前頭側頭葉変性症および筋萎縮性側索硬化症の原因である非翻訳領域のGGGGCCリピート配列はジペプチドリピートタンパク質に翻訳され脳に蓄積する
森 康治1・Dieter Edbauer 2
(1ドイツLudwig-Maximilians大学Munich,Adolf Butenandt Institute,2ドイツGerman Center for Neurodegenerative Diseases)
email:森 康治
DOI: 10.7875/first.author.2013.017
The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS.
Kohji Mori, Shih-Ming Weng, Thomas Arzberger, Stephanie May, Kristin Rentzsch, Elisabeth Kremmer, Bettina Schmid, Hans A. Kretzschmar, Marc Cruts, Christine Van Broeckhoven, Christian Haass, Dieter Edbauer
Science, 339, 1335-1338 (2013)
C9orf72遺伝子のもつGGGGCCリピート配列の延長は家族性の前頭側頭葉変性症および筋萎縮性側索硬化症の原因としてもっとも多いものであるが,どのようにして神経の変性がひき起こされるのかはわかっていない.多くの神経変性疾患にはその疾患を特徴づける封入体が存在し,C9orf72遺伝子に変異をもつ患者にもニューロンに特徴的な封入体がみられるが,その封入体を構成するタンパク質は不明であった.今回,筆者らは,この封入体にポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)の3種類のジペプチドリピートタンパク質が存在することを見い出した.さらに,これらのジペプチドリピートタンパク質はC9orf72遺伝子の非翻訳領域に存在するGGGGCCリピート配列が3つのリーディングフレームにおいて開始コドンなしに開始する異常な翻訳をうけることにより生じていた.これらの知見は,家族性の前頭側頭葉変性症および筋萎縮性側索硬化症に関連する遺伝子の変異と,その変異をもつ患者に顕著にみられる神経病理学的な変化とを直接に関連づけるものであった.
老人斑の主たる構成タンパク質がアミロイドβであることが明らかにされてアルツハイマー病の病態の理解が著しく進んだように,個々の神経変性疾患の病態の解明および治療法の開発において脳に蓄積するタンパク質の本態を明らかにすることはきわめて重要なステップである1).若年性認知症において2番目に多いとされる前頭側頭葉変性症(frontotemporal lobar degeneration:FTLD)は認知症,性格変化,言語障害,行動障害などをひき起こす神経変性疾患であり,脳におもにタウの蓄積がみられるものと,おもにユビキチン陽性の封入体がみられるものとに大別される.2006年に,ユビキチン陽性の封入体の本態がTDP-43であることが同定され2,3),以後,この分野の研究は飛躍的に進展した4).古典的に運動ニューロンの変性および筋力の低下を示す神経変性疾患とされてきた筋萎縮性側索硬化症(amyotrophic lateral sclerosis:ALS)においてもTDP-43が蓄積していることが示され,この2つの疾患の病態および臨床症状におけるオーバーラップが指摘されるようになった.2つの疾患とも大多数の患者にはリン酸化TDP-43強陽性の細胞内封入体がみられる.2011年に,新たにC9orf72遺伝子に存在するGGGGCCという6塩基のリピート配列が家族性の前頭側頭葉変性症および筋萎縮性側索硬化症においてもっとも多い遺伝子変異として見い出された5-7).この異常に延長したリピート配列はC9orf72遺伝子の翻訳領域の5’側にあり,転写産物のアイソフォームにより第1イントロンともなればプロモーター領域ともなる.リピート配列の全長の塩基配列の決定は困難であるが,患者におけるGGGGCCリピートの数は数百リピート以上であり,健常人におけるリピート数は20~25リピートよりも少ないと推定されている.C9orf72遺伝子に変異をもつ患者ではリン酸化TDP-43陽性を示す封入体にくわえて,小脳や海馬,前頭側頭新皮質のニューロンにおいてリン酸化TDP-43陰性かつユビキチン関連タンパク質p62/SQSTM1陽性の封入体がみられる8).これらTDP-43陰性の封入体はC9orf72遺伝子に変異をもつ患者にきわめて特徴的であるが,その構成タンパク質は明らかではない.
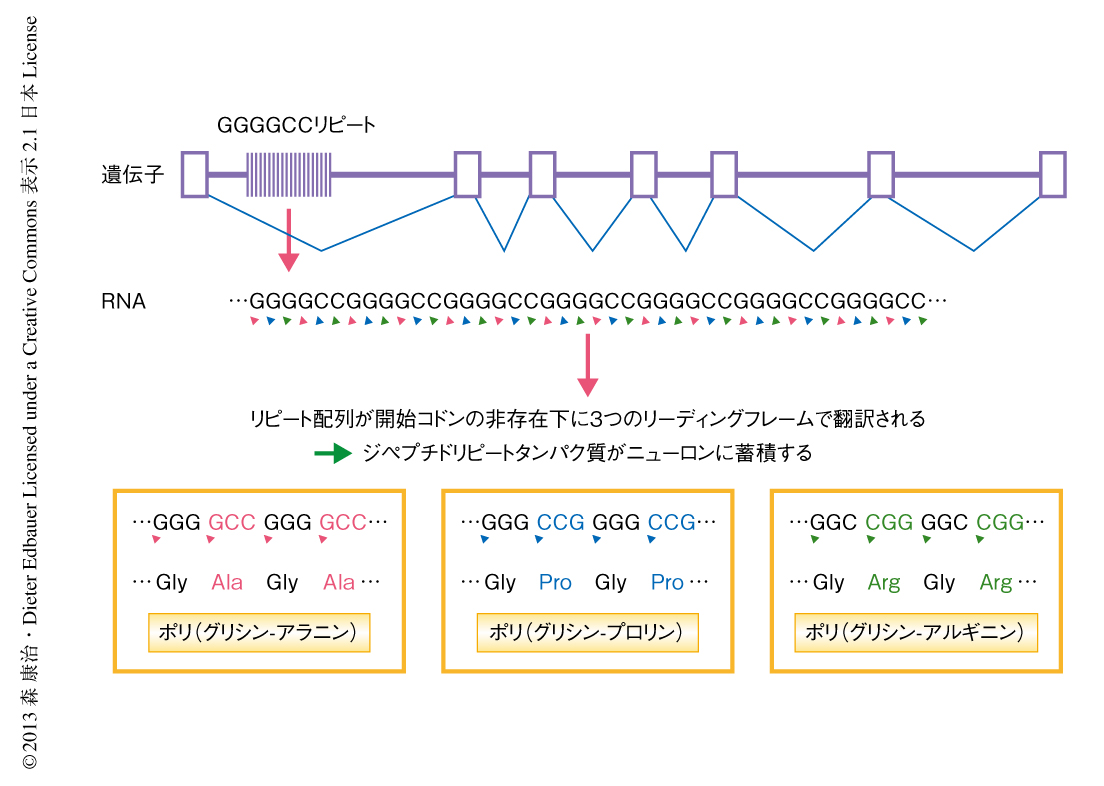
GGGGCCリピートの延長がひき起こす病態の分子機構として,2つの仮説が提唱されている.ひとつは,リピート配列がC9orf72変異対立遺伝子の転写もしくはスプライシングの異常をひき起こし,C9orf72タンパク質のハプロ不全が生じるとするものである6).C9orf72タンパク質の機能の詳細はいまだ不明であるが,小胞輸送に関連すると推測されている.もうひとつの仮説は,異常に延長したリピートをもつRNAにRNA結合タンパク質が選択的かつ大量に吸着されることにより,それらのRNA結合タンパク質の機能不全をきたし神経変性にいたるとするもので,同じく,非翻訳領域におけるリピートの延長によりひき起こされる神経変性疾患である筋緊張性ジストロフィーと同様の疾患機序を想定するものである5).筆者らは,第3の仮説として,イントロンに存在するGGGGCCリピート配列がジペプチドリピートタンパク質へと翻訳されるのではないかと考えた.きわめてまれではあるが,開始コドンであるATGの非存在下にて開始する特殊な形式での翻訳が起こりうるとされており,non-ATG-initiated translationとよばれている9).
GGGGCCリピート配列が3つのリーディングフレームにおいて翻訳されるとすると,3種類のジペプチドリピートタンパク質,すなわち,ポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)が生じることになる.このうち,ポリ(グリシン-アラニン)およびポリ(グリシン-プロリン)は極度に疎水性であるため細胞において凝集体を生じる可能性がある.ミエリン塩基性タンパク質と融合させた15リピートの(グリシン-アラニン)および15リピートの(グリシン-プロリン)を抗原として,抗ポリ(グリシン-アラニン)ポリクローナル抗体および抗ポリ(グリシン-プロリン)ポリクローナル抗体を作製し,さらに,6リピートの(グリシン-アルギニン)をもつEBNA2Aに対し作製された抗ポリ(グリシン-アルギニン)モノクローナル抗体を得た.アフィニティ精製ののち,これらの抗体はイムノブロット法において,ポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)をそれぞれ特異的に認識した.
GGGGCCリピート配列が開始コドンの非存在下でジペプチドリピートタンパク質に翻訳されるのかどうか確かめるため,C9orf72遺伝子に変異をもつ患者に由来する細胞からリピート数の異なるGGGGCCリピート配列をクローニングした.GGGGCCリピート配列の上流は開始コドンであるATGを欠いており,それぞれのリーディングフレームに4つもしくは5つの終止コドンを含んでいた.HEK293細胞にこれらのコンストラクトを発現させイムノブロット法により解析した.すると,抗ポリ(グリシン-アラニン)抗体は38リピート以上においてリピート数に応じたバンドを認識した.タンパク質のシグナル強度はリピート数が多くなるほどに強くなったことから,この翻訳機構はリピート配列が延長するほどより効率的になるものと考えられた.ポリ(グリシン-アルギニン)は検出できず,ポリ(グリシン-プロリン)はもっとも多いリピート数(およそ145リピート)のコンストラクトを発現させた細胞のみで検出された.
凝集したジペプチドリピートタンパク質が脳に存在するかどうかを解析するため,C9orf72遺伝子に変異をもつ患者3例,および,C9orf72遺伝子に変異をもたない6例(健常対照2例,前頭側頭葉変性症2例,筋萎縮性側索硬化症2例)の小脳のリン酸緩衝液不溶画分に2%SDSをくわえフィルタートラップアッセイを行った.C9orf72遺伝子に変異をもつ患者においてのみ,強いポリ(グリシン-アラニン)のシグナル,および,それより弱いながらもポリ(グリシン-プロリン)およびポリ(グリシン-アルギニン)のシグナルが検出された.さらに,小脳の2%SDS不溶画分に8%SDSをくわえ一部を可溶化してウェスタンブロット解析を行ったところ,凝集したジペプチドリピートタンパク質がC9orf72遺伝子に変異をもつ患者において特異的に,ゲルの最上部に相当する部位に検出された.
転写やスプライシングの異常によりC9orf72変異対立遺伝子に由来するmRNAの発現は低下していることが報告されている.筆者らの手においても,同様に,C9orf72遺伝子に変異をもつ患者の脳においてC9orf72 mRNAのレベルは変異の非保持者の約6割に低下していた.しかしながら,GGGGCCリピート配列の近傍にかぎると,センス鎖およびアンチセンス鎖に由来するRNAはともに,C9orf72遺伝子に変異をもつ患者では変異の非保持者に対し約8倍の増加が認められた.この結果から,C9orf72 mRNAの全体としては減少しているものの,リピート配列を含むmRNA前駆体,もしくは,そこから切り出されたリピート配列の近傍の領域のみが選択的に安定化されているものと考えられた.
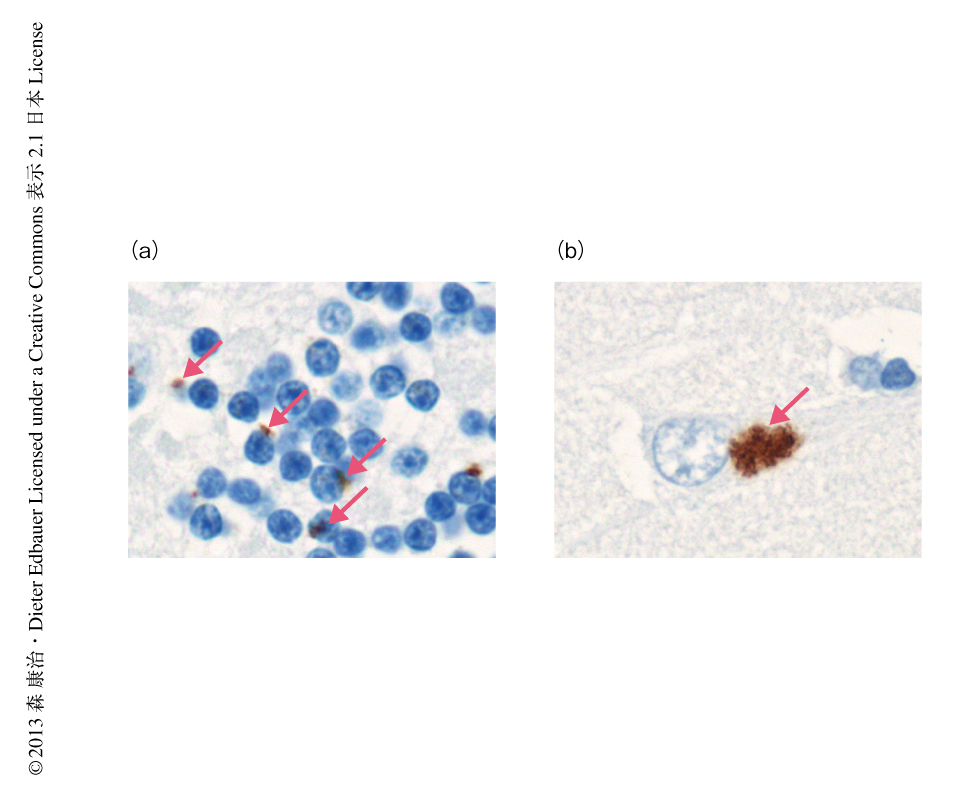
脳におけるジペプチドリピートタンパク質の分布を調べるため,C9orf72遺伝子に変異をもつ患者の剖検脳において,ユビキチン関連タンパク質であるp62陽性かつリン酸化TDP-43陰性の封入体のみられる海馬と小脳を中心に免疫組織化学染色を行った.抗ポリ(グリシン-アラニン)抗体はC9orf72遺伝子に変異をもつ患者の脳の顆粒細胞層のニューロンにおいて点状(dot-like)の封入体を検出した(図1a).その形態や数はp62陽性の封入体に酷似していた.さらに,同様の封入体は,少ないながらも抗ポリ(グリシン-プロリン抗体)および抗ポリ(グリシン-アルギニン)抗体によっても検出された.一方,海馬のニューロンにおいては,p62陽性を示す星状(star-like)の封入体と非常によく似た封入体の大多数が,3つの抗ジペプチドリピート抗体により陽性を示した(図1b).同様の封入体は海馬歯状回の顆粒細胞層や小脳の分子層,さらに,新皮質においても認められ,検索したC9orf72遺伝子に変異をもつ患者10例すべてにおいて,3種類すべての抗ジペプチドリピート抗体に対し,陽性を示す封入体がそれぞれ確認された.
これらジペプチドリピートタンパク質からなる封入体がp62陽性かつリン酸化TDP-43陰性の封入体と同一なのか検討するため,C9orf72遺伝子に変異をもつ患者の海馬および小脳の切片において免疫二重染色を行った.抗ポリ(グリシン-アラニン)抗体,抗ポリ(グリシン-プロリン)抗体,抗ポリ(グリシン-アルギニン)抗体に陽性の封入体は,p62陽性の封入体と共局在した.一方で,これらの抗ジペプチドリピート抗体に陽性の封入体とリン酸化TDP-43とは共局在しなかった.ごくまれではあったが,ポリ(グリシン-アラニン)陽性の小さな球状の封入体がリン酸化TDP-43陽性の封入体にかこまれるよう存在しTDP-43陽性の封入体の核を形成している像が得られたことから,ジペプチドリピートタンパク質の凝集はリン酸化TDP-43の凝集に先行している可能性が示唆された.定量的な解析においても,p62陽性の封入体とポリ(グリシン-アラニン)陽性の封入体とは共局在すること,p62陽性の封入体の一部はポリ(グリシン-プロリン)陽性の封入体およびポリ(グリシン-アルギニン)陽性の封入体と共局在することを確認した.フィルタートラップアッセイの結果と同様に,こうしたジペプチドリピートタンパク質からなる封入体は,CAGリピート配列の延長をもつハンチントン病を含む,C9orf72遺伝子に変異をもたない12例の対照(前頭側頭葉変性症2例,筋萎縮性側索硬化症2例,ハンチントン病2例,健常対照6例)のいずれにおいても認められなかった.
C9orf72遺伝子に変異をもつ家族性の前頭側頭葉変性症の患者には例外的に,脳にごくわずかしかリン酸化TDP-43陽性の封入体をもたず,一方で,著しいユビキチン陽性の封入体を示しFTLD-UPS(UPS:ubiquitin proteasome system)とよばれるケースがある.このような患者の側頭葉皮質において,多数のポリ(グリシン-アラニン)陽性の封入体,および,いくらかのポリ(グリシン-プロリン)陽性あるいはポリ(グリシン-アルギニン)陽性の封入体が認められた.このようにC9orf72遺伝子の変異に関連するFTLD-UPSのケースにおいては,TDP-43ではなくジペプチドリピートタンパク質が神経変性の過程においてきわめて重要な役割をはたしている可能性が示唆された.
筆者らは,C9orf72遺伝子に変異をもつ家族性の前頭側頭葉変性症および筋萎縮性側索硬化症の患者において,イントロン領域に存在する異常に延長したGGGGCCリピート配列が開始コドンであるATGの非存在下にて開始する特殊な形式での翻訳をうけ,その結果,産生された不溶性のジペプチドリピートタンパク質が脳に蓄積することを示した(図2).この結果は,C9orf72遺伝子におけるリピート配列の延長がC9orf72タンパク質のハプロ不全をひき起こす,もしくは,異常に延長したリピート配列をもつmRNAにRNA結合タンパク質が大量に結合し機能不全をひき起こすとするほかの2つの仮説を否定するものではなく,いずれの要因も重なり合ってC9orf72遺伝子変異による病態を構成している可能性がある.現に筆者らは,GGGGCCリピート配列に特異的なRNA結合タンパク質としてhnRNP A3を同定し,C9orf72遺伝子に変異をもつ患者のニューロンにおいてhnRNP A3が通常は存在するはずの核から細胞質へと移行しp62陽性の封入体の一部と共局在していることを示し,別に報告した10).C9orf72遺伝子に変異をもつ患者では核においてhnRNP A3の機能不全の生じている可能性がある.また,hnRNP A3はスプライシングの抑制およびRNAの核外輸送に関与するとされており,GGGGCCリピート配列をもつRNAの安定化および細胞質(すなわち,翻訳の場)への輸送にも関連している可能性がある.
一方で,多くの証拠がジペプチドリピートタンパク質の毒性を示唆している.第1に,ジペプチドリピートタンパク質の蓄積が海馬や前頭側頭皮質など臨床的に関連した脳領域において多くみられ,TDP-43の蓄積に先行している可能性のあること.第2に,C9orf72遺伝子に変異をもつ患者ではジペプチドリピートタンパク質の蓄積が特徴的にみられる小脳において萎縮が認められるが,これはほかの家族性の前頭側頭葉変性症および筋萎縮性側索硬化症では認められないこと.第3に,C9orf72遺伝子に変異をもつ患者の少なくとも一例において,ジペプチドリピートタンパク質の顕著な蓄積とともに行動型前頭側頭型認知症(behavioral variant frontotemporal dementia)がみられたが,TDP-43の蓄積は認められなかったこと.最後に,ジペプチドリピートタンパク質の蓄積は家族性の前頭側頭葉変性症および筋萎縮性側索硬化症のもっとも多い原因であるGGGGCCリピート配列の延長による直接の結果であること.これらのことから,筆者らは,神経病理分類を改定する際にFTLD/ALS-DPR(DPR:ジペプチドリピートタンパク質)という呼称を設けることを提唱した.
なお,ここで解説した筆者らの論文がScience誌に発表されてからおよそ1週間後,別のグループからも,ポリ(グリシン-プロリン)がC9orf72遺伝子に変異をもつ患者の脳において蓄積しているとする論文が発表された11).
略歴:2010年 大阪大学大学院医学系研究科 修了,2011年よりドイツLudwig-Maximilians大学Munichフンボルトリサーチフェロー.精神保健指定医.
研究テーマ:精神神経疾患の病理病態にもとづく診断法および治療法の開発.
Dieter Edbauer
ドイツGerman Center for Neurodegenerative Diseases教授.
© 2013 森 康治・Dieter Edbauer Licensed under CC 表示 2.1 日本
(1ドイツLudwig-Maximilians大学Munich,Adolf Butenandt Institute,2ドイツGerman Center for Neurodegenerative Diseases)
email:森 康治
DOI: 10.7875/first.author.2013.017
The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS.
Kohji Mori, Shih-Ming Weng, Thomas Arzberger, Stephanie May, Kristin Rentzsch, Elisabeth Kremmer, Bettina Schmid, Hans A. Kretzschmar, Marc Cruts, Christine Van Broeckhoven, Christian Haass, Dieter Edbauer
Science, 339, 1335-1338 (2013)
要 約
C9orf72遺伝子のもつGGGGCCリピート配列の延長は家族性の前頭側頭葉変性症および筋萎縮性側索硬化症の原因としてもっとも多いものであるが,どのようにして神経の変性がひき起こされるのかはわかっていない.多くの神経変性疾患にはその疾患を特徴づける封入体が存在し,C9orf72遺伝子に変異をもつ患者にもニューロンに特徴的な封入体がみられるが,その封入体を構成するタンパク質は不明であった.今回,筆者らは,この封入体にポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)の3種類のジペプチドリピートタンパク質が存在することを見い出した.さらに,これらのジペプチドリピートタンパク質はC9orf72遺伝子の非翻訳領域に存在するGGGGCCリピート配列が3つのリーディングフレームにおいて開始コドンなしに開始する異常な翻訳をうけることにより生じていた.これらの知見は,家族性の前頭側頭葉変性症および筋萎縮性側索硬化症に関連する遺伝子の変異と,その変異をもつ患者に顕著にみられる神経病理学的な変化とを直接に関連づけるものであった.
はじめに
老人斑の主たる構成タンパク質がアミロイドβであることが明らかにされてアルツハイマー病の病態の理解が著しく進んだように,個々の神経変性疾患の病態の解明および治療法の開発において脳に蓄積するタンパク質の本態を明らかにすることはきわめて重要なステップである1).若年性認知症において2番目に多いとされる前頭側頭葉変性症(frontotemporal lobar degeneration:FTLD)は認知症,性格変化,言語障害,行動障害などをひき起こす神経変性疾患であり,脳におもにタウの蓄積がみられるものと,おもにユビキチン陽性の封入体がみられるものとに大別される.2006年に,ユビキチン陽性の封入体の本態がTDP-43であることが同定され2,3),以後,この分野の研究は飛躍的に進展した4).古典的に運動ニューロンの変性および筋力の低下を示す神経変性疾患とされてきた筋萎縮性側索硬化症(amyotrophic lateral sclerosis:ALS)においてもTDP-43が蓄積していることが示され,この2つの疾患の病態および臨床症状におけるオーバーラップが指摘されるようになった.2つの疾患とも大多数の患者にはリン酸化TDP-43強陽性の細胞内封入体がみられる.2011年に,新たにC9orf72遺伝子に存在するGGGGCCという6塩基のリピート配列が家族性の前頭側頭葉変性症および筋萎縮性側索硬化症においてもっとも多い遺伝子変異として見い出された5-7).この異常に延長したリピート配列はC9orf72遺伝子の翻訳領域の5’側にあり,転写産物のアイソフォームにより第1イントロンともなればプロモーター領域ともなる.リピート配列の全長の塩基配列の決定は困難であるが,患者におけるGGGGCCリピートの数は数百リピート以上であり,健常人におけるリピート数は20~25リピートよりも少ないと推定されている.C9orf72遺伝子に変異をもつ患者ではリン酸化TDP-43陽性を示す封入体にくわえて,小脳や海馬,前頭側頭新皮質のニューロンにおいてリン酸化TDP-43陰性かつユビキチン関連タンパク質p62/SQSTM1陽性の封入体がみられる8).これらTDP-43陰性の封入体はC9orf72遺伝子に変異をもつ患者にきわめて特徴的であるが,その構成タンパク質は明らかではない.
GGGGCCリピートの延長がひき起こす病態の分子機構として,2つの仮説が提唱されている.ひとつは,リピート配列がC9orf72変異対立遺伝子の転写もしくはスプライシングの異常をひき起こし,C9orf72タンパク質のハプロ不全が生じるとするものである6).C9orf72タンパク質の機能の詳細はいまだ不明であるが,小胞輸送に関連すると推測されている.もうひとつの仮説は,異常に延長したリピートをもつRNAにRNA結合タンパク質が選択的かつ大量に吸着されることにより,それらのRNA結合タンパク質の機能不全をきたし神経変性にいたるとするもので,同じく,非翻訳領域におけるリピートの延長によりひき起こされる神経変性疾患である筋緊張性ジストロフィーと同様の疾患機序を想定するものである5).筆者らは,第3の仮説として,イントロンに存在するGGGGCCリピート配列がジペプチドリピートタンパク質へと翻訳されるのではないかと考えた.きわめてまれではあるが,開始コドンであるATGの非存在下にて開始する特殊な形式での翻訳が起こりうるとされており,non-ATG-initiated translationとよばれている9).
1.抗ジペプチドリピートタンパク質抗体の作製
GGGGCCリピート配列が3つのリーディングフレームにおいて翻訳されるとすると,3種類のジペプチドリピートタンパク質,すなわち,ポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)が生じることになる.このうち,ポリ(グリシン-アラニン)およびポリ(グリシン-プロリン)は極度に疎水性であるため細胞において凝集体を生じる可能性がある.ミエリン塩基性タンパク質と融合させた15リピートの(グリシン-アラニン)および15リピートの(グリシン-プロリン)を抗原として,抗ポリ(グリシン-アラニン)ポリクローナル抗体および抗ポリ(グリシン-プロリン)ポリクローナル抗体を作製し,さらに,6リピートの(グリシン-アルギニン)をもつEBNA2Aに対し作製された抗ポリ(グリシン-アルギニン)モノクローナル抗体を得た.アフィニティ精製ののち,これらの抗体はイムノブロット法において,ポリ(グリシン-アラニン),ポリ(グリシン-プロリン),ポリ(グリシン-アルギニン)をそれぞれ特異的に認識した.
2.非翻訳領域のGGGGCCリピート配列はジペプチドリピートタンパク質に翻訳される
GGGGCCリピート配列が開始コドンの非存在下でジペプチドリピートタンパク質に翻訳されるのかどうか確かめるため,C9orf72遺伝子に変異をもつ患者に由来する細胞からリピート数の異なるGGGGCCリピート配列をクローニングした.GGGGCCリピート配列の上流は開始コドンであるATGを欠いており,それぞれのリーディングフレームに4つもしくは5つの終止コドンを含んでいた.HEK293細胞にこれらのコンストラクトを発現させイムノブロット法により解析した.すると,抗ポリ(グリシン-アラニン)抗体は38リピート以上においてリピート数に応じたバンドを認識した.タンパク質のシグナル強度はリピート数が多くなるほどに強くなったことから,この翻訳機構はリピート配列が延長するほどより効率的になるものと考えられた.ポリ(グリシン-アルギニン)は検出できず,ポリ(グリシン-プロリン)はもっとも多いリピート数(およそ145リピート)のコンストラクトを発現させた細胞のみで検出された.
3.ジペプチドリピートタンパク質はC9orf72遺伝子に変異をもつ患者の脳において凝集する
凝集したジペプチドリピートタンパク質が脳に存在するかどうかを解析するため,C9orf72遺伝子に変異をもつ患者3例,および,C9orf72遺伝子に変異をもたない6例(健常対照2例,前頭側頭葉変性症2例,筋萎縮性側索硬化症2例)の小脳のリン酸緩衝液不溶画分に2%SDSをくわえフィルタートラップアッセイを行った.C9orf72遺伝子に変異をもつ患者においてのみ,強いポリ(グリシン-アラニン)のシグナル,および,それより弱いながらもポリ(グリシン-プロリン)およびポリ(グリシン-アルギニン)のシグナルが検出された.さらに,小脳の2%SDS不溶画分に8%SDSをくわえ一部を可溶化してウェスタンブロット解析を行ったところ,凝集したジペプチドリピートタンパク質がC9orf72遺伝子に変異をもつ患者において特異的に,ゲルの最上部に相当する部位に検出された.
4.GGGGCCリピート配列をもつmRNAはC9orf72遺伝子に変異をもつ患者の脳において増加する
転写やスプライシングの異常によりC9orf72変異対立遺伝子に由来するmRNAの発現は低下していることが報告されている.筆者らの手においても,同様に,C9orf72遺伝子に変異をもつ患者の脳においてC9orf72 mRNAのレベルは変異の非保持者の約6割に低下していた.しかしながら,GGGGCCリピート配列の近傍にかぎると,センス鎖およびアンチセンス鎖に由来するRNAはともに,C9orf72遺伝子に変異をもつ患者では変異の非保持者に対し約8倍の増加が認められた.この結果から,C9orf72 mRNAの全体としては減少しているものの,リピート配列を含むmRNA前駆体,もしくは,そこから切り出されたリピート配列の近傍の領域のみが選択的に安定化されているものと考えられた.
5.ジペプチドリピートタンパク質はC9orf72遺伝子に変異をもつ患者のニューロンにおいて封入体を形成する
脳におけるジペプチドリピートタンパク質の分布を調べるため,C9orf72遺伝子に変異をもつ患者の剖検脳において,ユビキチン関連タンパク質であるp62陽性かつリン酸化TDP-43陰性の封入体のみられる海馬と小脳を中心に免疫組織化学染色を行った.抗ポリ(グリシン-アラニン)抗体はC9orf72遺伝子に変異をもつ患者の脳の顆粒細胞層のニューロンにおいて点状(dot-like)の封入体を検出した(図1a).その形態や数はp62陽性の封入体に酷似していた.さらに,同様の封入体は,少ないながらも抗ポリ(グリシン-プロリン抗体)および抗ポリ(グリシン-アルギニン)抗体によっても検出された.一方,海馬のニューロンにおいては,p62陽性を示す星状(star-like)の封入体と非常によく似た封入体の大多数が,3つの抗ジペプチドリピート抗体により陽性を示した(図1b).同様の封入体は海馬歯状回の顆粒細胞層や小脳の分子層,さらに,新皮質においても認められ,検索したC9orf72遺伝子に変異をもつ患者10例すべてにおいて,3種類すべての抗ジペプチドリピート抗体に対し,陽性を示す封入体がそれぞれ確認された.
これらジペプチドリピートタンパク質からなる封入体がp62陽性かつリン酸化TDP-43陰性の封入体と同一なのか検討するため,C9orf72遺伝子に変異をもつ患者の海馬および小脳の切片において免疫二重染色を行った.抗ポリ(グリシン-アラニン)抗体,抗ポリ(グリシン-プロリン)抗体,抗ポリ(グリシン-アルギニン)抗体に陽性の封入体は,p62陽性の封入体と共局在した.一方で,これらの抗ジペプチドリピート抗体に陽性の封入体とリン酸化TDP-43とは共局在しなかった.ごくまれではあったが,ポリ(グリシン-アラニン)陽性の小さな球状の封入体がリン酸化TDP-43陽性の封入体にかこまれるよう存在しTDP-43陽性の封入体の核を形成している像が得られたことから,ジペプチドリピートタンパク質の凝集はリン酸化TDP-43の凝集に先行している可能性が示唆された.定量的な解析においても,p62陽性の封入体とポリ(グリシン-アラニン)陽性の封入体とは共局在すること,p62陽性の封入体の一部はポリ(グリシン-プロリン)陽性の封入体およびポリ(グリシン-アルギニン)陽性の封入体と共局在することを確認した.フィルタートラップアッセイの結果と同様に,こうしたジペプチドリピートタンパク質からなる封入体は,CAGリピート配列の延長をもつハンチントン病を含む,C9orf72遺伝子に変異をもたない12例の対照(前頭側頭葉変性症2例,筋萎縮性側索硬化症2例,ハンチントン病2例,健常対照6例)のいずれにおいても認められなかった.
6.ポリ(グリシン-アラニン)は著しいユビキチン陽性の封入体のみを示す前頭側頭葉変性症のサブタイプにおける主たる凝集タンパク質である
C9orf72遺伝子に変異をもつ家族性の前頭側頭葉変性症の患者には例外的に,脳にごくわずかしかリン酸化TDP-43陽性の封入体をもたず,一方で,著しいユビキチン陽性の封入体を示しFTLD-UPS(UPS:ubiquitin proteasome system)とよばれるケースがある.このような患者の側頭葉皮質において,多数のポリ(グリシン-アラニン)陽性の封入体,および,いくらかのポリ(グリシン-プロリン)陽性あるいはポリ(グリシン-アルギニン)陽性の封入体が認められた.このようにC9orf72遺伝子の変異に関連するFTLD-UPSのケースにおいては,TDP-43ではなくジペプチドリピートタンパク質が神経変性の過程においてきわめて重要な役割をはたしている可能性が示唆された.
おわりに
筆者らは,C9orf72遺伝子に変異をもつ家族性の前頭側頭葉変性症および筋萎縮性側索硬化症の患者において,イントロン領域に存在する異常に延長したGGGGCCリピート配列が開始コドンであるATGの非存在下にて開始する特殊な形式での翻訳をうけ,その結果,産生された不溶性のジペプチドリピートタンパク質が脳に蓄積することを示した(図2).この結果は,C9orf72遺伝子におけるリピート配列の延長がC9orf72タンパク質のハプロ不全をひき起こす,もしくは,異常に延長したリピート配列をもつmRNAにRNA結合タンパク質が大量に結合し機能不全をひき起こすとするほかの2つの仮説を否定するものではなく,いずれの要因も重なり合ってC9orf72遺伝子変異による病態を構成している可能性がある.現に筆者らは,GGGGCCリピート配列に特異的なRNA結合タンパク質としてhnRNP A3を同定し,C9orf72遺伝子に変異をもつ患者のニューロンにおいてhnRNP A3が通常は存在するはずの核から細胞質へと移行しp62陽性の封入体の一部と共局在していることを示し,別に報告した10).C9orf72遺伝子に変異をもつ患者では核においてhnRNP A3の機能不全の生じている可能性がある.また,hnRNP A3はスプライシングの抑制およびRNAの核外輸送に関与するとされており,GGGGCCリピート配列をもつRNAの安定化および細胞質(すなわち,翻訳の場)への輸送にも関連している可能性がある.
一方で,多くの証拠がジペプチドリピートタンパク質の毒性を示唆している.第1に,ジペプチドリピートタンパク質の蓄積が海馬や前頭側頭皮質など臨床的に関連した脳領域において多くみられ,TDP-43の蓄積に先行している可能性のあること.第2に,C9orf72遺伝子に変異をもつ患者ではジペプチドリピートタンパク質の蓄積が特徴的にみられる小脳において萎縮が認められるが,これはほかの家族性の前頭側頭葉変性症および筋萎縮性側索硬化症では認められないこと.第3に,C9orf72遺伝子に変異をもつ患者の少なくとも一例において,ジペプチドリピートタンパク質の顕著な蓄積とともに行動型前頭側頭型認知症(behavioral variant frontotemporal dementia)がみられたが,TDP-43の蓄積は認められなかったこと.最後に,ジペプチドリピートタンパク質の蓄積は家族性の前頭側頭葉変性症および筋萎縮性側索硬化症のもっとも多い原因であるGGGGCCリピート配列の延長による直接の結果であること.これらのことから,筆者らは,神経病理分類を改定する際にFTLD/ALS-DPR(DPR:ジペプチドリピートタンパク質)という呼称を設けることを提唱した.
なお,ここで解説した筆者らの論文がScience誌に発表されてからおよそ1週間後,別のグループからも,ポリ(グリシン-プロリン)がC9orf72遺伝子に変異をもつ患者の脳において蓄積しているとする論文が発表された11).
文 献
- Haass, C. & Selkoe, D. J.: Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer's amyloid β-peptide. Nat. Rev. Mol. Cell Biol., 8, 101-112 (2007)[PubMed]
- Arai, T., Hasegawa, M., Akiyama, H. et al.: TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun., 351, 602-611 (2006)[PubMed]
- Neumann, M., Sampathu, D. M., Kwong, L. K. et al.: Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science, 314, 130-133 (2006)[PubMed]
- Rademakers, R., Neumann, M., Mackenzie, I. R.: Advances in understanding the molecular basis of frontotemporal dementia. Nat. Rev. Neurol., 8, 423-434 (2012)[PubMed]
- DeJesus-Hernandez, M., Mackenzie, I. R., Boeve, B. F. et al.: Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron, 72, 245-256 (2011)[PubMed]
- Gijselinck, I., Van Langenhove, T., van der Zee, J. et al.: A C9orf72 promoter repeat expansion in a Flanders-Belgian cohort with disorders of the frontotemporal lobar degeneration-amyotrophic lateral sclerosis spectrum: a gene identification study. Lancet Neurol., 11, 54-65 (2012)[PubMed]
- Renton, A. E., Majounie, E., Waite, A. et al.: A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron, 72, 257-268 (2011)[PubMed]
- Al-Sarraj, S., King, A., Troakes, C. et al.: p62 positive, TDP-43 negative, neuronal cytoplasmic and intranuclear inclusions in the cerebellum and hippocampus define the pathology of C9orf72-linked FTLD and MND/ALS. Acta Neuropathol., 122, 691-702 (2011)[PubMed]
- Zu, T., Gibbens, B., Doty, N. S. et al.: Non-ATG-initiated translation directed by microsatellite expansions. Proc. Natl. Acad. Sci. USA, 108, 260-265 (2011)[PubMed]
- Mori, K., Lammich, S., Mackenzie, I. R., et al.: hnRNP A3 binds to GGGGCC repeats and is a constituent of p62-positive/TDP43-negative inclusions in the hippocampus of patients with C9orf72 mutations. Acta Neuropathol., 125, 413-423 (2013)[PubMed]
- Ash, P. E., Bieniek, K. F., Gendron, T. F. et al.: Unconventional translation of C9ORF72 GGGGCC expansion generates insoluble polypeptides specific to c9FTD/ALS. Neuron, 77, 639-646 (2013)[PubMed]
著者プロフィール
略歴:2010年 大阪大学大学院医学系研究科 修了,2011年よりドイツLudwig-Maximilians大学Munichフンボルトリサーチフェロー.精神保健指定医.
研究テーマ:精神神経疾患の病理病態にもとづく診断法および治療法の開発.
Dieter Edbauer
ドイツGerman Center for Neurodegenerative Diseases教授.
© 2013 森 康治・Dieter Edbauer Licensed under CC 表示 2.1 日本
