2つの雌性生殖細胞による独立の制御が複数の花粉管の誘引を阻止する
丸山大輔・東山哲也
(名古屋大学トランスフォーマティブ生命分子研究所)
email:丸山大輔
DOI: 10.7875/first.author.2013.070
Independent control by each female gamete prevents the attraction of multiple pollen tubes.
Daisuke Maruyama, Yuki Hamamura, Hidenori Takeuchi, Daichi Susaki, Moe Nishimaki, Daisuke Kurihara, Ryushiro D. Kasahara, Tetsuya Higashiyama
Developmental Cell, 25, 317-323 (2013)
被子植物の生殖の過程では,通常,最初に到達した花粉管により重複受精が完了する.受精した胚珠は後続の花粉管が来ないよう制御しているが,そのしくみについては明らかにされていなかった.筆者らは,シロイヌナズナを用いて,卵細胞または中央細胞のどちらかにおける受精の失敗が,2本目以降の花粉管の誘引を停止させる活性の低下をまねくことを示した.同様の表現型は,中央細胞において遺伝子発現を負に制御するFIS-PRC2の機能欠損によっても観察された.FIS-PRC2は胚乳において特異的に機能することから,卵細胞と中央細胞はそれぞれ独自のシグナル伝達経路を発達させてきたものと考えられた.興味深いことに,卵細胞または中央細胞のどちらか一方が受精した胚珠では,その不完全な重複受精を原因として誘引された2本目の花粉管により受精が回復し,その結果,遺伝子型の異なる胚と胚乳をもつ種子が得られた.したがって,卵細胞と中央細胞のそれぞれが受精したかどうかを独自に判断し,2本目の花粉管の誘引をつうじた受精回復システムを構築することにより,生殖における適応度をあげている可能性が示唆された.
鞭毛や繊毛をもたない被子植物の精細胞は自律的な移動ができず,代わりに,精細胞は花粉管という管状の細胞により輸送される.雌しべの柱頭において受粉が起こると,花粉から伸長をはじめた花粉管は互いに競争しながら雌しべの内部へと進んでいく.雌しべの内部には受精ののちに種子となる胚珠がならんでいる.花粉管は受精を完了した胚珠をさけて未受精の胚珠にむかうため,結果として,ほぼすべての胚珠が花粉管と一対一の関係で受精する1)(図1).この2本目の花粉管をしりぞけるしくみは多花粉管拒否とよばれ2),古くからその存在は知られていたものの,どのような分子機構によるものなのかは不明であった.

そのようななか,2012年,筆者らのグループおよび米国のグループは,シロイヌナズナを用い,重複受精が多花粉管拒否の開始の鍵をにぎることを示した2,3).胚珠には花粉管を誘引する物質を分泌する助細胞が2つ存在する.到達した花粉管は,片方の助細胞の崩壊とともに,2個の精細胞を放出して卵細胞と中央細胞という雌側の2つの配偶子の受精を完了する.この被子植物に独特の受精現象が重複受精である(図1,植物の受精のしくみについては,東山 哲也, 領域融合レビュー, 1, e007, 2012 も参照されたい).受精機能を欠損した精細胞のつくられるgcs1/hap2変異体4) の花粉管を受容することで重複受精に失敗した胚珠は,高頻度で2本目の花粉管を誘引することが明らかにされた2,3).このことから,多花粉管拒否の開始における重複受精の役割が示されたが,雌しべの組織の奥深くでほぼ同時に起こる卵細胞の受精および中央細胞の受精が,それぞれどれくらい多花粉管拒否に貢献しているかについては明らかにされていなかった.
精細胞が1つしかつくられないcdka;1変異体の花粉を用いて受粉実験を行った5).これにより,卵細胞のみが受精することで次世代の植物体となる胚だけが発達した種子や,反対に,中央細胞のみが受精することで胚乳だけが発達した種子がランダムにつくられる.これら単独受精を起こした種子は花粉管の誘引が完全には停止せず,いずれの場合も,2本目の花粉管を受け入れたものが約30%みられた.一方で,卵細胞および中央細胞ともに受精できなかった種子における2本目の花粉管の受容率80%と比較すると3),これら単独受精を示した種子は,ある程度の多花粉管拒否を起こしていることがわかった.したがって,多花粉管拒否には卵細胞の受精および中央細胞の受精により活性化される独立した経路の存在が示唆された.重複受精ではこの2つの経路は相加的に機能し,より強い多花粉管拒否が誘起されると考えられた(図1).
多花粉管拒否のしくみが花粉管への誘引能力の低下によるものかどうかを調べるため,授粉ののちの雌しべから摘出した胚珠が花粉管を誘引するかどうか培地において評価した.すると,gcs1/hap2により重複受精に失敗した約90%の胚珠,cdka;1変異体により卵細胞のみが受精した約50%の胚珠,そして,野生型の花粉管を受容して重複受精に成功した約20%の胚珠が,花粉管の誘引応答を示した.これらの傾向は,最終的な種子の表現型を調べたさきの実験結果とほぼ一致した.重要なのは,多花粉管拒否に重要なステップとみられていた“残されたほうの助細胞の核崩壊”よりまえで得られたことである.これにより,受精ののち花粉管への誘引能力を急速に低下させる多花粉管拒否のようすが明らかになった.
多花粉管拒否について分子的な理解の手がかりを得ることを目的に,中央細胞の機能に欠損を示す既知のシロイヌナズナ変異体について解析を行った.その結果,MEDEA,FIS2,FIEといったタンパク質を欠損する変異体は,重複受精するにもかかわらず高頻度で2本目の花粉管を受容することが示された.これらのタンパク質はヒストンH3の27番目のリジン残基のメチル化修飾を介して遺伝子発現を負に制御するポリコーム複合体のひとつ,FIS-PRC2(fertilization-independent seed polycomb repressive complex 2)を構成する6).MEAやFIS2は中央細胞や胚乳に特異的に発現することなどから,FIS-PRC2が中央細胞の受精のときに起こる多花粉管拒否の経路において機能していると結論づけた.
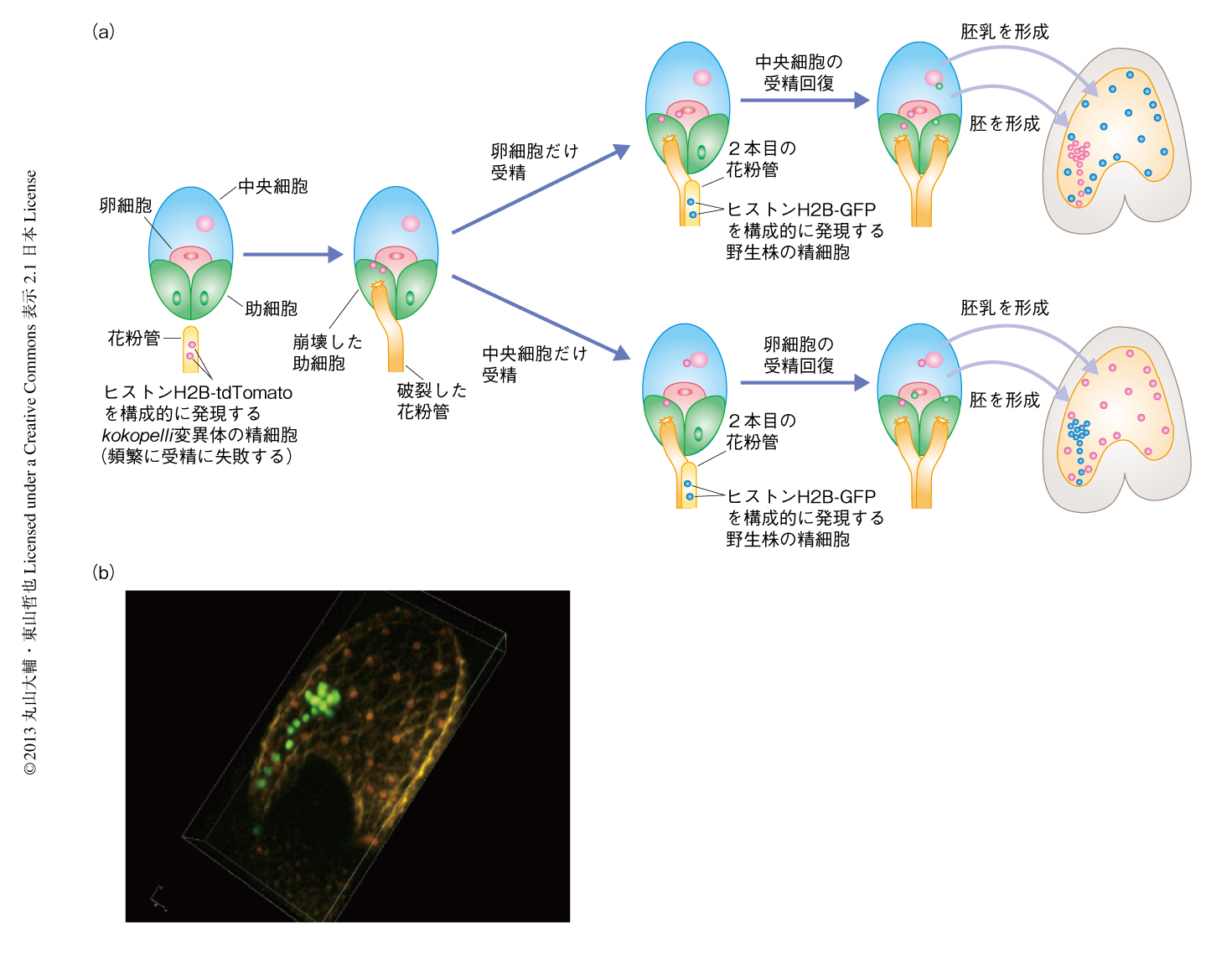
被子植物の多花粉管拒否は,どうして2種類の受精により制御される複雑なしくみになっているのだろうか? 卵細胞または中央細胞のみが受精に失敗した場合,それぞれの細胞が2本目の花粉管誘引を許可できたほうが,受精の失敗を柔軟に回復することができるのだろうと予想した.そこで,このような現象が起こりうるのかどうか検証するため,卵細胞や中央細胞において合計10%程度の単独受精をひき起こす精細胞異常変異体であるkokopelli変異体を実験に用いた7).まず,構成的な遺伝子プロモーターによりヒストンH2B-tdTomato融合タンパク質を発現するkokopelli変異体の花粉を,野生株の雌しべに授粉して一部に単独受精を誘起したのち,ヒストンH2B-GFP融合タンパク質を構成的に発現する形質転換体の花粉を追加するよう授粉した.すると予想どおり,胚の核がtdTomatoで胚乳の核がGFPで標識された種子や,逆に,胚の核がGFPで胚乳の核がtdTomatoで標識された種子が得られた(図2).このように,通常の受精様式では同じ遺伝子構成となるべき胚と胚乳が異なる遺伝子型をもつ特殊な受精現象はヘテロ受精とよばれ,これまでトウモロコシで研究されてきた8).今回,シロイヌナズナにおいてはじめてヘテロ受精を誘起することに成功した.これにより,ヘテロ受精が生じる背景には,卵細胞と中央細胞のそれぞれの受精による協調的な花粉管誘引の停止のしくみの存在することが示された.もしかすると,この柔軟なしくみが生殖における適応度の上昇に貢献したのかもしれない.
この研究により,これまで不可分とされてきた卵細胞の受精および中央細胞の受精が,それぞれ多花粉管拒否にはたす役割が明らかになった.ドイツのグループは,筆者らの報告と連報というかたちで,受精ののちに残されたほうの助細胞核の崩壊にエチレンシグナル伝達系が必要とされることを示している9).今後は,卵細胞あるいは中央細胞の受精がFIS-PRC2やエチレンシグナル伝達系とどのような関係にあるのかを特定することがひとつの課題になるだろう.そこで,助細胞の示す花粉管誘引の動的な変化をとらえる方法が重要になってくる.筆者らのグループは,シロイヌナズナにおいて助細胞から分泌される花粉管誘引ペプチドAtLURE1を同定した10).これにより,多花粉管拒否をAtLURE1の合成や分泌などの動的な制御の変化として理解することができるだろう.
一方で,シロイヌナズナにおいて誘導することのできるようになったヘテロ受精は,父親側から遺伝子型の異なる胚と胚乳をつくらせるツールとして利用できる.たとえば,胚乳から胚へと移動する小分子RNAなど情報分子の解析に役立つだろう.
略歴:2010年 名古屋大学大学院理学研究科 修了,同年 同 研究員を経て,2013年より名古屋大学トランスフォーマティブ生命分子研究所 研究員.
東山 哲也(Tetsuya Higashiyama)
名古屋大学トランスフォーマティブ生命分子研究所 教授.科学技術振興機構ERATO東山ライブホロニクスプロジェクト 研究総括 兼任.
研究室URL:http://www.higashiyama-lab.com,http://www.liveholonics.com/
© 2013 丸山大輔・東山哲也 Licensed under CC 表示 2.1 日本
(名古屋大学トランスフォーマティブ生命分子研究所)
email:丸山大輔
DOI: 10.7875/first.author.2013.070
Independent control by each female gamete prevents the attraction of multiple pollen tubes.
Daisuke Maruyama, Yuki Hamamura, Hidenori Takeuchi, Daichi Susaki, Moe Nishimaki, Daisuke Kurihara, Ryushiro D. Kasahara, Tetsuya Higashiyama
Developmental Cell, 25, 317-323 (2013)
要 約
被子植物の生殖の過程では,通常,最初に到達した花粉管により重複受精が完了する.受精した胚珠は後続の花粉管が来ないよう制御しているが,そのしくみについては明らかにされていなかった.筆者らは,シロイヌナズナを用いて,卵細胞または中央細胞のどちらかにおける受精の失敗が,2本目以降の花粉管の誘引を停止させる活性の低下をまねくことを示した.同様の表現型は,中央細胞において遺伝子発現を負に制御するFIS-PRC2の機能欠損によっても観察された.FIS-PRC2は胚乳において特異的に機能することから,卵細胞と中央細胞はそれぞれ独自のシグナル伝達経路を発達させてきたものと考えられた.興味深いことに,卵細胞または中央細胞のどちらか一方が受精した胚珠では,その不完全な重複受精を原因として誘引された2本目の花粉管により受精が回復し,その結果,遺伝子型の異なる胚と胚乳をもつ種子が得られた.したがって,卵細胞と中央細胞のそれぞれが受精したかどうかを独自に判断し,2本目の花粉管の誘引をつうじた受精回復システムを構築することにより,生殖における適応度をあげている可能性が示唆された.
はじめに
鞭毛や繊毛をもたない被子植物の精細胞は自律的な移動ができず,代わりに,精細胞は花粉管という管状の細胞により輸送される.雌しべの柱頭において受粉が起こると,花粉から伸長をはじめた花粉管は互いに競争しながら雌しべの内部へと進んでいく.雌しべの内部には受精ののちに種子となる胚珠がならんでいる.花粉管は受精を完了した胚珠をさけて未受精の胚珠にむかうため,結果として,ほぼすべての胚珠が花粉管と一対一の関係で受精する1)(図1).この2本目の花粉管をしりぞけるしくみは多花粉管拒否とよばれ2),古くからその存在は知られていたものの,どのような分子機構によるものなのかは不明であった.
そのようななか,2012年,筆者らのグループおよび米国のグループは,シロイヌナズナを用い,重複受精が多花粉管拒否の開始の鍵をにぎることを示した2,3).胚珠には花粉管を誘引する物質を分泌する助細胞が2つ存在する.到達した花粉管は,片方の助細胞の崩壊とともに,2個の精細胞を放出して卵細胞と中央細胞という雌側の2つの配偶子の受精を完了する.この被子植物に独特の受精現象が重複受精である(図1,植物の受精のしくみについては,東山 哲也, 領域融合レビュー, 1, e007, 2012 も参照されたい).受精機能を欠損した精細胞のつくられるgcs1/hap2変異体4) の花粉管を受容することで重複受精に失敗した胚珠は,高頻度で2本目の花粉管を誘引することが明らかにされた2,3).このことから,多花粉管拒否の開始における重複受精の役割が示されたが,雌しべの組織の奥深くでほぼ同時に起こる卵細胞の受精および中央細胞の受精が,それぞれどれくらい多花粉管拒否に貢献しているかについては明らかにされていなかった.
1.多花粉管拒否は卵細胞と中央細胞の二重支配をうける
精細胞が1つしかつくられないcdka;1変異体の花粉を用いて受粉実験を行った5).これにより,卵細胞のみが受精することで次世代の植物体となる胚だけが発達した種子や,反対に,中央細胞のみが受精することで胚乳だけが発達した種子がランダムにつくられる.これら単独受精を起こした種子は花粉管の誘引が完全には停止せず,いずれの場合も,2本目の花粉管を受け入れたものが約30%みられた.一方で,卵細胞および中央細胞ともに受精できなかった種子における2本目の花粉管の受容率80%と比較すると3),これら単独受精を示した種子は,ある程度の多花粉管拒否を起こしていることがわかった.したがって,多花粉管拒否には卵細胞の受精および中央細胞の受精により活性化される独立した経路の存在が示唆された.重複受精ではこの2つの経路は相加的に機能し,より強い多花粉管拒否が誘起されると考えられた(図1).
2.受精により花粉管の誘引能力はすばやく低下する
多花粉管拒否のしくみが花粉管への誘引能力の低下によるものかどうかを調べるため,授粉ののちの雌しべから摘出した胚珠が花粉管を誘引するかどうか培地において評価した.すると,gcs1/hap2により重複受精に失敗した約90%の胚珠,cdka;1変異体により卵細胞のみが受精した約50%の胚珠,そして,野生型の花粉管を受容して重複受精に成功した約20%の胚珠が,花粉管の誘引応答を示した.これらの傾向は,最終的な種子の表現型を調べたさきの実験結果とほぼ一致した.重要なのは,多花粉管拒否に重要なステップとみられていた“残されたほうの助細胞の核崩壊”よりまえで得られたことである.これにより,受精ののち花粉管への誘引能力を急速に低下させる多花粉管拒否のようすが明らかになった.
3.FIS-PRC2は多花粉管拒否の中央細胞経路において機能する
多花粉管拒否について分子的な理解の手がかりを得ることを目的に,中央細胞の機能に欠損を示す既知のシロイヌナズナ変異体について解析を行った.その結果,MEDEA,FIS2,FIEといったタンパク質を欠損する変異体は,重複受精するにもかかわらず高頻度で2本目の花粉管を受容することが示された.これらのタンパク質はヒストンH3の27番目のリジン残基のメチル化修飾を介して遺伝子発現を負に制御するポリコーム複合体のひとつ,FIS-PRC2(fertilization-independent seed polycomb repressive complex 2)を構成する6).MEAやFIS2は中央細胞や胚乳に特異的に発現することなどから,FIS-PRC2が中央細胞の受精のときに起こる多花粉管拒否の経路において機能していると結論づけた.
4.単独受精ののちの能動的な受精回復によりヘテロ受精が起こる
被子植物の多花粉管拒否は,どうして2種類の受精により制御される複雑なしくみになっているのだろうか? 卵細胞または中央細胞のみが受精に失敗した場合,それぞれの細胞が2本目の花粉管誘引を許可できたほうが,受精の失敗を柔軟に回復することができるのだろうと予想した.そこで,このような現象が起こりうるのかどうか検証するため,卵細胞や中央細胞において合計10%程度の単独受精をひき起こす精細胞異常変異体であるkokopelli変異体を実験に用いた7).まず,構成的な遺伝子プロモーターによりヒストンH2B-tdTomato融合タンパク質を発現するkokopelli変異体の花粉を,野生株の雌しべに授粉して一部に単独受精を誘起したのち,ヒストンH2B-GFP融合タンパク質を構成的に発現する形質転換体の花粉を追加するよう授粉した.すると予想どおり,胚の核がtdTomatoで胚乳の核がGFPで標識された種子や,逆に,胚の核がGFPで胚乳の核がtdTomatoで標識された種子が得られた(図2).このように,通常の受精様式では同じ遺伝子構成となるべき胚と胚乳が異なる遺伝子型をもつ特殊な受精現象はヘテロ受精とよばれ,これまでトウモロコシで研究されてきた8).今回,シロイヌナズナにおいてはじめてヘテロ受精を誘起することに成功した.これにより,ヘテロ受精が生じる背景には,卵細胞と中央細胞のそれぞれの受精による協調的な花粉管誘引の停止のしくみの存在することが示された.もしかすると,この柔軟なしくみが生殖における適応度の上昇に貢献したのかもしれない.
おわりに
この研究により,これまで不可分とされてきた卵細胞の受精および中央細胞の受精が,それぞれ多花粉管拒否にはたす役割が明らかになった.ドイツのグループは,筆者らの報告と連報というかたちで,受精ののちに残されたほうの助細胞核の崩壊にエチレンシグナル伝達系が必要とされることを示している9).今後は,卵細胞あるいは中央細胞の受精がFIS-PRC2やエチレンシグナル伝達系とどのような関係にあるのかを特定することがひとつの課題になるだろう.そこで,助細胞の示す花粉管誘引の動的な変化をとらえる方法が重要になってくる.筆者らのグループは,シロイヌナズナにおいて助細胞から分泌される花粉管誘引ペプチドAtLURE1を同定した10).これにより,多花粉管拒否をAtLURE1の合成や分泌などの動的な制御の変化として理解することができるだろう.
一方で,シロイヌナズナにおいて誘導することのできるようになったヘテロ受精は,父親側から遺伝子型の異なる胚と胚乳をつくらせるツールとして利用できる.たとえば,胚乳から胚へと移動する小分子RNAなど情報分子の解析に役立つだろう.
文 献
- Yadegari, R. & Drews, G. N.: Female gametophyte development. Plant Cell, 16, S133-S141 (2004)[PubMed]
- Beale, K. M., Leydon, A. R. & Johnson, M. A.: Gamete fusion is required to block multiple pollen tubes from entering an Arabidopsis ovule. Curr. Biol., 22, 1090-1094 (2012)[PubMed]
- Kasahara, R. D., Maruyama, D., Hamamura, Y. et al.: Fertilization recovery after defective sperm cell release in Arabidopsis. Curr. Biol., 22, 1084-1089 (2012)[PubMed]
- Mori, T., Kuroiwa, H., Higashiyama, T. et al.: GENERATIVE CELL SPECIFIC 1 is essential for angiosperm fertilization. Nat. Cell Biol., 8, 64-71 (2006)[PubMed]
- Aw, S. J., Hamamura, Y., Chen, Z. et al.: Sperm entry is sufficient to trigger division of the central cell but the paternal genome is required for endosperm development in Arabidopsis. Development, 137, 2683-2690 (2010)[PubMed]
- Kohler, C., Wolff, P. & Spillane, C.: Epigenetic mechanisms underlying genomic imprinting in plants. Annu. Rev. Plant Biol., 63, 331-352 (2012)[PubMed]
- Ron, M., Saez, M. A., Williams, L. E. et al.: Proper regulation of a sperm-specific cis-nat-siRNA is essential for double fertilization in Arabidopsis. Genes Dev., 24, 1010-1021 (2010)[PubMed]
- Sprague, G. F.: HETERO-FERTILIZATION IN MAIZE. Science, 69, 526-527 (1929)[PubMed]
- Volz, R., Heydlauff, J., Ripper, D. et al.: Ethylene signaling is required for synergid degeneration and the establishment of a pollen tube block. Dev. Cell, 25, 310-316 (2013)[PubMed]
- Takeuchi, H. & Higashiyama, T.: A species-specific cluster of defensin-like genes encodes diffusible pollen tube attractants in Arabidopsis. PLoS Biol., 10, e1001449 (2012)[PubMed]
著者プロフィール
略歴:2010年 名古屋大学大学院理学研究科 修了,同年 同 研究員を経て,2013年より名古屋大学トランスフォーマティブ生命分子研究所 研究員.
東山 哲也(Tetsuya Higashiyama)
名古屋大学トランスフォーマティブ生命分子研究所 教授.科学技術振興機構ERATO東山ライブホロニクスプロジェクト 研究総括 兼任.
研究室URL:http://www.higashiyama-lab.com,http://www.liveholonics.com/
© 2013 丸山大輔・東山哲也 Licensed under CC 表示 2.1 日本
