ショウジョウバエの転写因子Fruitlessに拮抗的に結合する2種類のクロマチン制御タンパク質のはたらきにより単一の性的二型ニューロンがつくられる
伊藤弘樹・山元大輔
(東北大学大学院生命科学研究科 脳機能遺伝分野)
email:伊藤弘樹,山元大輔
DOI: 10.7875/first.author.2012.076
Fruitless recruits two antagonistic chromatin factors to establish single-neuron sexual dimorphism.
Hiroki Ito, Kosei Sato, Masayuki Koganezawa, Manabu Ote, Ken Matsumoto, Chihiro Hama, Daisuke Yamamoto
Cell, 149, 1327-1338 (2012)
ショウジョウバエのfruitless遺伝子にコードされるFruitlessは転写因子であり,雄の性行動に必要な神経回路の主要部分を組み立てるはたらきをする.この研究では,Fruitlessは転写共役因子Bonusを介し2種類のクロマチン制御タンパク質HDAC1およびHP1aと複合体を形成するが,BonusがHDAC1をリクルートした場合にはFruitless-Bonus-HDAC1複合体は性的二型ニューロンの雄化を誘導し,BonusがHP1aをリクルートした場合にはFruitless-Bonus-HP1a複合体はそれを阻害することを明らかにした.おもしろいことに,HDAC1あるいはHP1aの発現をノックダウンした場合,性的二型ニューロンを構成する雄型の細胞と雌型の細胞との比率は変化するが,雌雄の中間のかたちをした細胞は決して出現しなかった.したがって,性的二型ニューロンの性は単一の細胞のレベルにおいて全か無かに制御されていると考えられた.
ヒトを含め,動物の脳には雌雄の差のあることが40年ほどまえから報告されてきた.たとえば,脳の中心の下部にある視床下部の付近には前視床下部間質核とよばれる領域があり,その第3神経細胞群は男性のほうが大きく細胞の数も多いことが報告されている.さらに,同性愛の男性ではこの部分が小さく女性のものとほぼ等しいという報告もある1).しかし,解析のむずかしさもあり性差を生み出す分子機構はほとんどわかっていない.一方,ショウジョウバエを用いた場合,遺伝学的な手法により個々のニューロンの標識と操作が可能であり,さらに,その性行動は定型的であるため行動を定量解析するのに適している.この研究では,ショウジョウバエの脳の雄化に重要な役割をはたすfruitless(fru)遺伝子について,その産物である転写因子Fruと協調してはたらくタンパク質を探索し,そのタンパク質のはたらきによる標的遺伝子のオンとオフが性差の形成の根源にあることを明らかにした.
ショウジョウバエの性行動は以下の要素からなる.まず雄が雌に定位(orientation)し,前脚で雌の腹部を叩き(tapping),雌を追いかけながら片方の翅を振ってラブソングを奏でる(singing).雌が立ち止まると雄は雌の背後にまわり口吻で雌の交尾器をなめ(licking),雌の背中に乗り腹部を内側に曲げて交尾を試みる(attempted copulation).雌が性的に受け入れる“気持ち”になると雌は膣口を開き交尾(copulation)がはじまる.雌による交尾の受け入れ以外はすべて雄に特異的な行動であり,野生型の雌は決して雄のような行動はしない.通常,野生型の雄は雌に対し求愛を行い,ほかの雄に対して持続的な求愛をすることはない.
しかし,fru遺伝子の機能を低下させた場合,雄が雄に求愛したり,雄雌のどちらにも求愛しなくなったりする.fru遺伝子のmRNAは性に特異的なスプライシングをうけ,翻訳によりFruタンパク質のつくられるのは雄の神経系のみであり,雌の神経系はFruタンパク質を欠く2-4).近年,ノックイン法を用いたfru遺伝子の改変により雌雄いずれの神経系でもFruのつくられるfruM変異体が作製されているが,おもしろいことに,このfruM変異体の雌はほかの雌に対し雄の行動パターンで求愛を行う5).つまり,個体の性が雄であるか雌であるかに関係なく,fru遺伝子を発現するニューロンにFruをつくらせれば雄の求愛行動を誘起できる.この事実は,Fruが神経系の雄化タンパク質であることを示唆している.ショウジョウバエの脳には約10万個のニューロンがあるが,そのうち2000個のニューロンにfru遺伝子は発現しており,このfru遺伝子の発現ニューロンの一部は細胞の数およびかたちについて性的二型ニューロンとなることが報告されている6).
FruはBTBドメインおよびジンクフィンガーをもつ転写因子である.そこで,Fruと協調してはたらくタンパク質をみつけるため,fru遺伝子をショウジョウバエの複眼に強制発現させて複眼の異常となった個体を作製した.野生型ショウジョウバエの複眼は個眼が規則正しく並んだ半球状のかたちをしているが,fru遺伝子を強制発現したショウジョウバエの複眼は個眼の配列が乱れ小さく細長いかたちになる.この個体に任意の変異をヘテロ接合として導入したとき複眼の表現型を野生型に近づけるような変異をスクリーニングした結果,bonus(bon)変異を見い出した.このbon遺伝子は哺乳類のTIF1ホモログである転写共役因子をコードしていた.TIF1は,ヒストン脱アセチル化酵素HDAC1あるいはヘテロクロマチン制御タンパク質HP1aと,核内受容体との複合体の形成を仲介し,標的遺伝子におけるクロマチンの構造を凝縮させその転写を抑制することが報告されている7).そこで,免疫沈降法により実際にFruがBon,HDAC1,HP1aと複合体を形成しているかどうか調べた結果,FruはFru-Bon-HDAC1複合体あるいはFru-Bon-HP1a複合体を形成しており,Bonを介してFruとHDAC1あるいはFruとHP1aとが結合することが明らかになった.
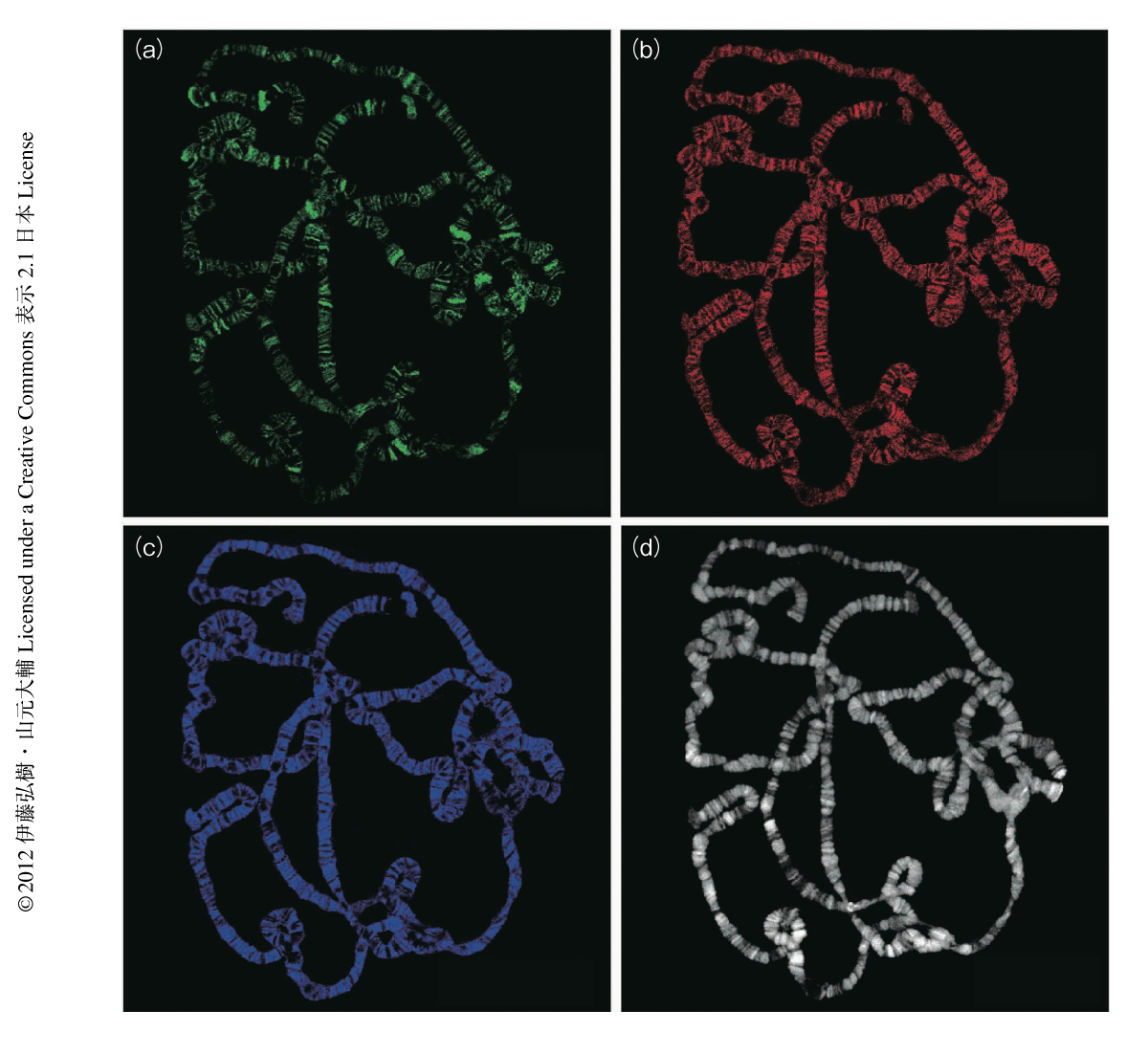
ショウジョウバエ3齢幼虫の唾腺細胞の染色体は多糸染色体を形成しており,染色体における遺伝子座の同定が容易である.この利点を生かし,野生型ショウジョウバエの唾腺染色体をFru,Bon,HDAC1,HP1aそれぞれの抗体を用いて免疫染色し,4本ある染色体(うち,第4染色体はきわめて短い)のどの遺伝子座にFru,Bon,HDAC1,HP1aが結合しているかを調べた(図1).すると,Fruの標的となる遺伝子座は約130あり,そのうち86の遺伝子座においてFru-Bon-HDAC1複合体の結合がみられ,20の遺伝子座においてFru-Bon-HP1a複合体の結合がみられた.3齢幼虫期においてBonを発現しないbon変異体の唾腺染色体を用いて同様の免疫染色を行ったところ,Fruの染色体への結合パターンにまったく変化はなかったが,Fruの標的遺伝子座であるかどうかにかかわらず,HDAC1の染色体への結合はまったくみられなかった.一方,HP1aの染色体へ結合については,Fruの標的遺伝子座において結合はみられず,Fruの標的ではない遺伝子座において結合は維持されていた.さらに,Fruを発現しないfru変異体の唾腺染色体では,Fruの標的遺伝子座においてBon,HDAC1,HP1aの結合はまったくみられなかった.したがって,唾腺染色体の免疫染色による結果も,Fruの標的遺伝子座においてBonを介してFruとHDAC1あるいはFruとHP1aとのあいだで複合体が形成されることを示した.
HDAC1およびHP1aが実際の雄の性行動にはたす役割を調べた.雄の性行動を観察するため,任意の遺伝子型の雄1匹と野生型の処女雌1匹を小さなスペースに入れ,5分間にわたり雄の行動を観察し,雄が定位から交尾の試行までの一連の求愛行動にどのくらいの割合の時間を費やすかを示す求愛指数を測定した.野生型の雄は観察時間の82%を求愛に費やした(つまり,求愛指数は82%).fru遺伝子の弱い欠損型変異体であるfru2/frusat変異体の雄を調べたところその求愛指数は37%に減少した.そこで,HDAC1遺伝子ヌル変異のヘテロ接合を導入したfru2/frusat変異体の雄を調べたところ,その求愛指数は11%とさらに減少した.一方,おもしろいことに,HP1a遺伝子ヌル変異のヘテロ接合を導入したfru2/frusat変異体の雄の求愛指数は53%であって,HP1a遺伝子に変異をもたない場合と比べ16%も上昇していた.この結果は,雄の性行動に対し,HDAC1はFruと同様に促進的にはたらき,HP1aは逆に抑制的にはたらくことを示していた.哺乳類のBonホモログであるTIF1の場合はHDAC1とHP1aのどちらと複合体を形成しても標的遺伝子の転写を抑制することから,ショウジョウバエにおけるこのHP1aの拮抗的なはたらきはまったくの予想外であった.そこで,免疫沈降法により,細胞におけるFru-Bon-HP1a複合体の形成がHDAC1の発現量の増加にしたがい阻害されるのかどうか調べたところ,答えはイエスであった.つまり,Fru-Bon-HDAC1複合体の形成とFru-Bon-HP1a複合体の形成は細胞においても競合関係にあった.
性的二型ニューロンにHDAC1およびHP1aがどのような影響をあたえているのかを調べるためMARCM(mosaic analysis with a repressible cell marker)法による解析を行った8).MARCM法は組換え誘導タンパク質フリッパーゼにより相同染色体における標的配列であるFRT配列を組換えることにより,一部のfru遺伝子の発現ニューロンのみをGFPにより標識することのできる手法であり,それと同時に,標識した細胞のHDAC1あるいはHP1aの発現をノックダウンすることが可能である.ショウジョウバエの胚のニューロブラストとよばれる神経幹細胞は非対称分裂によりニューロブラスト自体と母神経節細胞とをつくり,母神経節細胞はさらに細胞分裂により2つのニューロンをつくりだす.ショウジョウバエの成虫のさまざまなニューロンはこのような細胞分裂を胚期,幼虫期,蛹期のあいだくり返すことによりつくりだされている.したがって,ニューロブラストがつくられる時期に組換えを誘導すると,そののちにつくられる同一の細胞系譜におけるすべてのニューロンを標識することが可能であり,このような同一のニューロブラストから産生された細胞集団をニューロブラストクローンとよぶ.一方,母神経節細胞からニューロンのつくられる時期に組換えを誘導すると単一のニューロンを標識することができる.
脳にあるfru遺伝子の発現ニューロンのうち,フェロモンの情報処理にかかわるニューロンであるmALニューロンには以下の3つの特徴的な性差がみられる9).1)mALニューロンを構成する細胞の数は,雄は30個で,雌は5個.2)細胞体と反対側の脳半球の腹側に延びる樹状突起(反対側の突起)の先端は,雄では馬の尻尾のような房状であるのに対し,雌ではY字型に分岐している.3)雄のmALニューロンには細胞体と同じ脳半球の腹側に延びる樹状突起(同側の突起)をもつものがあるが,雌のmALニューロンには同側の突起をもつものはない.
雄のmALニューロンについてHDAC1をノックダウンしたニューロブラストクローンを観察したところ,細胞の数は21個に減少し,反対側の突起の先端に雌のようなY字型の分岐が現われた.単一のニューロンについて調べたところ,17%のニューロンの樹状突起は完全に雌型に変化しており,そのほかのニューロンでは雄型のままであった.つぎに,fru遺伝子の弱い欠損型変異体であるfru2/fruNP21変異体の雄のニューロブラストクローンを調べたところ,細胞の数は9個と大きく減少し,反対側の突起の先端にも雌のようなY字型の分岐がみられた.単一のニューロンを調べると,雄型あるいは雌型の樹状突起をもつニューロンの割合は1対1と強く雌化が起こっていた.さらに,fru2/fruNP21変異体の雄についてHP1aをノックダウンしたニューロブラストクローンを観察したところ,細胞の数は16個に増加し,単一のニューロンについても雄型と雌型との割合は3対1に変化していた.したがって,HP1aのノックダウンはfru変異による雌化を強く抑制していた.さらにおもしろいことに,いずれの変異体の単一のニューロンに関しても雌雄の中間のかたちの樹状突起をもつものはまったく観察されなかった.また,いずれの変異体も雌の表現型にはまったく異常はなかった.
成虫の前脚にある味覚受容細胞の軸索が腹部神経節に投射するとき,雄の軸索は腹部神経節の正中線をこえて投射するのに対し,雌の軸索は正中線の手前に投射する10).この味覚受容細胞の性差におけるHDAC1およびHP1aの役割を調べるため,軸索をGFPで標識し,同時にHDAC1またはHP1aの発現をノックダウンしてその影響を調べた.正中線を横切る軸索の多寡を定量化するため,野生型の雄の正中線における軸索のGFP蛍光の強度を測定したところ,その値は0.63であった.これに対し,HDAC1をノックダウンした雄の軸索の蛍光強度は0.30と顕著に減少していた.fru遺伝子の弱い欠損型変異体であるfru2/frusat変異体の雄の軸索の蛍光強度は0.02であって正中線を横切る軸索はほとんど観察されなかったが,HP1aをノックダウンしたfru2/frusat変異体の雄の軸索の蛍光強度は0.13となり正中線を横切る軸索の割合が大きく回復した.さらに,いずれの変異体も雌においては正中線を横切る軸索は現われなかった.
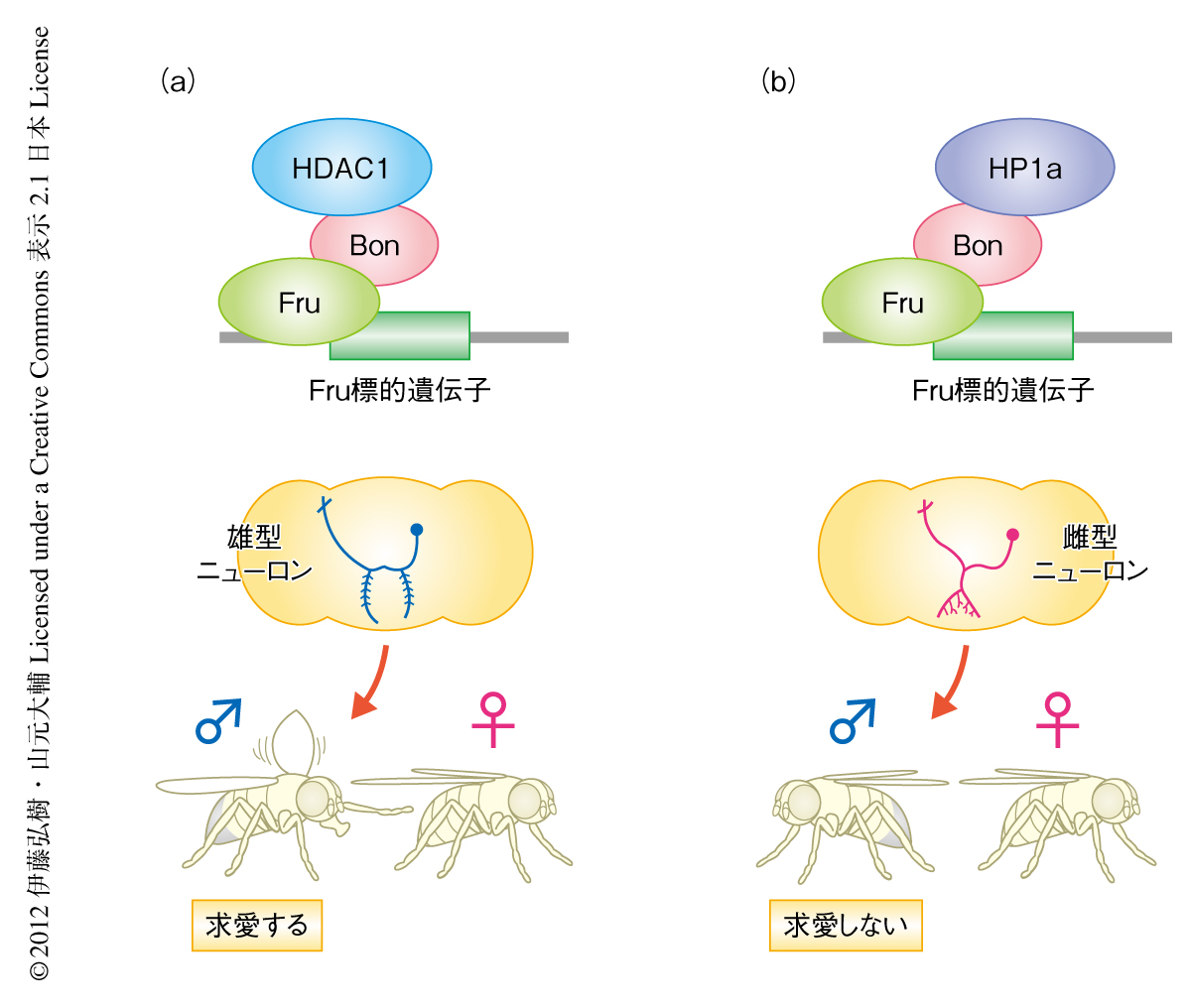
この研究は以下の3つの事実を明らかにした.1)ショウジョウバエの神経系の雄化タンパク質であるFruは転写共役因子Bonを介しヒストン脱アセチル化酵素HDAC1また,ヘテロクロマチン制御タンパク質HP1aと結合するが,HDAC1およびHP1aのFruへの結合は競合関係にある.2)雄の性行動,脳の性的二型ニューロン,味覚受容細胞の性的二型ニューロンの3つの表現型に関して,HDAC1は雄化を誘導するがHP1aはその雄化を阻害する.したがって,BonがFruへHDAC1をリクルートした場合には雄化のスイッチが入り,HP1aをリクルートした場合には雄化のスイッチが切れるものと考えられた(図2).3)mALニューロンを構成する個々の細胞の性をHDAC1およびHP1aは全か無かに制御する.
唾腺染色体の免疫染色の結果(図1)から,Fruの標的遺伝子は約130あり,そのうち少なくとも数個の遺伝子が性的二型ニューロンの形成において重要と思われるが,いまだ標的遺伝子は明らかになっていない.今後,この標的遺伝子を同定するとともに,HDAC1およびHP1aがその標的遺伝子のオンとオフをいかなるタイミングで制御しているかを明らかにすることが,発生の過程においていかに脳の性差がつくられるかを知るため重要と考えられる.
略歴:1993年 北海道大学大学院理学研究科 修了,1994年 科学技術振興事業団ERATO研究員,1998年 同CREST研究員,2005年 理化学研究所発生・再生科学総合研究センター 研究員を経て,2007年より東北大学大学院生命科学研究科 研究員.
研究テーマ:ショウジョウバエの性行動にかかわる遺伝子およびその制御機構と進化.
山元 大輔(Daisuke Yamamoto)
東北大学大学院生命科学研究科 教授.
© 2012 伊藤弘樹・山元大輔 Licensed under CC 表示 2.1 日本
(東北大学大学院生命科学研究科 脳機能遺伝分野)
email:伊藤弘樹,山元大輔
DOI: 10.7875/first.author.2012.076
Fruitless recruits two antagonistic chromatin factors to establish single-neuron sexual dimorphism.
Hiroki Ito, Kosei Sato, Masayuki Koganezawa, Manabu Ote, Ken Matsumoto, Chihiro Hama, Daisuke Yamamoto
Cell, 149, 1327-1338 (2012)
要 約
ショウジョウバエのfruitless遺伝子にコードされるFruitlessは転写因子であり,雄の性行動に必要な神経回路の主要部分を組み立てるはたらきをする.この研究では,Fruitlessは転写共役因子Bonusを介し2種類のクロマチン制御タンパク質HDAC1およびHP1aと複合体を形成するが,BonusがHDAC1をリクルートした場合にはFruitless-Bonus-HDAC1複合体は性的二型ニューロンの雄化を誘導し,BonusがHP1aをリクルートした場合にはFruitless-Bonus-HP1a複合体はそれを阻害することを明らかにした.おもしろいことに,HDAC1あるいはHP1aの発現をノックダウンした場合,性的二型ニューロンを構成する雄型の細胞と雌型の細胞との比率は変化するが,雌雄の中間のかたちをした細胞は決して出現しなかった.したがって,性的二型ニューロンの性は単一の細胞のレベルにおいて全か無かに制御されていると考えられた.
はじめに
ヒトを含め,動物の脳には雌雄の差のあることが40年ほどまえから報告されてきた.たとえば,脳の中心の下部にある視床下部の付近には前視床下部間質核とよばれる領域があり,その第3神経細胞群は男性のほうが大きく細胞の数も多いことが報告されている.さらに,同性愛の男性ではこの部分が小さく女性のものとほぼ等しいという報告もある1).しかし,解析のむずかしさもあり性差を生み出す分子機構はほとんどわかっていない.一方,ショウジョウバエを用いた場合,遺伝学的な手法により個々のニューロンの標識と操作が可能であり,さらに,その性行動は定型的であるため行動を定量解析するのに適している.この研究では,ショウジョウバエの脳の雄化に重要な役割をはたすfruitless(fru)遺伝子について,その産物である転写因子Fruと協調してはたらくタンパク質を探索し,そのタンパク質のはたらきによる標的遺伝子のオンとオフが性差の形成の根源にあることを明らかにした.
1.Fruは雄の性行動を制御する神経回路の形成にはたらき性的二型ニューロンがつくられる
ショウジョウバエの性行動は以下の要素からなる.まず雄が雌に定位(orientation)し,前脚で雌の腹部を叩き(tapping),雌を追いかけながら片方の翅を振ってラブソングを奏でる(singing).雌が立ち止まると雄は雌の背後にまわり口吻で雌の交尾器をなめ(licking),雌の背中に乗り腹部を内側に曲げて交尾を試みる(attempted copulation).雌が性的に受け入れる“気持ち”になると雌は膣口を開き交尾(copulation)がはじまる.雌による交尾の受け入れ以外はすべて雄に特異的な行動であり,野生型の雌は決して雄のような行動はしない.通常,野生型の雄は雌に対し求愛を行い,ほかの雄に対して持続的な求愛をすることはない.
しかし,fru遺伝子の機能を低下させた場合,雄が雄に求愛したり,雄雌のどちらにも求愛しなくなったりする.fru遺伝子のmRNAは性に特異的なスプライシングをうけ,翻訳によりFruタンパク質のつくられるのは雄の神経系のみであり,雌の神経系はFruタンパク質を欠く2-4).近年,ノックイン法を用いたfru遺伝子の改変により雌雄いずれの神経系でもFruのつくられるfruM変異体が作製されているが,おもしろいことに,このfruM変異体の雌はほかの雌に対し雄の行動パターンで求愛を行う5).つまり,個体の性が雄であるか雌であるかに関係なく,fru遺伝子を発現するニューロンにFruをつくらせれば雄の求愛行動を誘起できる.この事実は,Fruが神経系の雄化タンパク質であることを示唆している.ショウジョウバエの脳には約10万個のニューロンがあるが,そのうち2000個のニューロンにfru遺伝子は発現しており,このfru遺伝子の発現ニューロンの一部は細胞の数およびかたちについて性的二型ニューロンとなることが報告されている6).
2.FruはBonを介しFru-Bon-HDAC1複合体あるいはFru-Bon-HP1a複合体を形成する
FruはBTBドメインおよびジンクフィンガーをもつ転写因子である.そこで,Fruと協調してはたらくタンパク質をみつけるため,fru遺伝子をショウジョウバエの複眼に強制発現させて複眼の異常となった個体を作製した.野生型ショウジョウバエの複眼は個眼が規則正しく並んだ半球状のかたちをしているが,fru遺伝子を強制発現したショウジョウバエの複眼は個眼の配列が乱れ小さく細長いかたちになる.この個体に任意の変異をヘテロ接合として導入したとき複眼の表現型を野生型に近づけるような変異をスクリーニングした結果,bonus(bon)変異を見い出した.このbon遺伝子は哺乳類のTIF1ホモログである転写共役因子をコードしていた.TIF1は,ヒストン脱アセチル化酵素HDAC1あるいはヘテロクロマチン制御タンパク質HP1aと,核内受容体との複合体の形成を仲介し,標的遺伝子におけるクロマチンの構造を凝縮させその転写を抑制することが報告されている7).そこで,免疫沈降法により実際にFruがBon,HDAC1,HP1aと複合体を形成しているかどうか調べた結果,FruはFru-Bon-HDAC1複合体あるいはFru-Bon-HP1a複合体を形成しており,Bonを介してFruとHDAC1あるいはFruとHP1aとが結合することが明らかになった.
3.Fruの標的遺伝子座においてFru-Bon-HDAC1複合体あるいはFru-Bon-HP1a複合体が形成される
ショウジョウバエ3齢幼虫の唾腺細胞の染色体は多糸染色体を形成しており,染色体における遺伝子座の同定が容易である.この利点を生かし,野生型ショウジョウバエの唾腺染色体をFru,Bon,HDAC1,HP1aそれぞれの抗体を用いて免疫染色し,4本ある染色体(うち,第4染色体はきわめて短い)のどの遺伝子座にFru,Bon,HDAC1,HP1aが結合しているかを調べた(図1).すると,Fruの標的となる遺伝子座は約130あり,そのうち86の遺伝子座においてFru-Bon-HDAC1複合体の結合がみられ,20の遺伝子座においてFru-Bon-HP1a複合体の結合がみられた.3齢幼虫期においてBonを発現しないbon変異体の唾腺染色体を用いて同様の免疫染色を行ったところ,Fruの染色体への結合パターンにまったく変化はなかったが,Fruの標的遺伝子座であるかどうかにかかわらず,HDAC1の染色体への結合はまったくみられなかった.一方,HP1aの染色体へ結合については,Fruの標的遺伝子座において結合はみられず,Fruの標的ではない遺伝子座において結合は維持されていた.さらに,Fruを発現しないfru変異体の唾腺染色体では,Fruの標的遺伝子座においてBon,HDAC1,HP1aの結合はまったくみられなかった.したがって,唾腺染色体の免疫染色による結果も,Fruの標的遺伝子座においてBonを介してFruとHDAC1あるいはFruとHP1aとのあいだで複合体が形成されることを示した.
4.雄の性行動に対しHDAC1は促進的にはたらきHP1aは抑制的にはたらく
HDAC1およびHP1aが実際の雄の性行動にはたす役割を調べた.雄の性行動を観察するため,任意の遺伝子型の雄1匹と野生型の処女雌1匹を小さなスペースに入れ,5分間にわたり雄の行動を観察し,雄が定位から交尾の試行までの一連の求愛行動にどのくらいの割合の時間を費やすかを示す求愛指数を測定した.野生型の雄は観察時間の82%を求愛に費やした(つまり,求愛指数は82%).fru遺伝子の弱い欠損型変異体であるfru2/frusat変異体の雄を調べたところその求愛指数は37%に減少した.そこで,HDAC1遺伝子ヌル変異のヘテロ接合を導入したfru2/frusat変異体の雄を調べたところ,その求愛指数は11%とさらに減少した.一方,おもしろいことに,HP1a遺伝子ヌル変異のヘテロ接合を導入したfru2/frusat変異体の雄の求愛指数は53%であって,HP1a遺伝子に変異をもたない場合と比べ16%も上昇していた.この結果は,雄の性行動に対し,HDAC1はFruと同様に促進的にはたらき,HP1aは逆に抑制的にはたらくことを示していた.哺乳類のBonホモログであるTIF1の場合はHDAC1とHP1aのどちらと複合体を形成しても標的遺伝子の転写を抑制することから,ショウジョウバエにおけるこのHP1aの拮抗的なはたらきはまったくの予想外であった.そこで,免疫沈降法により,細胞におけるFru-Bon-HP1a複合体の形成がHDAC1の発現量の増加にしたがい阻害されるのかどうか調べたところ,答えはイエスであった.つまり,Fru-Bon-HDAC1複合体の形成とFru-Bon-HP1a複合体の形成は細胞においても競合関係にあった.
5.脳の性的二型ニューロンを構成する個々の細胞の性をHDAC1は雄化するがHP1aはその雄化を阻害する
性的二型ニューロンにHDAC1およびHP1aがどのような影響をあたえているのかを調べるためMARCM(mosaic analysis with a repressible cell marker)法による解析を行った8).MARCM法は組換え誘導タンパク質フリッパーゼにより相同染色体における標的配列であるFRT配列を組換えることにより,一部のfru遺伝子の発現ニューロンのみをGFPにより標識することのできる手法であり,それと同時に,標識した細胞のHDAC1あるいはHP1aの発現をノックダウンすることが可能である.ショウジョウバエの胚のニューロブラストとよばれる神経幹細胞は非対称分裂によりニューロブラスト自体と母神経節細胞とをつくり,母神経節細胞はさらに細胞分裂により2つのニューロンをつくりだす.ショウジョウバエの成虫のさまざまなニューロンはこのような細胞分裂を胚期,幼虫期,蛹期のあいだくり返すことによりつくりだされている.したがって,ニューロブラストがつくられる時期に組換えを誘導すると,そののちにつくられる同一の細胞系譜におけるすべてのニューロンを標識することが可能であり,このような同一のニューロブラストから産生された細胞集団をニューロブラストクローンとよぶ.一方,母神経節細胞からニューロンのつくられる時期に組換えを誘導すると単一のニューロンを標識することができる.
脳にあるfru遺伝子の発現ニューロンのうち,フェロモンの情報処理にかかわるニューロンであるmALニューロンには以下の3つの特徴的な性差がみられる9).1)mALニューロンを構成する細胞の数は,雄は30個で,雌は5個.2)細胞体と反対側の脳半球の腹側に延びる樹状突起(反対側の突起)の先端は,雄では馬の尻尾のような房状であるのに対し,雌ではY字型に分岐している.3)雄のmALニューロンには細胞体と同じ脳半球の腹側に延びる樹状突起(同側の突起)をもつものがあるが,雌のmALニューロンには同側の突起をもつものはない.
雄のmALニューロンについてHDAC1をノックダウンしたニューロブラストクローンを観察したところ,細胞の数は21個に減少し,反対側の突起の先端に雌のようなY字型の分岐が現われた.単一のニューロンについて調べたところ,17%のニューロンの樹状突起は完全に雌型に変化しており,そのほかのニューロンでは雄型のままであった.つぎに,fru遺伝子の弱い欠損型変異体であるfru2/fruNP21変異体の雄のニューロブラストクローンを調べたところ,細胞の数は9個と大きく減少し,反対側の突起の先端にも雌のようなY字型の分岐がみられた.単一のニューロンを調べると,雄型あるいは雌型の樹状突起をもつニューロンの割合は1対1と強く雌化が起こっていた.さらに,fru2/fruNP21変異体の雄についてHP1aをノックダウンしたニューロブラストクローンを観察したところ,細胞の数は16個に増加し,単一のニューロンについても雄型と雌型との割合は3対1に変化していた.したがって,HP1aのノックダウンはfru変異による雌化を強く抑制していた.さらにおもしろいことに,いずれの変異体の単一のニューロンに関しても雌雄の中間のかたちの樹状突起をもつものはまったく観察されなかった.また,いずれの変異体も雌の表現型にはまったく異常はなかった.
6.味覚受容細胞の軸索の投射における性的二型ニューロンについてもHDAC1は雄化するがHP1aはその雄化を阻害する
成虫の前脚にある味覚受容細胞の軸索が腹部神経節に投射するとき,雄の軸索は腹部神経節の正中線をこえて投射するのに対し,雌の軸索は正中線の手前に投射する10).この味覚受容細胞の性差におけるHDAC1およびHP1aの役割を調べるため,軸索をGFPで標識し,同時にHDAC1またはHP1aの発現をノックダウンしてその影響を調べた.正中線を横切る軸索の多寡を定量化するため,野生型の雄の正中線における軸索のGFP蛍光の強度を測定したところ,その値は0.63であった.これに対し,HDAC1をノックダウンした雄の軸索の蛍光強度は0.30と顕著に減少していた.fru遺伝子の弱い欠損型変異体であるfru2/frusat変異体の雄の軸索の蛍光強度は0.02であって正中線を横切る軸索はほとんど観察されなかったが,HP1aをノックダウンしたfru2/frusat変異体の雄の軸索の蛍光強度は0.13となり正中線を横切る軸索の割合が大きく回復した.さらに,いずれの変異体も雌においては正中線を横切る軸索は現われなかった.
おわりに
この研究は以下の3つの事実を明らかにした.1)ショウジョウバエの神経系の雄化タンパク質であるFruは転写共役因子Bonを介しヒストン脱アセチル化酵素HDAC1また,ヘテロクロマチン制御タンパク質HP1aと結合するが,HDAC1およびHP1aのFruへの結合は競合関係にある.2)雄の性行動,脳の性的二型ニューロン,味覚受容細胞の性的二型ニューロンの3つの表現型に関して,HDAC1は雄化を誘導するがHP1aはその雄化を阻害する.したがって,BonがFruへHDAC1をリクルートした場合には雄化のスイッチが入り,HP1aをリクルートした場合には雄化のスイッチが切れるものと考えられた(図2).3)mALニューロンを構成する個々の細胞の性をHDAC1およびHP1aは全か無かに制御する.
唾腺染色体の免疫染色の結果(図1)から,Fruの標的遺伝子は約130あり,そのうち少なくとも数個の遺伝子が性的二型ニューロンの形成において重要と思われるが,いまだ標的遺伝子は明らかになっていない.今後,この標的遺伝子を同定するとともに,HDAC1およびHP1aがその標的遺伝子のオンとオフをいかなるタイミングで制御しているかを明らかにすることが,発生の過程においていかに脳の性差がつくられるかを知るため重要と考えられる.
文 献
- Rahman, Q.: The neurodevelopment of human sexual orientation. Neurosci. Biobehav. Rev., 29, 1057-1066 (2005)[PubMed]
- Ito, H., Fujitani, K., Usui, K. et al.: Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. Proc. Natl. Acad. Sci. USA, 93, 9687-9692 (1996)[PubMed]
- Ryner, L. C., Goodwin, S. F., Castrillon, D. H. et al.: Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell, 87, 1079-1089 (1996)[PubMed]
- Usui-Aoki, K., Ito, H., Ui-Tei, K. et al.: Formation of the male-specific muscle in female Drosophila by ectopic fruitless expression. Nat. Cell Biol., 2, 500-506 (2000)[PubMed]
- Demir, E. & Dickson, B. J.: fruitless splicing specifies male courtship behavior in Drosophila. Cell, 121, 785-794 (2005)[PubMed]
- Kimura, K. -I., Hachiya, T., Koganezawa, M. et al.: Fruitless and doublesex coordinate to generate male-specific neurons that can initiate courtship. Neuron, 59, 759-769 (2008)[PubMed]
- Nielsen, A. L., Ortiz, J. A., You, J. et al.: Interaction with members of the heterochromatin protein 1 (HP1) family and histone deacetylation are differentially involved in transcriptional silencing by members of the TIF1 family. EMBO J., 18, 6385-6395 (1999)[PubMed]
- Lee, T. & Luo, L.: Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron, 22, 451-461 (1999)[PubMed]
- Kimura, K. -I., Ote, M., Tazawa, T. et al.: Fruitless specifies sexually dimorphic neural circuitry in the Drosophila brain. Nature, 438, 229-233 (2005)[PubMed]
- Possidente, D. R. & Murphey, R. K.: Genetic control of sexually dimorphic axon morphology in Drosophila sensory neurons. Dev. Biol., 132, 448-457 (1989)[PubMed]
著者プロフィール
略歴:1993年 北海道大学大学院理学研究科 修了,1994年 科学技術振興事業団ERATO研究員,1998年 同CREST研究員,2005年 理化学研究所発生・再生科学総合研究センター 研究員を経て,2007年より東北大学大学院生命科学研究科 研究員.
研究テーマ:ショウジョウバエの性行動にかかわる遺伝子およびその制御機構と進化.
山元 大輔(Daisuke Yamamoto)
東北大学大学院生命科学研究科 教授.
© 2012 伊藤弘樹・山元大輔 Licensed under CC 表示 2.1 日本
