サイトカイニンの作用の器官に対する特異性は側鎖の修飾により制御される
木羽隆敏・榊原 均
(理化学研究所環境資源科学研究センター 生産機能研究グループ)
email:木羽隆敏,榊原 均
DOI: 10.7875/first.author.2013.160
Side-chain modification of cytokinins controls shoot growth in Arabidopsis.
Takatoshi Kiba, Kentaro Takei, Mikiko Kojima, Hitoshi Sakakibara
Developmental Cell, 27, 452-461 (2013)
多様な器官から構成される植物が個体として生存に最適なバランスを維持するためには,細胞のあいだだけでなく器官のあいだにおいても緊密な情報のやりとりが必要となる.サイトカイニンはそのような情報のやりとりに関与する植物ホルモンである.サイトカイニンの基本骨格はアデニンにイソプレン側鎖のついた構造であり,その側鎖の構造に多様性の存在することは古くから知られていたが,その生物学的な意義は不明であった.筆者らは,サイトカイニンのトランスゼアチン型の側鎖の修飾を担うシトクロムP450モノオキシゲナーゼCYP735Aを同定することにより,側鎖の修飾が不全となった変異株の作出に成功した.この変異株では地上部における成長が遅延するのに対し,CYP735Aを過剰に発現する形質転換株では側鎖修飾は増加し成長の促進がみられた.一方,どちらの変異株においても根の成長は正常であった.側鎖の修飾が不全となった変異株の表現型はトランスゼアチン型サイトカイニンを投与することにより回復したが,側鎖修飾のないサイトカイニンの投与ではまったく効果はなかった.これらの結果から,サイトカイニンの側鎖の修飾は植物の地上部における成長制御において必要であることが明らかになった.
被子植物に代表されるいわゆる高等植物は多様な器官から構成される.光合成を行う葉と水や無機養分の吸収を担う根がその代表的なものである.まったく機能の異なる器官を統合し個体として生存に最適なバランスを維持するためには,細胞のあいだだけでなく器官のあいだにおいても緊密な情報のやりとりが必要となる.サイトカイニンは細胞の分裂および分化の制御から,栄養応答,老化の抑制,イネの着粒数の制御など,植物の成長および発達の全般に深くかかわる植物ホルモンである1,2).サイトカイニンは産生された細胞やその近傍の細胞に対し細胞間のシグナルとして作用するだけでなく,道管および師管を介して長距離を輸送され,器官間のシグナルとしても重要であることが明らかにされつつある3).
サイトカイニンの基本骨格はアデニンにイソプレン側鎖のついた構造であるが,側鎖の構造には多様性が存在し,修飾や二重結合の有無により,基本的にイソペンテニルアデニン型,トランスゼアチン型,シスゼアチン型,ジヒドロゼアチン型の4種類の分子種に分けられる2).モデル植物のシロイヌナズナではイソペンテニルアデニン型とトランスゼアチン型が高い活性をもつ主要なサイトカイニンであるが(図1),複数の分子種の存在することの生物学的な意義は不明であった.そのため,サイトカイニンの作用はおもにその“量”により決定されていると考えられてきた.筆者らは,側鎖の構造の違いの生物学的な意義を明らかにするため,トランスゼアチン型サイトカイニンの合成を担う酵素の遺伝子を同定することによりトランスゼアチン型サイトカイニンを欠損した変異株を作出し詳細に解析した.
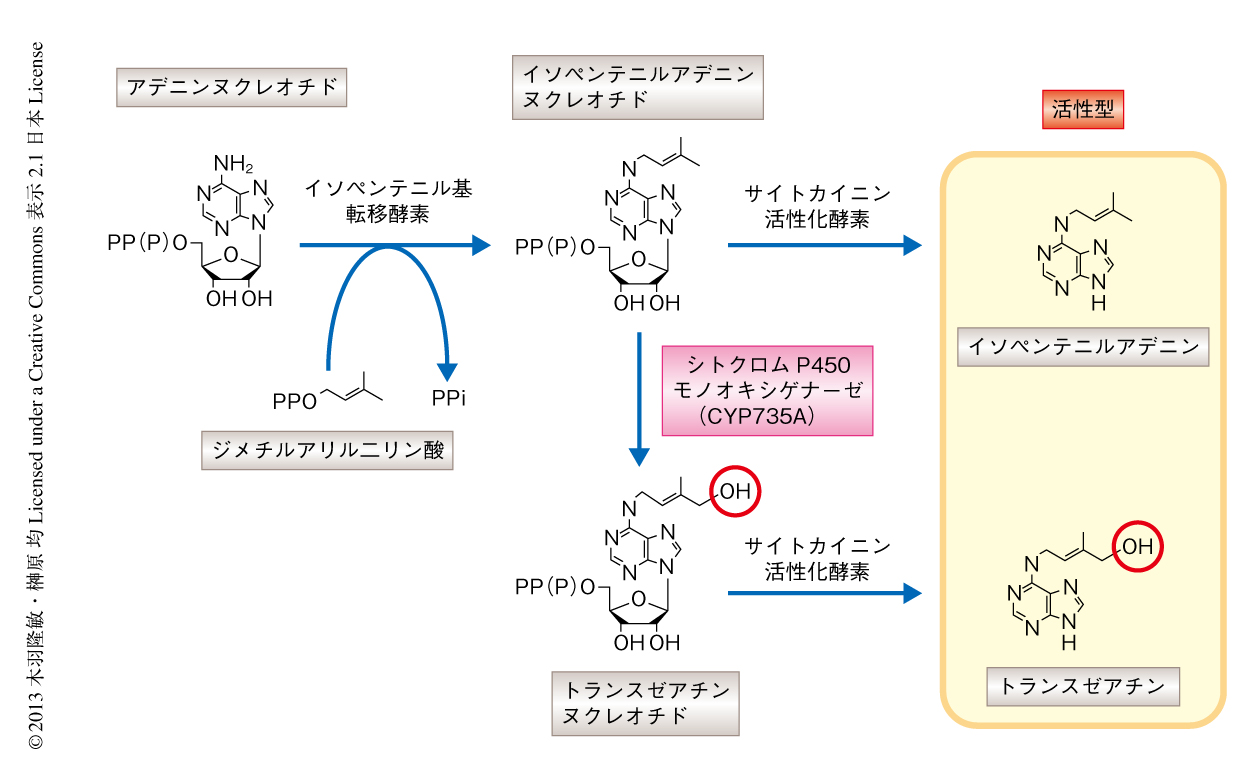
サイトカイニンの生合成の初発反応はイソペンテニル基転移酵素によるイソペンテニルアデニンヌクレオチドの合成である(図1).トランスゼアチン型サイトカイニンはイソペンテニルアデニンヌクレオチドの側鎖の末端のtrans位がシトクロムP450モノオキシゲナーゼCYP735Aにより水酸化されることにより作出されると想定されていた.筆者らは,すでにシロイヌナズナのCYP735AをコードするCYP735A1遺伝子およびCYP735A2遺伝子を同定していたため4),そのノックアウト株を作出し内生するサイトカイニンの量を分析した.単独の変異株,cyp735a1変異株およびcyp735a2変異株では野生型と比べ大きな変化はみられなかった.そこで,cyp735a1 cyp735a2二重変異株について解析したところ,地上部および根においてサイトカイニンの総量に変化はみられなかったが,トランスゼアチン型サイトカイニンはほとんど検出されなかった.この結果から,CYP735A1とCYP735A2がトランスゼアチン型サイトカイニンの合成のための側鎖の修飾に必須の酵素であることが示された.
cyp735a1 cyp735a2二重変異株では,ロゼット葉身,葉柄,花茎の大きさ,花芽数の減少など,地上部における成長の遅延が観察された.ロゼット葉の細胞数の減少および茎頂分裂組織の縮小がみられたことから,茎頂分裂組織の活性の低下がその原因であると考えられた.これらの表現型はサイトカイニンのシグナル伝達や生合成に異常のある変異株に共通してみられるものとよく似ていた5,6).
サイトカイニンのシグナル伝達や生合成に異常のある変異体では地上部の成長は抑制されるのに対し,根の成長は促進されることが知られている5,6).一方,cyp735a1 cyp735a2二重変異株の根は野生株と違いはなかった.サイトカイニン応答性遺伝子の発現を指標にしてサイトカイニンシグナルのレベルを調べても,地上部では低下がみられたのに対し根では有意な差は検出されなかった.
cyp735a1 cyp735a2二重変異株の表現型が側鎖修飾の欠損によりひき起こされているのかどうかを検証するため,サイトカイニンの投与実験を行った.スプレーによりトランスゼアチン型サイトカイニンを投与するとcyp735a1 cyp735a2二重変異株の表現型は回復した.それに対し,10倍も高い濃度のイソペンテニルアデニン型サイトカイニンを投与してもまったく効果はなかった.これらの結果から,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用は異なること,トランスゼアチン型サイトカイニンは地上部における成長の促進に必要であることが示された.
トランスゼアチン型サイトカイニンの生理作用についてさらに知見を得るため,CYP735Aを過剰に発現する形質転換株を作製した.この形質転換株ではサイトカイニンの総量に違いはみられなかったが,トランスゼアチン型サイトカイニンの割合が顕著に増加した.それにともない,地上部における成長が促進されることも明らかになった.一方,根の成長にはまったく影響はみられなかった.これらの結果は,トランスゼアチン型サイトカイニンは地上部における成長を促進する作用をもつこと,根においては必要のないことを裏づけた.
トランスゼアチン型サイトカイニンの合成部位を明らかにするため,CYP735A1遺伝子あるいはCYP735A2遺伝子のプロモーター領域とレポーターとの融合遺伝子を発現させた形質転換株を作製した.CYP735A1遺伝子の発現は非常に弱くレポーターの発現を検出することができなかったが,CYP735A2遺伝子の発現はおもに根において検出された.横断切片を作製してくわしく調べると,CYP735A2遺伝子は維管束の全体において発現していた.
サイトカイニンは長距離を輸送され器官のあいだのシグナルとしてはたらくことが知られている3).トランスゼアチン型サイトカイニンの主要な合成部位が根であるのに対して,変異株の表現型は地上部においてみられたため,トランスゼアチン型サイトカイニンが根から地上部へと輸送されているかどうかを接ぎ木により調べた.野生株の根にcyp735a1 cyp735a2二重変異株の地上部を接ぎ木すると地上部において成長の回復がみられた.これらの結果から,根において合成されたトランスゼアチン型サイトカイニンは地上部へと輸送され,そこでの成長を制御しうることが明らかになった.
シロイヌナズナには3種類のサイトカイニン受容体の遺伝子,AHK2遺伝子,AHK3遺伝子,AHK4/CRE1/WOL遺伝子が存在する.これらにコードされるサイトカイニン受容体はリガンドに対する親和性が異なり,AHK3はイソペンテニルアデニン型サイトカイニンよりもトランスゼアチン型サイトカイニンに対し顕著に高い親和性をもつのに対し,AHK2およびAHK4はどちらに対しても高い親和性を示す7).そこで,このリガンド指向性がトランスゼアチン型サイトカイニンの特異的な作用を規定しているのかどうかを明らかにするため,サイトカイニン受容体遺伝子の二重変異株,ahk2 ahk3二重変異株,ahk2 ahk4二重変異株,ahk3 ahh4二重変異株と,cyp735a1 cyp735a2二重変異株とを組み合わせた四重変異株を作製した.サイトカイニン受容体遺伝子の二重変異株はサイトカイニン受容体を1種類だけもつので,トランスゼアチン型サイトカイニンを欠損するcyp735a1 cyp735a2二重変異株と組み合わせることにより,そのサイトカイニン受容体に対するトランスゼアチン型サイトカイニンの欠損の影響を調べることができる.地上部における成長を指標に解析したところ,どのサイトカイニン受容体をもっていてもトランスゼアチン型サイトカイニンの欠損は成長を遅延させるという結果が得られた.cyp735a1 cyp735a2二重変異株ではサイトカイニンの総量は野生株と変わらない(イソペンテニルアデニン型サイトカイニンは十分に存在する)ことを考慮に入れると,この結果から,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用の違いはサイトカイニン受容体のリガンド指向性のみで決定されているわけではないと解釈された.
この研究により,サイトカイニンの側鎖の修飾はその作用の器官に対する特異性を制御することを明らかにした.これにより,サイトカイニンの“量”にくわえ“質”の制御も個体の統御において重要であることが示された(図2).シトクロムP450モノオキシゲナーゼCYP735Aのオーソログは,藻類,蘚苔類,シダ植物には存在しないことから,側鎖の修飾によるサイトカイニンの“質”の制御は高等植物における高次の器官統御を可能にしたのかもしれない.

一方,この研究は,サイトカイニンの作用の制御について新たな疑問も提示した.トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用の違いを決める,サイトカイニン受容体のリガンド指向性以外の因子は何だろうか? まず,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの組織における局在の異なる可能性が考えられる.確かに,師管液ではイソペンテニルアデニン型サイトカイニンが主要な分子種であるのに対し,道管液ではトランスゼアチン型サイトカイニンがほとんどであると報告されていることから8),これらを偏在させる機構の存在が示唆されている.また,細胞における局在の異なる可能性も考えられる.サイトカイニン受容体は細胞膜だけでなく小胞体膜にも存在するため9),細胞における局在も重要であろう.これらの分子機構を明らかにしていくことにより,高等植物の個体統御のしくみの理解が大きく進展すると期待される.
略歴:2003年 名古屋大学大学院生命農学研究科 修了,2004年 米国Rockefeller大学 ポスドクを経て,2008年より理化学研究所植物科学研究センター(現 環境資源科学研究センター)研究員.
研究テーマ:植物の環境応答.
榊原 均(Hitoshi Sakakibara)
理化学研究所環境資源科学研究センター グループディレクター.
© 2013 木羽隆敏・榊原 均 Licensed under CC 表示 2.1 日本
(理化学研究所環境資源科学研究センター 生産機能研究グループ)
email:木羽隆敏,榊原 均
DOI: 10.7875/first.author.2013.160
Side-chain modification of cytokinins controls shoot growth in Arabidopsis.
Takatoshi Kiba, Kentaro Takei, Mikiko Kojima, Hitoshi Sakakibara
Developmental Cell, 27, 452-461 (2013)
要 約
多様な器官から構成される植物が個体として生存に最適なバランスを維持するためには,細胞のあいだだけでなく器官のあいだにおいても緊密な情報のやりとりが必要となる.サイトカイニンはそのような情報のやりとりに関与する植物ホルモンである.サイトカイニンの基本骨格はアデニンにイソプレン側鎖のついた構造であり,その側鎖の構造に多様性の存在することは古くから知られていたが,その生物学的な意義は不明であった.筆者らは,サイトカイニンのトランスゼアチン型の側鎖の修飾を担うシトクロムP450モノオキシゲナーゼCYP735Aを同定することにより,側鎖の修飾が不全となった変異株の作出に成功した.この変異株では地上部における成長が遅延するのに対し,CYP735Aを過剰に発現する形質転換株では側鎖修飾は増加し成長の促進がみられた.一方,どちらの変異株においても根の成長は正常であった.側鎖の修飾が不全となった変異株の表現型はトランスゼアチン型サイトカイニンを投与することにより回復したが,側鎖修飾のないサイトカイニンの投与ではまったく効果はなかった.これらの結果から,サイトカイニンの側鎖の修飾は植物の地上部における成長制御において必要であることが明らかになった.
はじめに
被子植物に代表されるいわゆる高等植物は多様な器官から構成される.光合成を行う葉と水や無機養分の吸収を担う根がその代表的なものである.まったく機能の異なる器官を統合し個体として生存に最適なバランスを維持するためには,細胞のあいだだけでなく器官のあいだにおいても緊密な情報のやりとりが必要となる.サイトカイニンは細胞の分裂および分化の制御から,栄養応答,老化の抑制,イネの着粒数の制御など,植物の成長および発達の全般に深くかかわる植物ホルモンである1,2).サイトカイニンは産生された細胞やその近傍の細胞に対し細胞間のシグナルとして作用するだけでなく,道管および師管を介して長距離を輸送され,器官間のシグナルとしても重要であることが明らかにされつつある3).
サイトカイニンの基本骨格はアデニンにイソプレン側鎖のついた構造であるが,側鎖の構造には多様性が存在し,修飾や二重結合の有無により,基本的にイソペンテニルアデニン型,トランスゼアチン型,シスゼアチン型,ジヒドロゼアチン型の4種類の分子種に分けられる2).モデル植物のシロイヌナズナではイソペンテニルアデニン型とトランスゼアチン型が高い活性をもつ主要なサイトカイニンであるが(図1),複数の分子種の存在することの生物学的な意義は不明であった.そのため,サイトカイニンの作用はおもにその“量”により決定されていると考えられてきた.筆者らは,側鎖の構造の違いの生物学的な意義を明らかにするため,トランスゼアチン型サイトカイニンの合成を担う酵素の遺伝子を同定することによりトランスゼアチン型サイトカイニンを欠損した変異株を作出し詳細に解析した.
1.cyp735a1 cyp735a2二重変異株はトランスゼアチン型サイトカイニンを欠損する
サイトカイニンの生合成の初発反応はイソペンテニル基転移酵素によるイソペンテニルアデニンヌクレオチドの合成である(図1).トランスゼアチン型サイトカイニンはイソペンテニルアデニンヌクレオチドの側鎖の末端のtrans位がシトクロムP450モノオキシゲナーゼCYP735Aにより水酸化されることにより作出されると想定されていた.筆者らは,すでにシロイヌナズナのCYP735AをコードするCYP735A1遺伝子およびCYP735A2遺伝子を同定していたため4),そのノックアウト株を作出し内生するサイトカイニンの量を分析した.単独の変異株,cyp735a1変異株およびcyp735a2変異株では野生型と比べ大きな変化はみられなかった.そこで,cyp735a1 cyp735a2二重変異株について解析したところ,地上部および根においてサイトカイニンの総量に変化はみられなかったが,トランスゼアチン型サイトカイニンはほとんど検出されなかった.この結果から,CYP735A1とCYP735A2がトランスゼアチン型サイトカイニンの合成のための側鎖の修飾に必須の酵素であることが示された.
2.cyp735a1 cyp735a2二重変異株は地上部において成長の遅延を示すが根は正常である
cyp735a1 cyp735a2二重変異株では,ロゼット葉身,葉柄,花茎の大きさ,花芽数の減少など,地上部における成長の遅延が観察された.ロゼット葉の細胞数の減少および茎頂分裂組織の縮小がみられたことから,茎頂分裂組織の活性の低下がその原因であると考えられた.これらの表現型はサイトカイニンのシグナル伝達や生合成に異常のある変異株に共通してみられるものとよく似ていた5,6).
サイトカイニンのシグナル伝達や生合成に異常のある変異体では地上部の成長は抑制されるのに対し,根の成長は促進されることが知られている5,6).一方,cyp735a1 cyp735a2二重変異株の根は野生株と違いはなかった.サイトカイニン応答性遺伝子の発現を指標にしてサイトカイニンシグナルのレベルを調べても,地上部では低下がみられたのに対し根では有意な差は検出されなかった.
3.cyp735a1 cyp735a2二重変異株の表現型はトランスゼアチン型サイトカイニンの投与により回復する
cyp735a1 cyp735a2二重変異株の表現型が側鎖修飾の欠損によりひき起こされているのかどうかを検証するため,サイトカイニンの投与実験を行った.スプレーによりトランスゼアチン型サイトカイニンを投与するとcyp735a1 cyp735a2二重変異株の表現型は回復した.それに対し,10倍も高い濃度のイソペンテニルアデニン型サイトカイニンを投与してもまったく効果はなかった.これらの結果から,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用は異なること,トランスゼアチン型サイトカイニンは地上部における成長の促進に必要であることが示された.
4.CYP735Aを過剰に発現する形質転換株では地上部における成長が促進される
トランスゼアチン型サイトカイニンの生理作用についてさらに知見を得るため,CYP735Aを過剰に発現する形質転換株を作製した.この形質転換株ではサイトカイニンの総量に違いはみられなかったが,トランスゼアチン型サイトカイニンの割合が顕著に増加した.それにともない,地上部における成長が促進されることも明らかになった.一方,根の成長にはまったく影響はみられなかった.これらの結果は,トランスゼアチン型サイトカイニンは地上部における成長を促進する作用をもつこと,根においては必要のないことを裏づけた.
5.トランスゼアチン型サイトカイニンは根において合成され地上部における成長を制御する
トランスゼアチン型サイトカイニンの合成部位を明らかにするため,CYP735A1遺伝子あるいはCYP735A2遺伝子のプロモーター領域とレポーターとの融合遺伝子を発現させた形質転換株を作製した.CYP735A1遺伝子の発現は非常に弱くレポーターの発現を検出することができなかったが,CYP735A2遺伝子の発現はおもに根において検出された.横断切片を作製してくわしく調べると,CYP735A2遺伝子は維管束の全体において発現していた.
サイトカイニンは長距離を輸送され器官のあいだのシグナルとしてはたらくことが知られている3).トランスゼアチン型サイトカイニンの主要な合成部位が根であるのに対して,変異株の表現型は地上部においてみられたため,トランスゼアチン型サイトカイニンが根から地上部へと輸送されているかどうかを接ぎ木により調べた.野生株の根にcyp735a1 cyp735a2二重変異株の地上部を接ぎ木すると地上部において成長の回復がみられた.これらの結果から,根において合成されたトランスゼアチン型サイトカイニンは地上部へと輸送され,そこでの成長を制御しうることが明らかになった.
6.サイトカイニンの分子種の生理作用の違いは受容体の基質親和性のみでは説明できない
シロイヌナズナには3種類のサイトカイニン受容体の遺伝子,AHK2遺伝子,AHK3遺伝子,AHK4/CRE1/WOL遺伝子が存在する.これらにコードされるサイトカイニン受容体はリガンドに対する親和性が異なり,AHK3はイソペンテニルアデニン型サイトカイニンよりもトランスゼアチン型サイトカイニンに対し顕著に高い親和性をもつのに対し,AHK2およびAHK4はどちらに対しても高い親和性を示す7).そこで,このリガンド指向性がトランスゼアチン型サイトカイニンの特異的な作用を規定しているのかどうかを明らかにするため,サイトカイニン受容体遺伝子の二重変異株,ahk2 ahk3二重変異株,ahk2 ahk4二重変異株,ahk3 ahh4二重変異株と,cyp735a1 cyp735a2二重変異株とを組み合わせた四重変異株を作製した.サイトカイニン受容体遺伝子の二重変異株はサイトカイニン受容体を1種類だけもつので,トランスゼアチン型サイトカイニンを欠損するcyp735a1 cyp735a2二重変異株と組み合わせることにより,そのサイトカイニン受容体に対するトランスゼアチン型サイトカイニンの欠損の影響を調べることができる.地上部における成長を指標に解析したところ,どのサイトカイニン受容体をもっていてもトランスゼアチン型サイトカイニンの欠損は成長を遅延させるという結果が得られた.cyp735a1 cyp735a2二重変異株ではサイトカイニンの総量は野生株と変わらない(イソペンテニルアデニン型サイトカイニンは十分に存在する)ことを考慮に入れると,この結果から,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用の違いはサイトカイニン受容体のリガンド指向性のみで決定されているわけではないと解釈された.
おわりに
この研究により,サイトカイニンの側鎖の修飾はその作用の器官に対する特異性を制御することを明らかにした.これにより,サイトカイニンの“量”にくわえ“質”の制御も個体の統御において重要であることが示された(図2).シトクロムP450モノオキシゲナーゼCYP735Aのオーソログは,藻類,蘚苔類,シダ植物には存在しないことから,側鎖の修飾によるサイトカイニンの“質”の制御は高等植物における高次の器官統御を可能にしたのかもしれない.
一方,この研究は,サイトカイニンの作用の制御について新たな疑問も提示した.トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの生理作用の違いを決める,サイトカイニン受容体のリガンド指向性以外の因子は何だろうか? まず,トランスゼアチン型サイトカイニンとイソペンテニルアデニン型サイトカイニンの組織における局在の異なる可能性が考えられる.確かに,師管液ではイソペンテニルアデニン型サイトカイニンが主要な分子種であるのに対し,道管液ではトランスゼアチン型サイトカイニンがほとんどであると報告されていることから8),これらを偏在させる機構の存在が示唆されている.また,細胞における局在の異なる可能性も考えられる.サイトカイニン受容体は細胞膜だけでなく小胞体膜にも存在するため9),細胞における局在も重要であろう.これらの分子機構を明らかにしていくことにより,高等植物の個体統御のしくみの理解が大きく進展すると期待される.
文 献
- Werner, T. & Schmulling, T.: Cytokinin action in plant development. Curr. Opin. Plant Biol., 12, 527-538 (2009)[PubMed]
- Sakakibara, H.: Cytokinins: activity, biosynthesis, and translocation. Annu. Rev. Plant Biol., 57, 431-449 (2006)[PubMed]
- Kudo, T., Kiba, T. & Sakakibara, H.: Metabolism and long-distance translocation of cytokinins. J. Integr. Plant Biol., 52, 53-60 (2010)[PubMed]
- Takei, T., Yamaya, T., Sakakibara, H.: Arabidopsis CYP735A1 and CYP735A2 encode cytokinin hydroxylases that catalyze the biosynthesis of trans-zeatin. J. Biol. Chem., 279, 41866-41872 (2004)[PubMed]
- Riefler, M., Novak, O., Strnad, M. et al.: Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell, 18, 40-54 (2006)[PubMed]
- Miyawaki, K., Tarkowski, P., Matsumoto-Kitano, M. et al.: Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA, 103, 16598-16603 (2006)[PubMed]
- Stolz, A., Riefler, M., Lomin, S. N. et al.: The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J., 67, 157-168 (2011)[PubMed]
- Hirose, N., Takei, K., Kuroha, T. et al.: Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot., 59, 75-83 (2008)[PubMed]
- Wulfetange, K., Lomin, S. N., Romanov, G. A. et al.: The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum. Plant Physiol., 156, 1808-1818 (2011)[PubMed]
著者プロフィール
略歴:2003年 名古屋大学大学院生命農学研究科 修了,2004年 米国Rockefeller大学 ポスドクを経て,2008年より理化学研究所植物科学研究センター(現 環境資源科学研究センター)研究員.
研究テーマ:植物の環境応答.
榊原 均(Hitoshi Sakakibara)
理化学研究所環境資源科学研究センター グループディレクター.
© 2013 木羽隆敏・榊原 均 Licensed under CC 表示 2.1 日本
