 同 研究員,2012年 基礎生物学研究所 研究員を経て,2013年より米国Massachusetts Institute of Technology研究員.
研究テーマ:恋愛感情を制御する脳神経コンポーネントの解析.
抱負:感情と記憶の人工的な操作(書換え)を介して,ヒトの“感情の設計図”を明らかにしていきたい.
竹内 秀明(Hideaki Takeuchi)
東京大学大学院理学系研究科 助教.
© 2014 奥山輝大・竹内秀明 Licensed under CC 表示 2.1 日本
(東京大学大学院理学系研究科 生物科学専攻細胞生理化学研究室)
email:奥山輝大,竹内秀明
DOI: 10.7875/first.author.2014.017
A neural mechanism underlying mating preferences for familiar individuals in medaka fish.
Teruhiro Okuyama, Saori Yokoi, Hideki Abe, Yasuko Isoe, Yuji Suehiro, Haruka Imada, Minoru Tanaka, Takashi Kawasaki, Shunsuke Yuba, Yoshihito Taniguchi, Yasuhiro Kamei, Kataaki Okubo, Atsuko Shimada, Kiyoshi Naruse, Hiroyuki Takeda, Yoshitaka Oka, Takeo Kubo, Hideaki Takeuchi
Science, 343, 91-94 (2014)
要 約
社会的な親密性は多くの動物において配偶者を選択する行動に影響をあたえる.しかしながら,その神経基盤については未知な部分が多かった.筆者らは,メダカの雌が配偶者を選択する際,そのまえに視覚的に認識していた見知った雄を記憶していて,その求愛を積極的に受け入れることを見い出した.さらに,配偶者の選択行動に異常を示す変異体を同定し解析した結果,視床下部の外にて神経を修飾する機能をもつと考えられていた終神経の性腺刺激ホルモン放出ホルモン3ニューロンが,選好性を決定するのに中心的な役割を担っていることを明らかにした.終神経の性腺刺激ホルモン放出ホルモン3ニューロンからの性腺刺激ホルモン放出ホルモン3ペプチドの放出を介した自己活性化が,配偶者の選択行動において拒絶から受け入れへのスイッチを担うことが示唆され,この研究により,配偶者の選択行動における意思決定の神経基盤がはじめて明らかになった.
はじめに
雌が種に特異的な価値基準をもとに配偶の相手を選り好む現象はダーウィンにより“配偶者選択行動”と名づけられ,形態進化の強い駆動力になると考えられている.近年になり,形態的な特徴だけでなく,社会的な親密性も配偶者の選択行動に影響をあたえることがわかってきた1).たとえば,グッピーは新奇な個体を配偶の相手として選択する傾向にあり,一方,一夫一妻制をいとなむプレーリーハタネズミは生活をともにするパートナーを交尾の相手として選択する.さまざまな動物が配偶者の選択行動を示すが,モデル生物を用いた分子遺伝学的な研究はほとんどなく,その神経基盤および分子基盤は明らかになっていない.そこで,この研究では,分子遺伝学におけるモデル生物であるメダカに着目した.メダカは最先端の遺伝学的な手法の利用が可能であり,性周期がわずか1日であるため,毎朝,おのおののペアの配偶行動を解析することができる.この研究では,メダカの雌は見知った雄を配偶の相手として好む傾向にあり,終神経の性腺刺激ホルモン放出ホルモン3ニューロンが配偶の相手を選択する神経基盤において中心的な役割をもつことを示した.
1.メダカの雌は見知った雄を配偶の相手として短時間で受け入れる
メダカの配偶行動は遺伝的に規定された行動ステップから構成される.雄が雌に対し特徴的な求愛行動を行うと,雌はその求愛を拒絶し,再度,雄が求愛するというやりとりを何回かくり返したのち,数分以内に雌は雄を受け入れて放卵および放精にいたる2).そこで,メダカにおいて配偶の相手との親密性が配偶行動に影響をあたえるかどうかを検討した.前夜から,互いの姿が見えるように隔離したペアと,不透明な壁を用いて見えないように隔離したペアを準備し,翌朝,隔離を解除したのちの配偶行動を観察した.その結果,互いの姿が見えるよう隔離したペアでは,互いの姿が見えないよう隔離したペアと比べ,雌の求愛を受け入れるまでの時間が有意に減少した.また,雄の求愛行動の頻度や求愛を開始するまでの時間に変化はなかった.このことから,おのおののペアにおける親密性は,雄の求愛行動には影響をあたえないが,雌の求愛の受け入れを亢進することがわかった.
さらに,前夜からそばにいた特定の雄に対してのみ,雌の求愛の受け入れの亢進が起こる可能性について検討した.雌に配偶の相手とは異なる雄を見せた場合,互いを見ていた2つのペアを配偶行動の直前に入れ替えた場合,雄に配偶の相手とは異なる雌を見せた場合を比較した.その結果,これらの条件では雌の求愛を受け入れるまでの時間は,互いの姿が見えないよう隔離した場合と同様に長く,雌の求愛の受け入れの亢進は起こらないことがわかった.よって,雌は視覚的に個体を識別し,見知った(視認していた)雄を特異的に配偶の相手として受け入れるというルールがあることが示唆された.さらに,眼球を外科的に切除した雌の個体についてこの配偶行動アッセイを行ったところ,配偶の相手との親密性は雌の求愛の受け入れに影響をあたえなかったため,雌の求愛の受け入れの亢進は視覚情報に依存することが示された.
2.CXCR7変異体およびCXCR4変異体の雌は見ていなかった雄を短時間で受け入れる
この配偶行動アッセイ系を用いて,N-エチル-N-ニトロソウレア(ENU)の処理により得た変異体を対象として,雌の求愛の受け入れの亢進に異常を示す変異体を探索したところ,見ていなかった雄に対しても求愛の受け入れの亢進を示す変異体としてCXCR7変異体およびCXCR4変異体を同定した.CXCR7およびCXCR4は共通のリガンドCXCL12/SDFに対する受容体であり,メダカのほかマウスやゼブラフィッシュにおいても細胞移動などの形態形成に協調的に関与することが報告されている3-6).CXCR7遺伝子を特異的にノックダウンするモルフォリノアンチセンスオリゴを顕微注入したところ,変異体と同じ行動異常がみられた.モルフォリノアンチセンスオリゴは顕微注入ののち約1週間で分解されてしまうため,胚発生期におけるCXCR7遺伝子の発現が正常な配偶行動を行うために必要であることが示された.また,これら変異体を用いて,視運動反応試験,群れ行動試験,鏡へのアプローチ試験を行ったところ,視覚能力,運動能力,社会性行動などには異常がないことが確認された.
3.始原生殖細胞の増減は雌の求愛の受け入れには影響しない
ゼブラフィッシュやメダカでは胚発生期においてCXCRシグナルが始原生殖細胞の生殖巣への細胞移動に必要であるため3,4),メダカのCXCR7変異体およびCXCR4変異体の雌では生殖巣において始原生殖細胞が減少している.そこで,CXCR7変異体およびCXCR4変異体における行動異常が生殖巣における始原生殖細胞の減少によるのかどうかを検証した.野生型の始原生殖細胞をCXCR7変異体へ細胞移植することにより,CXCR7変異体において始原生殖細胞の数を増加させたが,表現型の回復はみられなかった.一方,始原生殖細胞の生存に必須なDead-End遺伝子をノックダウンするモルフォリノアンチセンスオリゴを顕微注入することにより始原生殖細胞の数を減少させたが,野生型と同様の行動異常を示した.以上より,CXCR7変異体およびCXCR4変異体における行動異常は,生殖巣における始原生殖細胞の減少に依存するものではないことが示された.
4.終神経の性腺刺激ホルモン放出ホルモンニューロンの形態形成にはCXCRシグナルが必要である
マウスおよびゼブラフィッシュではCXCRシグナルが性腺刺激ホルモン放出ホルモン(gonadotropin releasing hormone:GnRH)ニューロンの発生に関与しているため5,6),CXCR7変異体あるいはCXCR4変異体における行動異常は性腺刺激ホルモン放出ホルモンニューロンの発生の異常を原因とする可能性について検討した.メダカには性腺刺激ホルモン放出ホルモンをコードする3つのパラログ遺伝子が存在し,視索前野のGnRH1ニューロンにgnrh1遺伝子,中脳のGnRH2ニューロンにgnrh2遺伝子,終神経のGnRH3ニューロンにgnrh3遺伝子が発現している7-9).それぞれの性腺刺激ホルモン放出ホルモンニューロンを可視化できるトランスジェニック系統に対し,CXCR7遺伝子あるいはCXCR4遺伝子をノックダウンするモルフォリノアンチセンスオリゴを顕微注入したところ,視索前野GnRH1ニューロンおよび中脳GnRH2ニューロンの発生は正常であった一方,終神経GnRH3ニューロンの細胞移動に異常が生じ異所的な軸索投射が生じた.また,遺伝学的にCXCR7を欠損させた系統では同様に終神経GnRH3ニューロンの形態形成に異常が観察された.以上より,終神経GnRH3ニューロンの細胞移動にCXCR7シグナルおよびCXCR4シグナルが必要であることが示された.
5.終神経GnRH3ニューロンを破壊した雌は見ていなかった雄を短時間で受け入れる
これまでの性腺刺激ホルモン放出ホルモンニューロンの研究においては,脳下垂体へと投射し黄体形成ホルモンあるいは卵胞形成ホルモンの放出の制御することにより性成熟や性周期を制御する視索前野GnRH1ニューロンのみが注目されていた.一方で,終神経GnRH3ニューロンは網膜あるいは嗅球から小脳にいたるまで広範囲に軸索を投射するという特徴的な形態をもち,性的な動機づけにかかわると予想されてはいたが,これまで性行動との関連について直接的な証拠はなかった8).そこで,終神経GnRH3ニューロンが雌の求愛の受け入れの制御に関与している可能性を直接的に検討する目的で,レーザーアブレーション法により終神経GnRH3ニューロンを特異的に破壊した.その結果,終神経GnRH3ニューロンを破壊した雌は見ていなかった雄を短時間で受け入れ,これはCXCR7変異体あるいはCXCR4変異体の表現型と同様であった.したがって,終神経GnRH3ニューロンは見ていなかった雄の短時間での受け入れを抑制していることが明らかになった.
6.雄との親密性は終神経GnRH3ニューロンの規則的な自発発火の頻度を上昇させる
雄との親密性が雌の脳における終神経GnRH3ニューロンの神経活動に影響をあたえるかどうか検討した.雌の終神経GnRH3ニューロンの神経活動を,雄を見ていたときと見ていなかったときとで比較した.全脳を摘出したのちに終神経GnRH3ニューロンの神経活動を電気生理学的に記録することが可能であり,この状態では終神経GnRH3ニューロンは一定のリズムで自発的に発火する.ここでは,雄との親密性によりこの自発的な発火の頻度が変化するかどうか検討した.その結果,雄を見ていなかったときは自発発火の頻度は約2 Hzであったのに対し,雄を見ていたときには頻度は4~5 Hzにまで有意に上昇することを見い出した.また,性成熟まえの雌や性成熟してない老齢の雌ではこの自発発火の頻度の上昇は観察されず,頻度は2 Hz以下であった.以上より,自発発火の頻度と雌の求愛の受け入れの程度とのあいだに正の相関関係のあることが示唆された.
7.GnRH3ペプチドは雌の拒絶から求愛の受け入れへのスイッチに必要である
終神経GnRH3ニューロンから放出されるGnRH3ペプチドの機能を調べる目的で,TILLING(targeting induced local lesions in genomes)法10) を用いて,10アミノ残基からなるGnRH3ペプチドにおいてSerがProへと置換した変異を同定した.変異を起こしたSerは性腺刺激ホルモン放出ホルモンをコードするパラログ遺伝子のあいだ,および,種間において進化的に保存されている残基であったため,このgnrh3変異体における機能の喪失が期待された.そこで,性腺刺激ホルモン放出ホルモン受容体を発現させた培養細胞において,ルシフェラーゼ法を用いてGnRH受容体1,GnRH受容体2,GnRH受容体3のそれぞれの応答性を調べたところ,いずれも有意に低下していることが示された.配偶行動アッセイを行ったところ,終神経GnRH3ニューロンを破壊した雌とは逆に,このgnrh3変異体の雌は見知った雄を短時間で受け入れることはなかった.よって,GnRH3ペプチドは拒絶から受け入れへのスイッチに必要であることが示唆された.さらに,gnrh3変異体の雌において規則的な自発発火のパターンを記録したところ,発火の頻度は2 Hz以下であり,親密性による頻度の上昇もみられなかった.以上の結果より,GnRH3ペプチドは終神経GnRH3ニューロンにおける視認に依存的な自発発火の頻度の上昇に必要であることが示された.
8.雌との親密性により雄の適応度は上昇する
雌の配偶者の選択行動に親密性が影響をあたえるかどうかを調べる目的で,1匹の雌と2匹の雄を用いて配偶行動アッセイを行った.一方の雄を野生型,もう一方の雄にはGFP遺伝子をホモにもつトランスジェニック系統を用い,子のGFP蛍光を指標として父親を検定した.雌が両方の雄を見ていた場合,雌がトランスジェニック系統の雄を配偶の相手として選択する割合は約5割であった.しかし,トランスジェニック系統の雄を見せて野生型の雄を見せない場合には,トランスジェニック系統の雄を配偶の相手として選択する割合は約9割にまで上昇した.以上より,雌との親密性により雄の適応度は上昇することがわかった.終神経GnRH3ニューロンが配偶者の選択行動に関与するかどうかを検証するため,この1匹の雌と2匹の雄による配偶行動アッセイをCXCR7変異体の雌やgnrh3変異体の雌を用いて行った.その結果,CXCR7変異体の雌もgnrh3変異体の雌も,親密性は配偶の相手の選択には影響をあたえないことがわかった.この結果は,終神経GnRH3ニューロンが配偶者の選択において必須であることを示唆するものであった.
おわりに
この研究では,個体の識別を介した配偶者の選択行動における意思決定の神経基盤の一端をはじめて解明した.終神経GnRH3ニューロンは求愛の受け入れと拒絶の両方を二相的に制御しており,拒絶から受け入れへのスイッチにはGnRH3ペプチドが必要であることが示された.終神経GnRH3ニューロンは興奮が上昇することによりGnRH3ペプチドを放出することが示唆されているが,その方法として,自己放出経路と,ほかのニューロンの興奮性を変化させるための漏出分泌経路の2つが存在する9,11).魚類の一種ドワーフグラミーにおいてGnRH3ペプチドの暴露により終神経GnRH3ニューロン自体の興奮が上昇することが報告されており12),ペプチドの放出と興奮性の上昇とが正のフィードバックループをつくりだす.gnrh3変異体の雌では見ていた雄に対しても受け入れは亢進せず,終神経GnRH3ニューロンの自発的な発火の頻度は低いままであった.この結果から,雄との親密性が高くなると終神経GnRH3ニューロンの自発的な発火の頻度が徐々に上昇し,ある閾値をこえた時点で正のフィードバックループが機能しはじめ,拒絶から受け入れへのスイッチがオンになるというモデルを提唱した(図1).また,自発的な発火の頻度が低い状態において終神経GnRH3ニューロンが雌の求愛の受け入れを抑制する機構にGnRH3ペプチドは必要がないことが示された.終神経GnRH3ニューロンはグルタミン酸作動性ニューロンであり13),FMRFペプチドを共発現することから,グルタミン酸あるいはFMRFペプチドが雌の求愛の受け入れの抑制に関与しているのかもしれない.終神経GnRH3ニューロンと相同な機能をもつニューロンは哺乳類ではまだ同定されていない.今後,ほかの動物において同様の機能をもつ神経を検索することにより,メダカの研究からヒトを含む脊椎動物において保存された配偶者の選択行動の神経基盤が解明されることが期待される.
文 献
- Verzijden, M. N., ten Cate, C., Servedio, M. R. et al.: The impact of learning on sexual selection and speciation. Trends Ecol. Evol., 27, 511-519 (2012)[PubMed]
- Ono, Y. & Uematsu, T.: Mating ethogram in Oryzias latipes. J. Fac. Sci. Hokkaido Univ. Ser. 6, Zool., 13, 197-202 (1957)
- Doitsidou, M., Reichman-Fried, M., Stebler, J. et al.: Guidance of primordial germ cell migration by the chemokine SDF-1. Cell, 111, 647-659 (2002)[PubMed]
- Sasado, T., Yasuoka, A., Abe, K. et al.: Distinct contributions of CXCR4b and CXCR7/RDC1 receptor systems in regulation of PGC migration revealed by medaka mutants kazura and yanagi. Dev. Biol., 320, 328-339 (2008)[PubMed]
- Toba, Y., Tiong, J. D., Ma, Q. et al.: CXCR4/SDF-1 system modulates development of GnRH-1 neurons and the olfactory system. Dev. Neurobiol., 68, 487-503 (2008)[PubMed]
- Palevitch, O., Abraham, E., Borodovsky. N. et al.: Cxcl12a-Cxcr4b signaling is important for proper development of the forebrain GnRH system in zebrafish. Gen. Comp. Endocrinol., 165, 262-268 (2010)[PubMed]
- Okubo, K., Sakai, F., Lau, E. L. et al.: Forebrain gonadotropin-releasing hormone neuronal development: insights from transgenic medaka and the relevance to X-linked Kallmann syndrome. Endocrinology, 147, 1076-1084 (2006)[PubMed]
- Oka, Y.: Three types of gonadotrophin-releasing hormone neurones and steroid-sensitive sexually dimorphic kisspeptin neurones in teleosts. J. Neuroendocrinol., 21, 334-338 (2009)[PubMed]
- Karigo, T. & Oka, Y.: Neurobiological study of fish brains gives insights into the nature of gonadotropin-releasing hormone 1-3 neurons. Front. Endocrinol., 4, 177 (2013)[PubMed]
- Ishikawa, T., Kamei, Y., Otozai, S. et al.: High-resolution melting curve analysis for rapid detection of mutations in a Medaka TILLING library. BMC Mol. Biol., 11, 70 (2010)[PubMed]
- Abe, H. & Oka, Y.: Mechanisms of neuromodulation by a nonhypophysiotropic GnRH system controlling motivation of reproductive behavior in the teleost brain. J. Reprod. Dev., 57, 665-674 (2011)[PubMed]
- Abe, H. & Oka, Y.: Modulation of pacemaker activity by salmon gonadotropin-releasing hormone (sGnRH) in terminal nerve (TN)-GnRH neurons. J. Neurophysiol., 83, 3196-3200 (2000)[PubMed]
- Akazome, Y., Kanda, S. & Oka, Y.: Expression of vesicular glutamate transporter-2.1 in medaka terminal nerve gonadotrophin-releasing hormone neurones. J. Neuroendocrinol., 23, 570-576 (2011)[PubMed]
著者プロフィール
略歴:2011年 東京大学大学院理学系研究科 修了,同年 同 研究員,2012年 基礎生物学研究所 研究員を経て,2013年より米国Massachusetts Institute of Technology研究員.
研究テーマ:恋愛感情を制御する脳神経コンポーネントの解析.
抱負:感情と記憶の人工的な操作(書換え)を介して,ヒトの“感情の設計図”を明らかにしていきたい.
竹内 秀明(Hideaki Takeuchi)
東京大学大学院理学系研究科 助教.
© 2014 奥山輝大・竹内秀明 Licensed under CC 表示 2.1 日本
