神経幹細胞の長く伸びた形態は核の移動を効率化し混雑を防ぐことにより脳の組織づくりを下支えしている
岡本麻友美・宮田卓樹
(名古屋大学大学院医学系研究科 細胞生物学分野)
email:岡本麻友美,宮田卓樹
DOI: 10.7875/first.author.2013.118
TAG-1-assisted progenitor elongation streamlines nuclear migration to optimize subapical crowding.
Mayumi Okamoto, Takashi Namba, Tomoyasu Shinoda, Takefumi Kondo, Tadashi Watanabe, Yasuhiro Inoue, Kosei Takeuchi, Yukiko Enomoto, Kumiko Ota, Kanako Oda, Yoshino Wada, Ken Sagou, Kanako Saito, Akira Sakakibara, Ayano Kawaguchi, Kazunori Nakajima, Taiji Adachi, Toshihiko Fujimori, Masahiro Ueda, Shigeo Hayashi, Kozo Kaibuchi, Takaki Miyata
Nature Neuroscience, 16, 1556-1566 (2013)
神経幹細胞は脳のさまざまな細胞をつくる役目を担っている.この研究は,神経幹細胞がこの細胞の産生という役目にくわえ,それ自体の細長く伸びた形態を利用して核を適切に移動させ,そののちの脳の3次元的な組織づくりが順調に進むよう下支えしていることを明らかにした. RNAi法によりTAG-1という細胞表面タンパク質をノックダウンすると神経幹細胞は短小化した.この短小化した神経幹細胞は核を十分に移動させることができず,脳原基のもっとも内の脳室に面する付近において過剰な混雑におちいった.こうして過剰な力学的な負荷にさらされた神経幹細胞は,そののち,脳室面という正常な位置から離脱し,本来はニューロンによりしめられるはずの外側の領域に侵入した.その結果,脳の組織の構造はひどく乱れた.今回の成果は,これまで原因のわからなかったヒトの先天性脳形成不全の病因の解明のきっかけにもなると期待される.
胎生期に脳が形成される過程においては,神経幹細胞の分裂により多くの種類のニューロンが必要な数だけつくられること,そして,つくられたニューロンがきちんとならんで組織の構造および神経回路を形成すること,という2つのステップが重要である.神経幹細胞は脳の壁の内のほう(脳室に面する付近)において分裂し,ニューロンは壁の外側に配置されることが知られている1,2).もし,ニューロンが壁の外側に適切にならぶことができないと,神経回路の構造が乱れてんかんなどの症状にもつながる.これまで,ニューロンが壁の外側に適切にならぶことのできない病態の理由として,もっぱらニューロン移動の障害があげられてきた.本来,ニューロンは壁の内側から壁の外側にむけ移動するが,この移動がはたせないためにならびが不十分になるというものである.ヒトの先天性の大脳形成の異常には,このニューロンの移動障害により説明されるものがある3).また,原因となる遺伝子の変異もいくつかわかっている.これらの理由にくわえ,神経幹細胞が本来の内側ではなく外側,すなわち,本来はニューロンだけが存在する場所に入り込んでしまい,そこで分裂してニューロンをでたらめにばらまいてしまうというタイプのニューロンの配置異常も知られている3).ただし,これまでのところ,なぜ神経幹細胞が脳室の付近から離脱してしまうのか,まったくわかっていなかった.
じつは,神経幹細胞は脳室面から0.1 mm程度の範囲で核の反復運動をすることが知られている4-7).分裂はつねに壁の端(脳室に面する場所)で起こり,それ以外のタイミングにある細胞の核は,脳室から離れるか脳室にむかうかのどちらかである.たくさんの神経幹細胞がそれぞれのリズムで反復運動するので,集団としてみると,あちこちでたえずすれちがいが起こっていることになる.こうした動きをとる神経幹細胞の存在する環境は,かなり細胞密度の高い状況にあり空間的な余裕はほとんどない.こうした核の“対向流”の意義について,これまで実験的に問われたことはなかった.一方,どうやって狭いなかでうまくすれちがいがなしとげられるのか,“対向流”が効率よく成立している原理もわかっていなかった.
筆者らは,今回,ニューロンの配置異常と神経幹細胞が狭いなかですれちがうことを結びつける研究を行った.それにあたり,神経幹細胞が脳室から離れるむきに移動しない,すなわち,神経幹細胞の脳室の近くのスペースからの移動が渋滞する状況というのは,脳室の近くの過剰な混雑をうむかもしれないと考えた.そして,この過剰な混雑がある限界をこえたとき,神経幹細胞はそれ以上この場所にいつづけることはできず,緊急退避のような行動をとるのではないかと仮説をたてた.つまり,この過剰な混雑により神経幹細胞の脳室面からの移動が促されるのではないかと考えた.そして,神経幹細胞の核の移動のうち,脳室の近くからの離脱をさまたげる可能性を念頭において,神経幹細胞が細長い形態をしていることに注目し8),脳室の付近にある核にとり,この細長い形態をずっと維持していれば移動に役立つのではないかと考えた.そして,この細長い形態をとらせない実験を行った.
神経幹細胞に長い形態をとらせないために,発生の初期のマウス(ヒトの胎齢で6~7週ごろに相当)において,TAG-1という細胞表面タンパク質をノックダウンした.大脳の壁に存在する神経幹細胞は,本来は壁の外側にまで伸びた長い形態をとっているが,TAG-1のノックダウンにより,神経幹細胞は外側への伸長が保てず,脳室の側にしか細胞体のないきわめて短い形態になった.
TAG-1のノックダウンにより短い形態なった神経幹細胞(胎生12日目,ヒトの7~8週ごろに相当)は,脳室から離れるむきへの核の移動ができなくなった.脳室の近くのスペースでは核の渋滞が起こり,神経幹細胞は過剰な混雑の状況におちいった.
過剰な混雑におちいった神経幹細胞は,翌日(胎生13日目,ヒトの8~9週ごろに相当)までに,脳室に面する位置から離脱し,より外側に無秩序に移動していった.この外側の領域は,本来は移動をおえたニューロンによりしめられるべき場所であるが,神経幹細胞はそこに侵入し異所性に分裂した(図1).
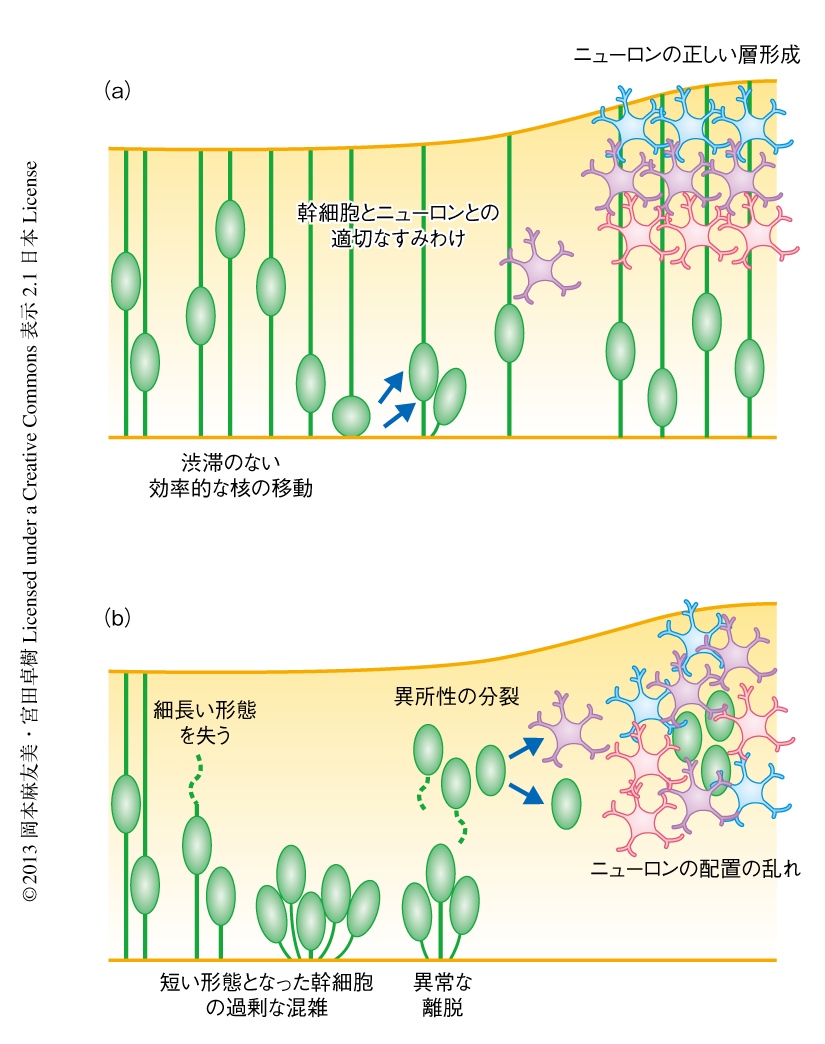
神経幹細胞の異所性の分裂は,胎生15日目(ヒトの12~13週ごろ)からそれ以降にかけてもつづいた.大脳皮質では何日もかけて多くの種類のニューロンが産生されるが,この神経幹細胞の異所性の分裂もニューロンの種類をそろえることには十分に貢献することがわかった.しかし重要なことに,神経幹細胞から本来の場所ではない異所性につくられたニューロンは,正しい層構造ではなく,モザイク状のでたらめな配置パターンを示した.
神経幹細胞が分裂して2つの子細胞が誕生する際,親細胞のもつ細長い形態が片方の子細胞に丸ごと受け継がれることが知られていた8).つまり,生まれた2つのきょうだい細胞には,生まれたときから片方は細長く片方はそうでないという形態的な差がある.これまで,こうした分裂様式の意義は不明であったが,今回,細長い形態を受け継いた子細胞のほうが早く移動することが判明した.つまり,最初にあたえられた形態的な差により,きょうだい細胞のあいだでの核の移動に関する不要な競争がさけられ,狭いトンネル状のスペースにむけ2つの子細胞が順に迅速に移動していくことのできるしくみになっていた.
TAG-1のノックダウンにより短い形態になった神経幹細胞から生じた子細胞は,すべてが細長い形態をとっておらず,本来の効率的に核を移動できる状態から逸脱していた.そして,脳室の近くの過剰な混雑はどんどん悪化した.このように,この研究から,神経幹細胞のとる細長い形態の重要さが明らかになった.
神経幹細胞には,混雑の度合いを感知し本来の場所である脳室面から離脱するという性質があるとわかったことにより,以下のような進展が期待される.
まず,医学的には,先天性脳形成不全の病因の解明に貢献することが期待される.これまで,ニューロンの配置の乱れの理由として,想定はされながらもくわしい理由のわかっていなかった神経幹細胞の異所性の分裂について,今回の成果がきっかけとなりくわしい分子機構の解明にいたることが期待される.過剰な混雑および力学的な負荷を細胞がどう具体的に感知し,どんな細胞内イベントの結果として脳室面からの離脱が起こるのかなど,今回,とらえられた細胞レベルでの現象にもとづき,分子レベルでの解析がはじまり新しい知見が得られるものと期待される.
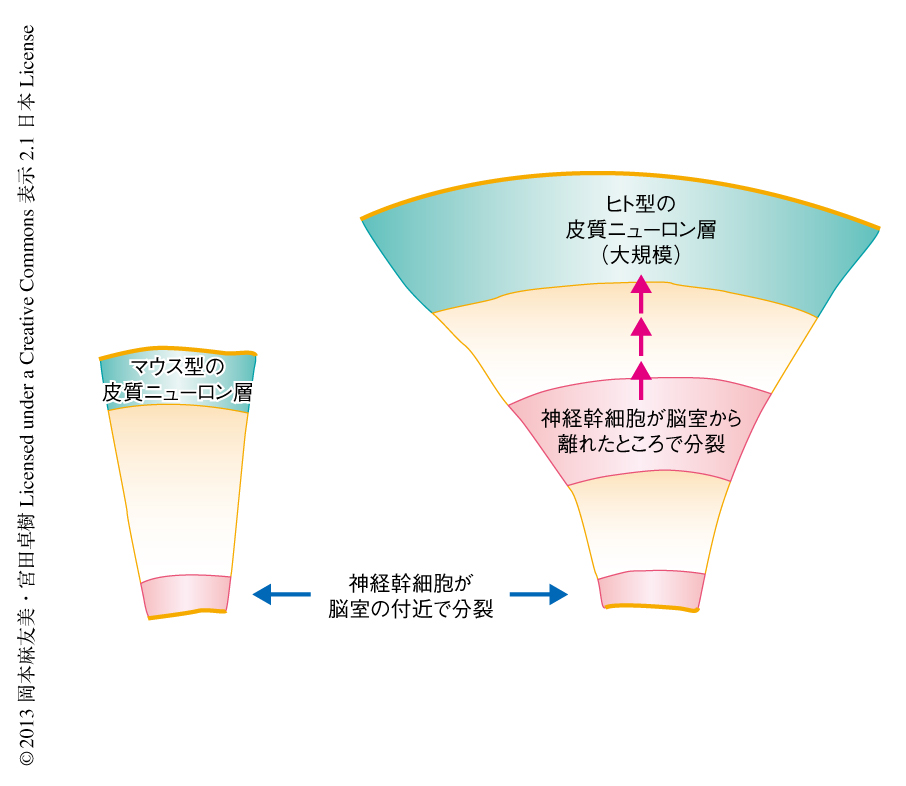
一方,神経科学の分野では,大脳の構造はどのように進化してヒト型にいたったのか,重大な関心がもたれている.じつは,ヒトの大脳では神経幹細胞が脳室面から離れたところへ移動して分裂することが正常なイベントとして高頻度に起こっている2,9,10).最近,これがマウスなどの大脳との大きな違いであることがわかってきた(図2).今回,マウスに対する実験をつうじとらえることのできた,混雑の感知をきっかけとする脳室面からの離脱の分子機構が,進化の過程でなんらかの意味をもった可能性について,今後の研究が期待される.
なお,今回の研究は,すべての細胞の動きの追跡をつうじ定量的かつ統計的なデータを取得する,という新しい研究スタイルをとった.このデータをもとにして,マウス以外の動物での研究や,数理モデルあるいはシミュレーションを用いる研究が進めば,大脳の形成の様式につき動物種をこえて理解が進むものと考えられる.
略歴:2009年 徳島大学大学院先端技術科学教育部博士後期課程 修了,同年 名古屋大学大学院医学系研究科 ポスドクを経て,2010年より同 特任助教.
研究テーマ:大脳の発生の分子機構.
関心事:生物の多様で複雑なかたちがつくられるしくみを知りたい.
宮田 卓樹(Takaki Miyata)
名古屋大学大学院医学系研究科 教授.
研究室URL:http://www.takaki-miyata-lab.org/
© 2013 岡本麻友美・宮田卓樹 Licensed under CC 表示 2.1 日本
(名古屋大学大学院医学系研究科 細胞生物学分野)
email:岡本麻友美,宮田卓樹
DOI: 10.7875/first.author.2013.118
TAG-1-assisted progenitor elongation streamlines nuclear migration to optimize subapical crowding.
Mayumi Okamoto, Takashi Namba, Tomoyasu Shinoda, Takefumi Kondo, Tadashi Watanabe, Yasuhiro Inoue, Kosei Takeuchi, Yukiko Enomoto, Kumiko Ota, Kanako Oda, Yoshino Wada, Ken Sagou, Kanako Saito, Akira Sakakibara, Ayano Kawaguchi, Kazunori Nakajima, Taiji Adachi, Toshihiko Fujimori, Masahiro Ueda, Shigeo Hayashi, Kozo Kaibuchi, Takaki Miyata
Nature Neuroscience, 16, 1556-1566 (2013)
要 約
神経幹細胞は脳のさまざまな細胞をつくる役目を担っている.この研究は,神経幹細胞がこの細胞の産生という役目にくわえ,それ自体の細長く伸びた形態を利用して核を適切に移動させ,そののちの脳の3次元的な組織づくりが順調に進むよう下支えしていることを明らかにした. RNAi法によりTAG-1という細胞表面タンパク質をノックダウンすると神経幹細胞は短小化した.この短小化した神経幹細胞は核を十分に移動させることができず,脳原基のもっとも内の脳室に面する付近において過剰な混雑におちいった.こうして過剰な力学的な負荷にさらされた神経幹細胞は,そののち,脳室面という正常な位置から離脱し,本来はニューロンによりしめられるはずの外側の領域に侵入した.その結果,脳の組織の構造はひどく乱れた.今回の成果は,これまで原因のわからなかったヒトの先天性脳形成不全の病因の解明のきっかけにもなると期待される.
はじめに
胎生期に脳が形成される過程においては,神経幹細胞の分裂により多くの種類のニューロンが必要な数だけつくられること,そして,つくられたニューロンがきちんとならんで組織の構造および神経回路を形成すること,という2つのステップが重要である.神経幹細胞は脳の壁の内のほう(脳室に面する付近)において分裂し,ニューロンは壁の外側に配置されることが知られている1,2).もし,ニューロンが壁の外側に適切にならぶことができないと,神経回路の構造が乱れてんかんなどの症状にもつながる.これまで,ニューロンが壁の外側に適切にならぶことのできない病態の理由として,もっぱらニューロン移動の障害があげられてきた.本来,ニューロンは壁の内側から壁の外側にむけ移動するが,この移動がはたせないためにならびが不十分になるというものである.ヒトの先天性の大脳形成の異常には,このニューロンの移動障害により説明されるものがある3).また,原因となる遺伝子の変異もいくつかわかっている.これらの理由にくわえ,神経幹細胞が本来の内側ではなく外側,すなわち,本来はニューロンだけが存在する場所に入り込んでしまい,そこで分裂してニューロンをでたらめにばらまいてしまうというタイプのニューロンの配置異常も知られている3).ただし,これまでのところ,なぜ神経幹細胞が脳室の付近から離脱してしまうのか,まったくわかっていなかった.
じつは,神経幹細胞は脳室面から0.1 mm程度の範囲で核の反復運動をすることが知られている4-7).分裂はつねに壁の端(脳室に面する場所)で起こり,それ以外のタイミングにある細胞の核は,脳室から離れるか脳室にむかうかのどちらかである.たくさんの神経幹細胞がそれぞれのリズムで反復運動するので,集団としてみると,あちこちでたえずすれちがいが起こっていることになる.こうした動きをとる神経幹細胞の存在する環境は,かなり細胞密度の高い状況にあり空間的な余裕はほとんどない.こうした核の“対向流”の意義について,これまで実験的に問われたことはなかった.一方,どうやって狭いなかでうまくすれちがいがなしとげられるのか,“対向流”が効率よく成立している原理もわかっていなかった.
筆者らは,今回,ニューロンの配置異常と神経幹細胞が狭いなかですれちがうことを結びつける研究を行った.それにあたり,神経幹細胞が脳室から離れるむきに移動しない,すなわち,神経幹細胞の脳室の近くのスペースからの移動が渋滞する状況というのは,脳室の近くの過剰な混雑をうむかもしれないと考えた.そして,この過剰な混雑がある限界をこえたとき,神経幹細胞はそれ以上この場所にいつづけることはできず,緊急退避のような行動をとるのではないかと仮説をたてた.つまり,この過剰な混雑により神経幹細胞の脳室面からの移動が促されるのではないかと考えた.そして,神経幹細胞の核の移動のうち,脳室の近くからの離脱をさまたげる可能性を念頭において,神経幹細胞が細長い形態をしていることに注目し8),脳室の付近にある核にとり,この細長い形態をずっと維持していれば移動に役立つのではないかと考えた.そして,この細長い形態をとらせない実験を行った.
1.神経幹細胞が長い形態をとるにはTAG-1が必要である
神経幹細胞に長い形態をとらせないために,発生の初期のマウス(ヒトの胎齢で6~7週ごろに相当)において,TAG-1という細胞表面タンパク質をノックダウンした.大脳の壁に存在する神経幹細胞は,本来は壁の外側にまで伸びた長い形態をとっているが,TAG-1のノックダウンにより,神経幹細胞は外側への伸長が保てず,脳室の側にしか細胞体のないきわめて短い形態になった.
2.短い形態になった神経幹細胞の核は脳室から離れることができず渋滞した
TAG-1のノックダウンにより短い形態なった神経幹細胞(胎生12日目,ヒトの7~8週ごろに相当)は,脳室から離れるむきへの核の移動ができなくなった.脳室の近くのスペースでは核の渋滞が起こり,神経幹細胞は過剰な混雑の状況におちいった.
3.過剰な混雑におちいった神経幹細胞は脳室面から離脱し本来とは異なる場所で分裂した
過剰な混雑におちいった神経幹細胞は,翌日(胎生13日目,ヒトの8~9週ごろに相当)までに,脳室に面する位置から離脱し,より外側に無秩序に移動していった.この外側の領域は,本来は移動をおえたニューロンによりしめられるべき場所であるが,神経幹細胞はそこに侵入し異所性に分裂した(図1).
4.神経幹細胞の異所性の分裂により大脳皮質の層形成は乱れた
神経幹細胞の異所性の分裂は,胎生15日目(ヒトの12~13週ごろ)からそれ以降にかけてもつづいた.大脳皮質では何日もかけて多くの種類のニューロンが産生されるが,この神経幹細胞の異所性の分裂もニューロンの種類をそろえることには十分に貢献することがわかった.しかし重要なことに,神経幹細胞から本来の場所ではない異所性につくられたニューロンは,正しい層構造ではなく,モザイク状のでたらめな配置パターンを示した.
5.神経幹細胞は細長い形態を子細胞に受け継ぎその動きを助ける
神経幹細胞が分裂して2つの子細胞が誕生する際,親細胞のもつ細長い形態が片方の子細胞に丸ごと受け継がれることが知られていた8).つまり,生まれた2つのきょうだい細胞には,生まれたときから片方は細長く片方はそうでないという形態的な差がある.これまで,こうした分裂様式の意義は不明であったが,今回,細長い形態を受け継いた子細胞のほうが早く移動することが判明した.つまり,最初にあたえられた形態的な差により,きょうだい細胞のあいだでの核の移動に関する不要な競争がさけられ,狭いトンネル状のスペースにむけ2つの子細胞が順に迅速に移動していくことのできるしくみになっていた.
TAG-1のノックダウンにより短い形態になった神経幹細胞から生じた子細胞は,すべてが細長い形態をとっておらず,本来の効率的に核を移動できる状態から逸脱していた.そして,脳室の近くの過剰な混雑はどんどん悪化した.このように,この研究から,神経幹細胞のとる細長い形態の重要さが明らかになった.
おわりに
神経幹細胞には,混雑の度合いを感知し本来の場所である脳室面から離脱するという性質があるとわかったことにより,以下のような進展が期待される.
まず,医学的には,先天性脳形成不全の病因の解明に貢献することが期待される.これまで,ニューロンの配置の乱れの理由として,想定はされながらもくわしい理由のわかっていなかった神経幹細胞の異所性の分裂について,今回の成果がきっかけとなりくわしい分子機構の解明にいたることが期待される.過剰な混雑および力学的な負荷を細胞がどう具体的に感知し,どんな細胞内イベントの結果として脳室面からの離脱が起こるのかなど,今回,とらえられた細胞レベルでの現象にもとづき,分子レベルでの解析がはじまり新しい知見が得られるものと期待される.
一方,神経科学の分野では,大脳の構造はどのように進化してヒト型にいたったのか,重大な関心がもたれている.じつは,ヒトの大脳では神経幹細胞が脳室面から離れたところへ移動して分裂することが正常なイベントとして高頻度に起こっている2,9,10).最近,これがマウスなどの大脳との大きな違いであることがわかってきた(図2).今回,マウスに対する実験をつうじとらえることのできた,混雑の感知をきっかけとする脳室面からの離脱の分子機構が,進化の過程でなんらかの意味をもった可能性について,今後の研究が期待される.
なお,今回の研究は,すべての細胞の動きの追跡をつうじ定量的かつ統計的なデータを取得する,という新しい研究スタイルをとった.このデータをもとにして,マウス以外の動物での研究や,数理モデルあるいはシミュレーションを用いる研究が進めば,大脳の形成の様式につき動物種をこえて理解が進むものと考えられる.
文 献
- Gotz, M. & Huttner, W. B.: The cell biology of neurogensis. Nat. Rev. Mol. Cell Biol., 6, 777-788 (2005)[PubMed]
- Lui, J. H., Hansen, D. V. & Kriegstein, A. R.: Development and evolution of the human neocortex. Cell, 146, 18-36 (2011)[PubMed]
- Barkovich A. J., Guerrini, R., Kuzniecky, R. I. et al.: A developmental and genetic classification for malformations of cortical development: update 2012. Brain, 135, 1348-1369 (2012)[PubMed]
- Sauer, F. C.: Mitosis in the neural tube. J. Comp. Neurol., 62, 377-405 (1935)
- Smart, I. H. M.: The operation of ependymal ‘choke’ in neurogenesis. J. Anat., 99, 941-943 (1965)
- Norden, C., Young, S., Link, B. A. et al.: Actomyosin is the main driver of interkinetic nuclear migration in the retina. Cell, 138, 1195-1208 (2009)[PubMed]
- Taverna, E. & Huttner, W. B.: Neural progenitor nuclei in motion. Neuron, 67, 906-914 (2010)[PubMed]
- Miyata, T., Kawaguchi, A., Okano, H. et al.: Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron, 31, 727-741 (2001)[PubMed]
- Fietz, S. A., Kelava, I., Vogt, J. et al.: OSVZ progenitors of human and ferret neocortex are epithelial-like and expand by integrin signaling. Nat. Neurosci., 13, 690-699 (2010)[PubMed]
- Hansen, D. V., Lui, J. H., Parker, P. R. et al.: Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature, 464, 554-561 (2010)[PubMed]
著者プロフィール
略歴:2009年 徳島大学大学院先端技術科学教育部博士後期課程 修了,同年 名古屋大学大学院医学系研究科 ポスドクを経て,2010年より同 特任助教.
研究テーマ:大脳の発生の分子機構.
関心事:生物の多様で複雑なかたちがつくられるしくみを知りたい.
宮田 卓樹(Takaki Miyata)
名古屋大学大学院医学系研究科 教授.
研究室URL:http://www.takaki-miyata-lab.org/
© 2013 岡本麻友美・宮田卓樹 Licensed under CC 表示 2.1 日本
