形質細胞様樹状細胞の生体での炎症反応とT細胞の免疫応答における役割
高木秀明・佐藤克明
(理化学研究所免疫・アレルギー科学総合研究センター 樹状細胞機能研究チーム)
email:佐藤克明
DOI: 10.7875/first.author.2012.001
Plasmacytoid dendritic cells are crucial for the initiation of inflammation and T cell immunity in vivo.
Hideaki Takagi, Tomohiro Fukaya, Kawori Eizumi, Yumiko Sato, Kaori Sato, Azusa Shibazaki, Haruna Otsuka, Atsushi Hijikata, Takashi Watanabe, Osamu Ohara, Tsuneyasu Kaisho, Bernard Malissen, Katsuaki Sato
Immunity, 35, 958-971 (2011)
形質細胞様樹状細胞はエンドソームでのToll様受容体によるリガンドの認識ののち多量にI型インターフェロンを産生する免疫細胞であり,シアル酸をもつ糖鎖を認識するレクチンのひとつSiglec-Hを特異的に発現している.しかしながら,その生体での免疫応答における役割は不明であった.この研究では,Siglec-Hノックアウトマウスおよび形質細胞様樹状細胞を消失させたマウスを用い,形質細胞様樹状細胞が生体での炎症反応とT細胞の免疫応答に重要な役割を担っていることを見い出した.形質細胞様樹状細胞はToll様受容体のリガンドや細菌あるいはウイルスの感染による炎症反応の惹起に必要であった.形質細胞様樹状細胞は抗原免疫や細菌あるいはウイルスの感染においてCD4陽性T細胞の免疫応答の誘導を抑制したが,CD8陽性T細胞の免疫応答の開始に関与していた.さらに,Siglec-Hは形質細胞様樹状細胞におけるこれら免疫応答の調節機能を制御していた.以上の結果から,これまで不明であった形質細胞様樹状細胞の生体での自然免疫応答と獲得免疫応答における役割とその機能制御の機構が明らかになった.
樹状細胞は樹状突起をもつ系統マーカー陰性,MHCクラスII陽性(MHC:major histocompatibility complex,主要組織適合遺伝子複合体)の抗原提示細胞であり,通常型樹状細胞と形質細胞様樹状細胞とに大別される複数のサブセットから構成される1).形質細胞様樹状細胞はウイルス感染により多量のI型インターフェロンを産生する免疫細胞として同定された2,3).通常型樹状細胞とは異なり,エンドソームに存在する核酸受容体であるTLR7とTLR9のみを高発現し(TLR:Toll-like receptor,Toll様受容体),核酸の認識ののちMyD88に依存的なNF-κBの活性化を介して炎症性サイトカインを産生し,IκBキナーゼαとIRF7の活性化を介してI型インターフェロンを産生する4).
形質細胞様樹状細胞はI型インターフェロンの高産生能にもとづくウイルス感染防御への主要なメディエーターと考えられている2,3).また,T細胞の免疫応答において活性化あるいは抑制に多面的な役割を担っているものと推測されている5-7).しかしながら,これらの推察は試験管内での実験や免疫細胞の移入実験による知見にもとづいており,形質細胞様樹状細胞の生体での詳細な免疫学的な役割は不明である.形質細胞様樹状細胞の生体での免疫応答における解析では,形質細胞様樹状細胞で発現の認められるGr-1やBST2に対する抗体を用いた除去法が用いられてきた3).しかしながら,この手法の問題点として,これらのタンパク質はほかの免疫細胞でも発現し,抗Gr-1抗体や抗BST2抗体の投与では形質細胞様樹状細胞を含む多種の免疫細胞が除去されることから,形質細胞様樹状細胞の生体での免疫学的な役割の解釈には齟齬が生じている3).生体での形質細胞様樹状細胞の機能解析はその特異的な除去法がないことによりさまたげられている.
筆者らは,形質細胞様樹状細胞の機能制御の機構を明らかにするため,特異的な発現タンパク質であるSiglec-Hを欠損したSiglec-Hノックアウトマウスを作製した.さらに,形質細胞様樹状細胞の生体での免疫学的な役割を解明するため,Siglec-H遺伝子の一部に自殺遺伝子を導入することにより形質細胞様樹状細胞それ自体が消失した形質細胞様樹状細胞の特異的な消失マウスを作製した.
はじめに,Siglec-Hノックアウトマウスより得た形質細胞様樹状細胞の性状解析を行った.Siglec-Hノックアウト形質細胞様樹状細胞では野生型の形質細胞様樹状細胞と比較して,CD11cとBST2の発現亢進,B220とMHCクラスIIの発現低下が認められた.このことから,Siglec-Hは転写因子の発現の制御により形質細胞様樹状細胞の分化を制御していることが示唆された.また,Siglec-Hノックアウト形質細胞様樹状細胞は野生型と比較して,TLR9のリガンドであるCpG-Aによる刺激ののちにI型インターフェロンおよびインターロイキン12の著しい産生亢進を示した.さらに,Siglec-Hノックアウト形質細胞様樹状細胞ではCpG-Aの刺激によるNF-κBの活性化やIκBキナーゼαおよびIRF7の活性化が増強していた.これらの結果から,Siglec-HはTLR9-MyD88介在性シグナルを阻害することによりTLR9リガンド誘導性のサイトカイン産生を抑制することが考えられた.さらに,Siglec-Hによる形質細胞様樹状細胞のもつT細胞の活性化能の制御について検討した.その結果,Siglec-Hノックアウト形質細胞様樹状細胞は野生型の形質細胞様樹状細胞と比較してCD4陽性T細胞の抗原特異的な活性化能が減弱し,これはMHCクラスIIの低下によることが考えられた.また,Siglec-Hノックアウト形質細胞様樹状細胞は野生型と比較してCD8陽性T細胞の抗原特異的な活性化能も減弱していた.野生型の形質細胞様樹状細胞とSiglec-Hノックアウト形質細胞様樹状細胞は同じ程度のMHCクラスIの発現を示していたことから,これは抗原クロスプレゼンテーション8) の障害によることが考えられた.
形質細胞様樹状細胞のSiglec-Hを介したToll様受容体リガンド誘導性の炎症反応に対する制御について検討した.野生型マウスではCpG-AとD-ガラクトサミンの投与により血清におけるI型インターフェロンや炎症性サイトカインの産生が認められ,投与ののち24時間以内に炎症性ショックにより全例が死亡した.一方,Siglec-Hノックアウトマウスではこれら血清におけるサイトカイン産生の著しい亢進が認められた.しかしながら,形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンのほぼ完全な産生抑制,炎症性サイトカインについては部分的な産生抑制が認められ,死亡率の低下が示された.これらの結果から,形質細胞様樹状細胞は生体においてTLR9リガンドの刺激に対するI型インターフェロンの主要な産生細胞であり,炎症性サイトカインの産生には部分的に寄与していることが示され,Siglec-Hがこれらサイトカインの産生を直接的に制御していることが明らかとなった.また,Toll様受容体リガンド誘導性の通常型樹状細胞の成熟について検討したところ,野生型マウスのもつ通常型樹状細胞におけるMHCクラスIIと共刺激を担うタンパク質の発現の増加と比較して,Siglec-Hノックアウトマウスではこれらのさらなる発現増強が示されたが,形質細胞様樹状細胞の特異的な消失マウスではこれらの発現は低下していた.以上の結果から,通常型樹状細胞におけるToll様受容体リガンド誘導性の成熟機構には形質細胞様樹状細胞によるサイトカインの産生が関与していることが示された.
形質細胞様樹状細胞のSiglec-Hを介したCD4陽性T細胞の抗原特異的な応答に対する制御について検討した.抗原と完全フロイントアジュバントの投与(免疫)ののち,Siglec-Hノックアウトマウスでは野生型マウスと比較してCD4陽性T細胞のより強い抗原特異的な増殖が認められ,形質細胞様樹状細胞の特異的な消失マウスではそのさらなる増強が示された.一方,抗原特異的なインターフェロンγ産生CD4陽性T細胞(Th1細胞)の誘導は,野生型マウスと比較してSiglec-Hノックアウトマウスでは減弱し,形質細胞様樹状細胞の特異的な消失マウスでは増強が認められた.これらの結果から,生体において形質細胞様樹状細胞は,Siglec-Hの関与するMHCクラスII拘束性の抗原提示とサイトカイン産生の制御にもとづきCD4陽性T細胞の抗原特異的な活性化を抑制していることが考えられた.
形質細胞様樹状細胞のSiglec-Hを介したCD8陽性T細胞の抗原特異的な応答への制御について検討した.野生型マウスでは抗原とCpG-Aの投与により抗原特異的なキラーT細胞が産生された.一方,Siglec-Hノックアウトマウスでは抗原特異的なキラーT細胞の産生が低下し,形質細胞様樹状細胞の特異的な消失マウスではさらなる低下が認められた.これらの結果から,生体において形質細胞様樹状細胞はSiglec-Hが関与する抗原クロスプレゼンテーションを介してCD8陽性T細胞の惹起を行い,キラーT細胞の産生に寄与していることが明らかになった.
細菌感染による誘導性の炎症反応に対する形質細胞様樹状細胞の制御についてリステリア感染モデルを用いて検討した.野生型マウスへのリステリアの感染では血清における炎症性サイトカインの高産生を示すとともに感染5日目までに全例が死亡し,細菌感染性の敗血症と類似していた9).一方,形質細胞様樹状細胞の特異的な消失マウスでは野生型マウスと比較して,リステリアの感染ののち炎症性サイトカインの血清における産生の低下と細菌感染性の致死に対する抵抗性を示した.したがって,形質細胞様樹状細胞は初期の重度な細菌感染においてSiglec-Hの制御を介して炎症性サイトカイン産生の惹起および増幅に重要な役割を担い,敗血症ショックを導いていることが考えられた.
さらに,細菌感染での形質細胞様樹状細胞のSiglec-Hを介したT細胞の応答の制御について検討した.野生型マウスへのリステリアの感染ではCD4陽性T細胞やCD8陽性T細胞の抗原特異的な活性化が示され,リステリアに特異的なキラーT細胞の応答が誘導された.一方,Siglec-Hノックアウトマウスや形質細胞様樹状細胞の特異的な消失マウスでは,野生型マウスと比較してCD4陽性T細胞のより強い抗原特異的な活性化が認められた.しかしながら,野生型マウスでのリステリア感染によるCD8陽性T細胞の抗原特異的な活性化と比較して,Siglec-Hノックアウトマウスよりも形質細胞様樹状細胞の特異的な消失マウスではそのより顕著な減弱が認められるとともに,リステリアに特異的なキラーT細胞の応答の低下を示した.これらの結果から,形質細胞様樹状細胞は細菌に対するキラーT細胞の産生を介して細菌感染に対する免疫応答に寄与しているものと考えられた.
形質細胞様樹状細胞のSiglec-Hを介したウイルス感染への免疫応答の制御について,単純ヘルペスI型ウイルス(herpes simplex virus-1:HSV-1)感染モデルを用いて検討した.HSV-1感染によりSiglec-Hノックアウトマウスでは野生型マウスと比較して,血清におけるI型インターフェロンとインターロイキン12の高産生を示し,形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンの著しい産生抑制,インターロイキン12の弱い産生抑制が認められた.また,野生型マウスでのHSV-1感染による特異的なキラーT細胞の産生と比較して,Siglec-Hノックアウトマウスと形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンの産生に対する異なった効果が認められたにもかかわらず,ともに,HSV-1に特異的なキラーT細胞の産生が減弱していた.さらに,野生型マウスでは感染ののち脾臓においてHSV-1は検出されなかったが,形質細胞様樹状細胞の特異的な消失マウスではSiglec-Hノックアウトマウスよりも高いHSV-1感染量を示した.これらの結果から,形質細胞様樹状細胞はI型インターフェロンの産生とウイルス抗原のクロスプレゼンテーションによるウイルスに対するキラーT細胞の産生を介して,ウイルス感染に対する免疫応答に寄与していることが考えられた.
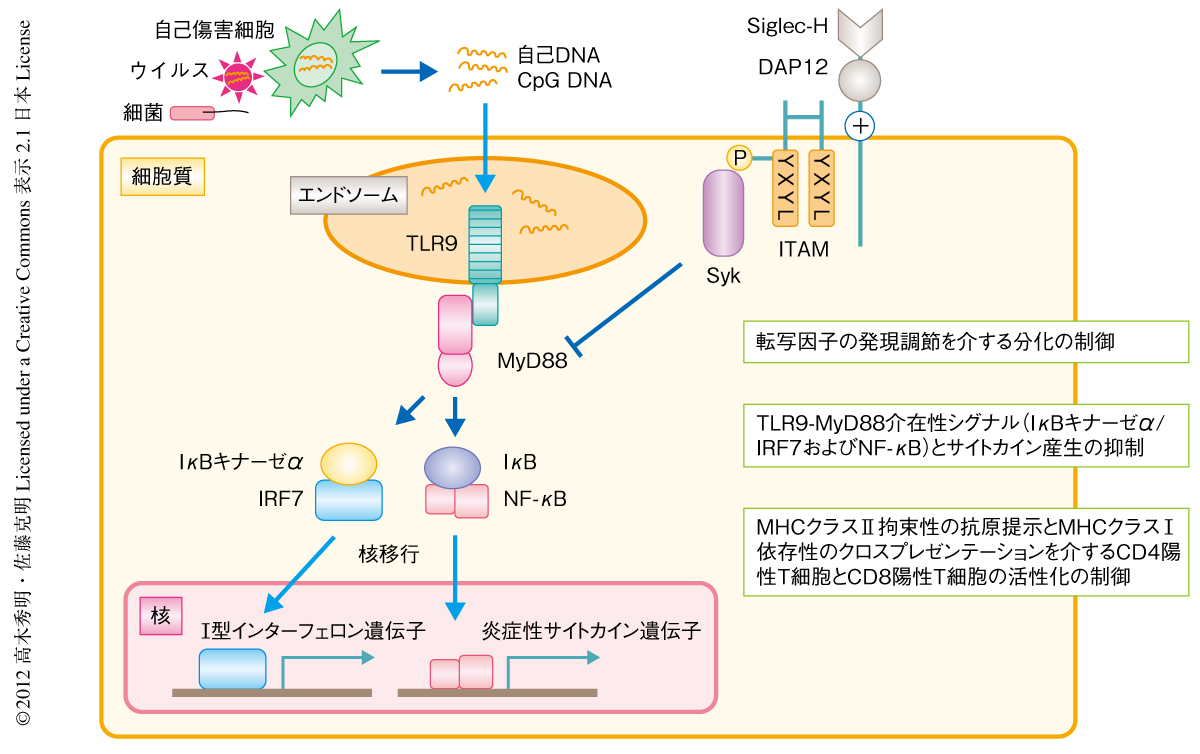
この研究から,Siglec-Hは形質細胞様樹状細胞の分化,Toll様受容体リガンド誘導性のサイトカインの産生,T細胞の活性化を制御していることが明らかになった(図1).自然免疫応答では形質細胞様樹状細胞ははじめに細菌やウイルスを感知してI型インターフェロンや炎症性サイトカインを産生することで炎症反応を惹起し,通常型樹状細胞などのほかの免疫細胞を活性化して炎症反応を亢進していることをつきとめた(図2a).また,細菌やウイルスの感染に対する獲得免疫応答では形質細胞様樹状細胞はCD4陽性エフェクターT細胞の誘導を抑制するが,キラーT細胞を積極的に産生することによりこれら病原性微生物やその感染細胞を生体から効率的に排除していることを解明した(図2b).したがって,この研究の成果を応用することで感染症に対する新しい治療法の開発につながる可能性が期待される.今後は,自己免疫疾患の発症や増悪における形質細胞様樹状細胞の役割とその制御機構を解明したい.

略歴:千葉大学大学院医学薬学府博士課程 在学中.
研究テーマ:形質細胞様樹状細胞の免疫応答における役割と機能制御の機構.
佐藤 克明(Katsuaki Sato)
理化学研究所免疫・アレルギー科学総合研究センター チームリーダー.
研究室URL:http://www.riken.jp/r-world/research/lab/rcai/cell/index.html
© 2012 高木秀明・佐藤克明 Licensed under CC 表示 2.1 日本
(理化学研究所免疫・アレルギー科学総合研究センター 樹状細胞機能研究チーム)
email:佐藤克明
DOI: 10.7875/first.author.2012.001
Plasmacytoid dendritic cells are crucial for the initiation of inflammation and T cell immunity in vivo.
Hideaki Takagi, Tomohiro Fukaya, Kawori Eizumi, Yumiko Sato, Kaori Sato, Azusa Shibazaki, Haruna Otsuka, Atsushi Hijikata, Takashi Watanabe, Osamu Ohara, Tsuneyasu Kaisho, Bernard Malissen, Katsuaki Sato
Immunity, 35, 958-971 (2011)
この論文に出現する遺伝子・タンパク質のUniprot ID
Toll様受容体, I型インターフェロン, Siglec-H(Q3Y597), CD4(P06332), CD8, MHCクラスII陽性, MHC, major histocompatibility complex, 主要組織適合遺伝子複合体, TLR7(P58681), TLR9(Q9EQU3), TLR, Toll-like receptor, MyD88(P22366), NF-κB, 炎症性サイトカイン, IκBキナーゼα(Q60680), IRF7(P70434), Gr-1, BST2(Q8R2Q8), CD11c(Q9QXH4), B220(P06800), MHCクラスII, インターロイキン12, サイトカイン, MHCクラスI, インターフェロンγ(P01580)
要 約
形質細胞様樹状細胞はエンドソームでのToll様受容体によるリガンドの認識ののち多量にI型インターフェロンを産生する免疫細胞であり,シアル酸をもつ糖鎖を認識するレクチンのひとつSiglec-Hを特異的に発現している.しかしながら,その生体での免疫応答における役割は不明であった.この研究では,Siglec-Hノックアウトマウスおよび形質細胞様樹状細胞を消失させたマウスを用い,形質細胞様樹状細胞が生体での炎症反応とT細胞の免疫応答に重要な役割を担っていることを見い出した.形質細胞様樹状細胞はToll様受容体のリガンドや細菌あるいはウイルスの感染による炎症反応の惹起に必要であった.形質細胞様樹状細胞は抗原免疫や細菌あるいはウイルスの感染においてCD4陽性T細胞の免疫応答の誘導を抑制したが,CD8陽性T細胞の免疫応答の開始に関与していた.さらに,Siglec-Hは形質細胞様樹状細胞におけるこれら免疫応答の調節機能を制御していた.以上の結果から,これまで不明であった形質細胞様樹状細胞の生体での自然免疫応答と獲得免疫応答における役割とその機能制御の機構が明らかになった.
はじめに
樹状細胞は樹状突起をもつ系統マーカー陰性,MHCクラスII陽性(MHC:major histocompatibility complex,主要組織適合遺伝子複合体)の抗原提示細胞であり,通常型樹状細胞と形質細胞様樹状細胞とに大別される複数のサブセットから構成される1).形質細胞様樹状細胞はウイルス感染により多量のI型インターフェロンを産生する免疫細胞として同定された2,3).通常型樹状細胞とは異なり,エンドソームに存在する核酸受容体であるTLR7とTLR9のみを高発現し(TLR:Toll-like receptor,Toll様受容体),核酸の認識ののちMyD88に依存的なNF-κBの活性化を介して炎症性サイトカインを産生し,IκBキナーゼαとIRF7の活性化を介してI型インターフェロンを産生する4).
形質細胞様樹状細胞はI型インターフェロンの高産生能にもとづくウイルス感染防御への主要なメディエーターと考えられている2,3).また,T細胞の免疫応答において活性化あるいは抑制に多面的な役割を担っているものと推測されている5-7).しかしながら,これらの推察は試験管内での実験や免疫細胞の移入実験による知見にもとづいており,形質細胞様樹状細胞の生体での詳細な免疫学的な役割は不明である.形質細胞様樹状細胞の生体での免疫応答における解析では,形質細胞様樹状細胞で発現の認められるGr-1やBST2に対する抗体を用いた除去法が用いられてきた3).しかしながら,この手法の問題点として,これらのタンパク質はほかの免疫細胞でも発現し,抗Gr-1抗体や抗BST2抗体の投与では形質細胞様樹状細胞を含む多種の免疫細胞が除去されることから,形質細胞様樹状細胞の生体での免疫学的な役割の解釈には齟齬が生じている3).生体での形質細胞様樹状細胞の機能解析はその特異的な除去法がないことによりさまたげられている.
筆者らは,形質細胞様樹状細胞の機能制御の機構を明らかにするため,特異的な発現タンパク質であるSiglec-Hを欠損したSiglec-Hノックアウトマウスを作製した.さらに,形質細胞様樹状細胞の生体での免疫学的な役割を解明するため,Siglec-H遺伝子の一部に自殺遺伝子を導入することにより形質細胞様樹状細胞それ自体が消失した形質細胞様樹状細胞の特異的な消失マウスを作製した.
1.形質細胞様樹状細胞の機能制御におけるSiglec-Hの役割
はじめに,Siglec-Hノックアウトマウスより得た形質細胞様樹状細胞の性状解析を行った.Siglec-Hノックアウト形質細胞様樹状細胞では野生型の形質細胞様樹状細胞と比較して,CD11cとBST2の発現亢進,B220とMHCクラスIIの発現低下が認められた.このことから,Siglec-Hは転写因子の発現の制御により形質細胞様樹状細胞の分化を制御していることが示唆された.また,Siglec-Hノックアウト形質細胞様樹状細胞は野生型と比較して,TLR9のリガンドであるCpG-Aによる刺激ののちにI型インターフェロンおよびインターロイキン12の著しい産生亢進を示した.さらに,Siglec-Hノックアウト形質細胞様樹状細胞ではCpG-Aの刺激によるNF-κBの活性化やIκBキナーゼαおよびIRF7の活性化が増強していた.これらの結果から,Siglec-HはTLR9-MyD88介在性シグナルを阻害することによりTLR9リガンド誘導性のサイトカイン産生を抑制することが考えられた.さらに,Siglec-Hによる形質細胞様樹状細胞のもつT細胞の活性化能の制御について検討した.その結果,Siglec-Hノックアウト形質細胞様樹状細胞は野生型の形質細胞様樹状細胞と比較してCD4陽性T細胞の抗原特異的な活性化能が減弱し,これはMHCクラスIIの低下によることが考えられた.また,Siglec-Hノックアウト形質細胞様樹状細胞は野生型と比較してCD8陽性T細胞の抗原特異的な活性化能も減弱していた.野生型の形質細胞様樹状細胞とSiglec-Hノックアウト形質細胞様樹状細胞は同じ程度のMHCクラスIの発現を示していたことから,これは抗原クロスプレゼンテーション8) の障害によることが考えられた.
2.形質細胞様樹状細胞による炎症反応の制御
形質細胞様樹状細胞のSiglec-Hを介したToll様受容体リガンド誘導性の炎症反応に対する制御について検討した.野生型マウスではCpG-AとD-ガラクトサミンの投与により血清におけるI型インターフェロンや炎症性サイトカインの産生が認められ,投与ののち24時間以内に炎症性ショックにより全例が死亡した.一方,Siglec-Hノックアウトマウスではこれら血清におけるサイトカイン産生の著しい亢進が認められた.しかしながら,形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンのほぼ完全な産生抑制,炎症性サイトカインについては部分的な産生抑制が認められ,死亡率の低下が示された.これらの結果から,形質細胞様樹状細胞は生体においてTLR9リガンドの刺激に対するI型インターフェロンの主要な産生細胞であり,炎症性サイトカインの産生には部分的に寄与していることが示され,Siglec-Hがこれらサイトカインの産生を直接的に制御していることが明らかとなった.また,Toll様受容体リガンド誘導性の通常型樹状細胞の成熟について検討したところ,野生型マウスのもつ通常型樹状細胞におけるMHCクラスIIと共刺激を担うタンパク質の発現の増加と比較して,Siglec-Hノックアウトマウスではこれらのさらなる発現増強が示されたが,形質細胞様樹状細胞の特異的な消失マウスではこれらの発現は低下していた.以上の結果から,通常型樹状細胞におけるToll様受容体リガンド誘導性の成熟機構には形質細胞様樹状細胞によるサイトカインの産生が関与していることが示された.
3.形質細胞様樹状細胞によるCD4陽性T細胞の抗原特異的な応答の制御
形質細胞様樹状細胞のSiglec-Hを介したCD4陽性T細胞の抗原特異的な応答に対する制御について検討した.抗原と完全フロイントアジュバントの投与(免疫)ののち,Siglec-Hノックアウトマウスでは野生型マウスと比較してCD4陽性T細胞のより強い抗原特異的な増殖が認められ,形質細胞様樹状細胞の特異的な消失マウスではそのさらなる増強が示された.一方,抗原特異的なインターフェロンγ産生CD4陽性T細胞(Th1細胞)の誘導は,野生型マウスと比較してSiglec-Hノックアウトマウスでは減弱し,形質細胞様樹状細胞の特異的な消失マウスでは増強が認められた.これらの結果から,生体において形質細胞様樹状細胞は,Siglec-Hの関与するMHCクラスII拘束性の抗原提示とサイトカイン産生の制御にもとづきCD4陽性T細胞の抗原特異的な活性化を抑制していることが考えられた.
4.形質細胞様樹状細胞によるCD8陽性T細胞の抗原特異的な応答の制御
形質細胞様樹状細胞のSiglec-Hを介したCD8陽性T細胞の抗原特異的な応答への制御について検討した.野生型マウスでは抗原とCpG-Aの投与により抗原特異的なキラーT細胞が産生された.一方,Siglec-Hノックアウトマウスでは抗原特異的なキラーT細胞の産生が低下し,形質細胞様樹状細胞の特異的な消失マウスではさらなる低下が認められた.これらの結果から,生体において形質細胞様樹状細胞はSiglec-Hが関与する抗原クロスプレゼンテーションを介してCD8陽性T細胞の惹起を行い,キラーT細胞の産生に寄与していることが明らかになった.
5.形質細胞様樹状細胞の細菌感染に対する免疫応答における役割
細菌感染による誘導性の炎症反応に対する形質細胞様樹状細胞の制御についてリステリア感染モデルを用いて検討した.野生型マウスへのリステリアの感染では血清における炎症性サイトカインの高産生を示すとともに感染5日目までに全例が死亡し,細菌感染性の敗血症と類似していた9).一方,形質細胞様樹状細胞の特異的な消失マウスでは野生型マウスと比較して,リステリアの感染ののち炎症性サイトカインの血清における産生の低下と細菌感染性の致死に対する抵抗性を示した.したがって,形質細胞様樹状細胞は初期の重度な細菌感染においてSiglec-Hの制御を介して炎症性サイトカイン産生の惹起および増幅に重要な役割を担い,敗血症ショックを導いていることが考えられた.
さらに,細菌感染での形質細胞様樹状細胞のSiglec-Hを介したT細胞の応答の制御について検討した.野生型マウスへのリステリアの感染ではCD4陽性T細胞やCD8陽性T細胞の抗原特異的な活性化が示され,リステリアに特異的なキラーT細胞の応答が誘導された.一方,Siglec-Hノックアウトマウスや形質細胞様樹状細胞の特異的な消失マウスでは,野生型マウスと比較してCD4陽性T細胞のより強い抗原特異的な活性化が認められた.しかしながら,野生型マウスでのリステリア感染によるCD8陽性T細胞の抗原特異的な活性化と比較して,Siglec-Hノックアウトマウスよりも形質細胞様樹状細胞の特異的な消失マウスではそのより顕著な減弱が認められるとともに,リステリアに特異的なキラーT細胞の応答の低下を示した.これらの結果から,形質細胞様樹状細胞は細菌に対するキラーT細胞の産生を介して細菌感染に対する免疫応答に寄与しているものと考えられた.
6.形質細胞様樹状細胞のウイルス感染に対する免疫応答における役割
形質細胞様樹状細胞のSiglec-Hを介したウイルス感染への免疫応答の制御について,単純ヘルペスI型ウイルス(herpes simplex virus-1:HSV-1)感染モデルを用いて検討した.HSV-1感染によりSiglec-Hノックアウトマウスでは野生型マウスと比較して,血清におけるI型インターフェロンとインターロイキン12の高産生を示し,形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンの著しい産生抑制,インターロイキン12の弱い産生抑制が認められた.また,野生型マウスでのHSV-1感染による特異的なキラーT細胞の産生と比較して,Siglec-Hノックアウトマウスと形質細胞様樹状細胞の特異的な消失マウスではI型インターフェロンの産生に対する異なった効果が認められたにもかかわらず,ともに,HSV-1に特異的なキラーT細胞の産生が減弱していた.さらに,野生型マウスでは感染ののち脾臓においてHSV-1は検出されなかったが,形質細胞様樹状細胞の特異的な消失マウスではSiglec-Hノックアウトマウスよりも高いHSV-1感染量を示した.これらの結果から,形質細胞様樹状細胞はI型インターフェロンの産生とウイルス抗原のクロスプレゼンテーションによるウイルスに対するキラーT細胞の産生を介して,ウイルス感染に対する免疫応答に寄与していることが考えられた.
おわりに
この研究から,Siglec-Hは形質細胞様樹状細胞の分化,Toll様受容体リガンド誘導性のサイトカインの産生,T細胞の活性化を制御していることが明らかになった(図1).自然免疫応答では形質細胞様樹状細胞ははじめに細菌やウイルスを感知してI型インターフェロンや炎症性サイトカインを産生することで炎症反応を惹起し,通常型樹状細胞などのほかの免疫細胞を活性化して炎症反応を亢進していることをつきとめた(図2a).また,細菌やウイルスの感染に対する獲得免疫応答では形質細胞様樹状細胞はCD4陽性エフェクターT細胞の誘導を抑制するが,キラーT細胞を積極的に産生することによりこれら病原性微生物やその感染細胞を生体から効率的に排除していることを解明した(図2b).したがって,この研究の成果を応用することで感染症に対する新しい治療法の開発につながる可能性が期待される.今後は,自己免疫疾患の発症や増悪における形質細胞様樹状細胞の役割とその制御機構を解明したい.
文 献
- Shortman, K. & Naik, S. H.: Steady-state and inflammatory dendritic-cell development. Nat. Rev. Immunol., 7, 19-30 (2007)[PubMed]
- Gilliet, M., Cao, W. & Liu, Y. J.: Plasmacytoid dendritic cells: sensing nucleic acids in viral infection and autoimmune diseases. Nat. Rev. Immunol., 8, 594-606 (2008)[PubMed]
- Swiecki, M. & Colonna, M.: Unraveling the functions of plasmacytoid dendritic cells during viral infections, autoimmunity, and tolerance. Immunol. Rev., 234, 142-162 (2010)[PubMed]
- Hoshino, K., Sugiyama, T., Matsumoto, M. et al.: IκB kinase-α is critical for interferon-α production induced by Toll-like receptors 7 and 9. Nature, 440, 949-953 (2006)[PubMed]
- Salio, M., Palmowski, M. J., Atzberger, A. et al.: CpG-matured murine plasmacytoid dendritic cells are capable of in vivo priming of functional CD8 T cell responses to endogenous but not exogenous antigens. J. Exp. Med., 199, 567-579 (2004)[PubMed]
- Ochando, J. C., Homma, C., Yang, Y. et al.: Alloantigen-presenting plasmacytoid dendritic cells mediate tolerance to vascularized grafts. Nat. Immunol., 6, 652-662 (2006)[PubMed]
- Irla, M., Kupfer, N., Suter, T. et al.: MHC class II-restricted antigen presentation by plasmacytoid dendritic cells inhibits T cell-mediated autoimmunity. J. Exp. Med., 207, 1891-1905 (2010)[PubMed]
- Villadangos, J. A. & Young, L.: Antigen-presentation properties of plasmacytoid dendritic cells. Immunity, 29, 352-361 (2008)[PubMed]
- Fujita, S., Seino, K., Sato, K. et al.: Regulatory dendritic cells act as regulators of acute lethal systemic inflammatory response. Blood, 107, 3656-3664 (2006)[PubMed]
著者プロフィール
略歴:千葉大学大学院医学薬学府博士課程 在学中.
研究テーマ:形質細胞様樹状細胞の免疫応答における役割と機能制御の機構.
佐藤 克明(Katsuaki Sato)
理化学研究所免疫・アレルギー科学総合研究センター チームリーダー.
研究室URL:http://www.riken.jp/r-world/research/lab/rcai/cell/index.html
© 2012 高木秀明・佐藤克明 Licensed under CC 表示 2.1 日本
