転写メディエーター複合体のヘッドモジュールの構造
今崎 剛・高木雄一郎
(米国Indiana大学School of Medicine,Department of Biochemistry and Molecular Biology)
email:今崎 剛,高木雄一郎
DOI: 10.7875/first.author.2011.119
Architecture of the Mediator head module.
Tsuyoshi Imasaki, Guillermo Calero, Gang Cai, Kuang-Lei Tsai, Kentaro Yamada, Francesco Cardelli, Hediye Erdjument-Bromage, Paul Tempst, Imre Berger, Guy Lorch Kornberg, Francisco J. Asturias, Roger D. Kornberg, Yuichiro Takagi
Nature, 475, 240-243 (2011)
転写メディエーター複合体は遺伝子の発現機構において重要な役割をはたし,エンハンサーからの情報をRNAポリメラーゼIIへと伝達するのがその役割である.その構造および機能は酵母からヒトまで保存されており,酵母では21個のサブユニットからなる超分子複合体を形成し,ヘッドモジュール,ミドル/アームモジュール,テイルモジュールと名づけられた3つの独立したモジュールから構成されている.転写機構の解明において転写メディエーター複合体の機能および構造の解析は必須であるが,転写メディエーター複合体は巨大かつ複雑であるため研究は困難をきたしていた.今回,筆者らは,転写メディエーター複合体において主要な機能をつかさどるヘッドモジュールのX線結晶構造解析を行い,分解能4.3Åでの構造の決定に成功した.このヘッドモジュールは7つのサブユニットが複雑にからみあうというまったく新規な構造をとっていた.そのことから3つのドメインからなる柔軟性と安定性とをもちあわせ,RNAポリメラーゼIIおよび基本転写因子との複雑な相互作用を可能にしていることを解明した.
転写は遺伝子の発現においてもっとも重要な生命現象のひとつであり,その分子機構の解明は生命科学における重要な課題である.真核生物ではRNAポリメラーゼIIがすべてのmRNAの合成を行っている.このRNAポリメラーゼIIは遺伝子の上流の領域に結合するエンハンサー,あるいは,基本転写因子などさまざまな因子により制御されている.超分子複合体である転写メディエーター複合体はこれらを機能的に結びつけており,転写を理解するうえでもっとも重要な因子のひとつである1).転写メディエーター複合体は酵母からヒトにいたるまで機能的および構造的に保存されている.出芽酵母では21個のサブユニットからなり,ヘッドモジュール,ミドル/アームモジュール,テイルモジュールと名づけられた3つの独立したモジュールから構成されている2).しかし,多数のサブユニットから構成されるという複雑さ,細胞における発現量の少なさ,1 MDaをこえる大きさから構造研究は難航しており,分子機構についての詳細はほとんどわかっていない.
筆者らは,数々の最新技術を導入することにより,転写メディエーター複合体の主要な機能をつかさどるヘッドモジュール3) の構造決定に成功した.驚くべきことに,このヘッドモジュールは7つのサブユニットが複雑にからみあい3つのドメインを形成して安定性と柔軟性とをあわせもつという構造をとっていた.構造解析,生化学的な実験,遺伝学的な解析,そして,過去の研究報告から,転写メディエーター複合体のヘッドモジュールはRNAポリメラーゼIIや基本転写因子と相互作用し,それぞれを適切な位置に配置する調整役として機能する機構を明らかにした.
超分子複合体の構造解析はその大きさ,複雑さ,結晶化試料の大量調製のむずかしさ,高品質な結晶調製の困難さ,などさまざまな問題があることから難解である.現在までに構造解析された超分子複合体は,リボソーム,RNAポリメラーゼIIを含め,天然に発現したタンパク質から調製した試料を用いた例がほとんどである.しかし,転写メディエーター複合体の発現量は出芽酵母を100リットル培養しても精製ののち数mgと,この方法で30 mg程度は得られるRNAポリメラーゼIIなどと比べ非常に低く,結晶構造解析には組換えタンパク質による大量発現法を用いることが必須であった.そこで,バキュロウイルス-昆虫細胞を用いた組換えタンパク質発現系を用い,転写メディエーター複合体の主要な機能をつかさどるヘッドモジュールを発現させた.超分子複合体の発現のため開発されたMultiBacシステム4) と,複数の遺伝子を同時にクローニングすることのできるSLIC(sequence and ligation independent iloning)法5) とを組み合わせ,高効率な超分子複合体の発現系を実現した.この発現系を最大限に利用することで,64種類にものぼるコンストラクトについてスクリーニングを行った結果,結晶構造解析に適した結晶を得ることに成功し,それを用いてX線結晶構造解析を行った.不運にもこの結晶は単結晶ではなく双晶(merohedral twin)であったため,それをできるだけ単結晶に修正する条件の検索に多大な時間を費やすことになった.
構造解析は重原子クラスターを用いた初期位相の決定とセレノメチオニン置換タンパク質の単波長異常分散法とを組み合わせ,初期電子密度を分解能4.3Åで得ることに成功した.この低い分解能のため,低分解能でもモデル構築のしやすい領域のモデル構築をまず行い,さらにセレノメチオニンに由来するメチオニン残基の位置,2次構造の予測,サイズの大きい芳香族側鎖の位置を目印にアミノ酸残基の位置決定を行った.その結果,7つのサブユニットからなる超分子複合体,転写メディエーター複合体のヘッドモジュールの結晶構造を分解能4.3Åで決定することに成功した(図1,PDB ID:3RJ1).
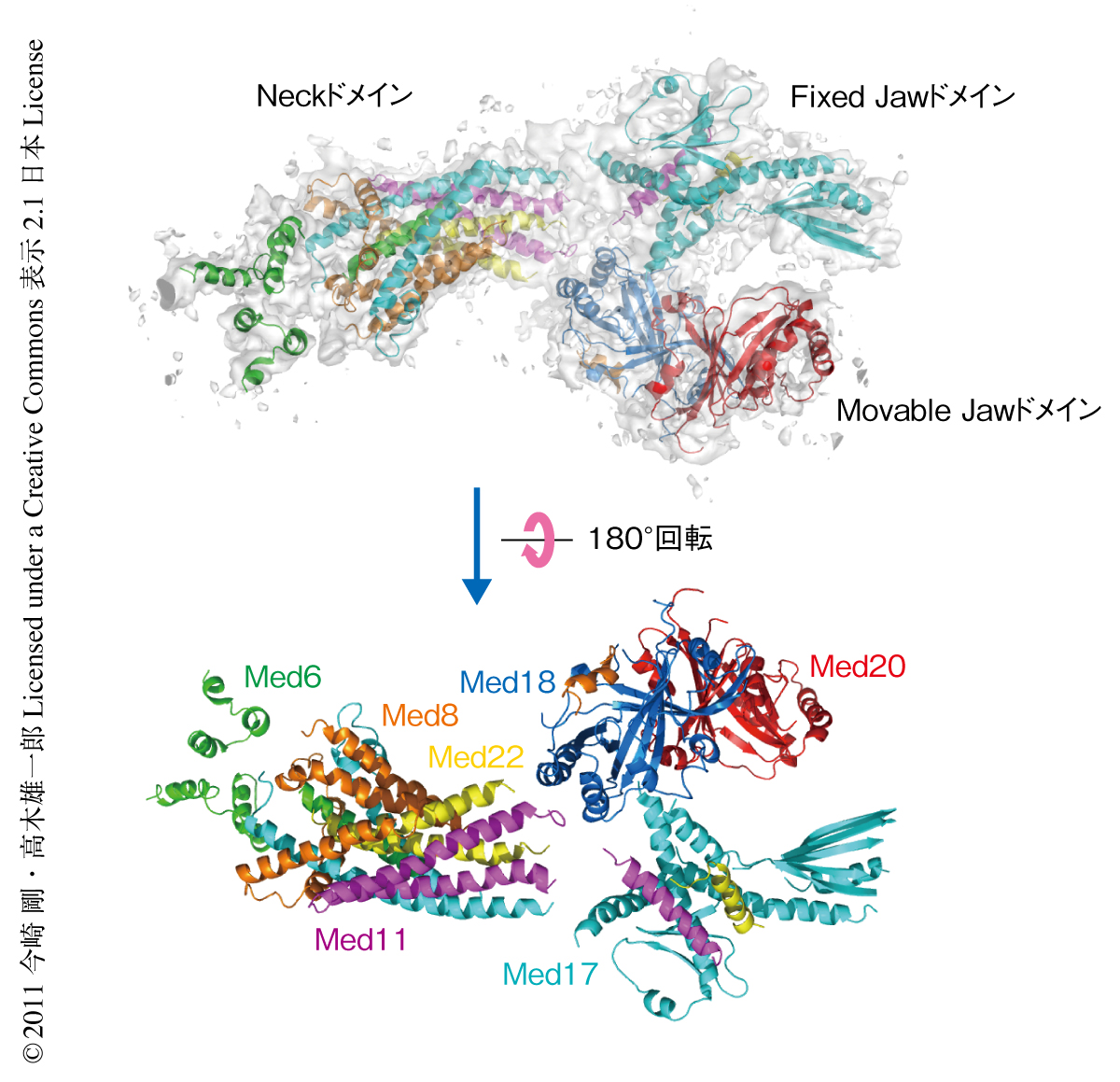
ヘッドモジュールの全体構造は,以前,筆者らのグループが報告した電子線顕微鏡による構造解析の結果と同じく,アルファベットのYのようなかたちをしていた6)(図1).7つのサブユニットが複雑にからみあい3つの独立したドメインを形成しており,ドメインどうしは柔軟なリンカー領域でつながれていた.この3つのドメインをNeckドメイン,Fixed Jawドメイン,Movable Jawドメインと名づけた.柔軟なリンカーでつながれていることは,電子線顕微鏡による構造解析においてMovable Jawドメインがいくつかの異なったコンフォメーションをとることを非常によく説明することができた.
複合体の形成機構の謎はNeckドメインのユニークな構造にかくされていた.Neckドメインは5種類のサブユニットからなる10本のαヘリカルバンドルをコアとした非常にユニークな構造をとっていた(図2a).αヘリカルバンドル構造は膜タンパク質などによくみられそれ自体に目新しいことはない.ところが驚くべきことに,Neckドメインの場合,10本のαヘリカルバンドルが5種類のまったく違ったサブユニットから構成されていた.この構造をさらにくわしくみると,NeckドメインにおいてはまずMed11-Med17-Med22からなる5本の長いαヘリカルバンドル構造が安定な相互作用をつくり,そこにMed6-Med8がからみつくようにして複合体のあいだの相互作用を形成していた.また,転写メディエーター複合体のヘッドモジュールは基本転写因子TFIIDのサブユニットのひとつ,TBP(TATA binding protein)やTFIIHとの相互作用が報告されている7,8).これらの相互作用の領域はNeckドメインにあり,このことから,Neckドメインは複合体の形成のみならずタンパク質間相互作用にも重要な役割をもつことが示唆された.
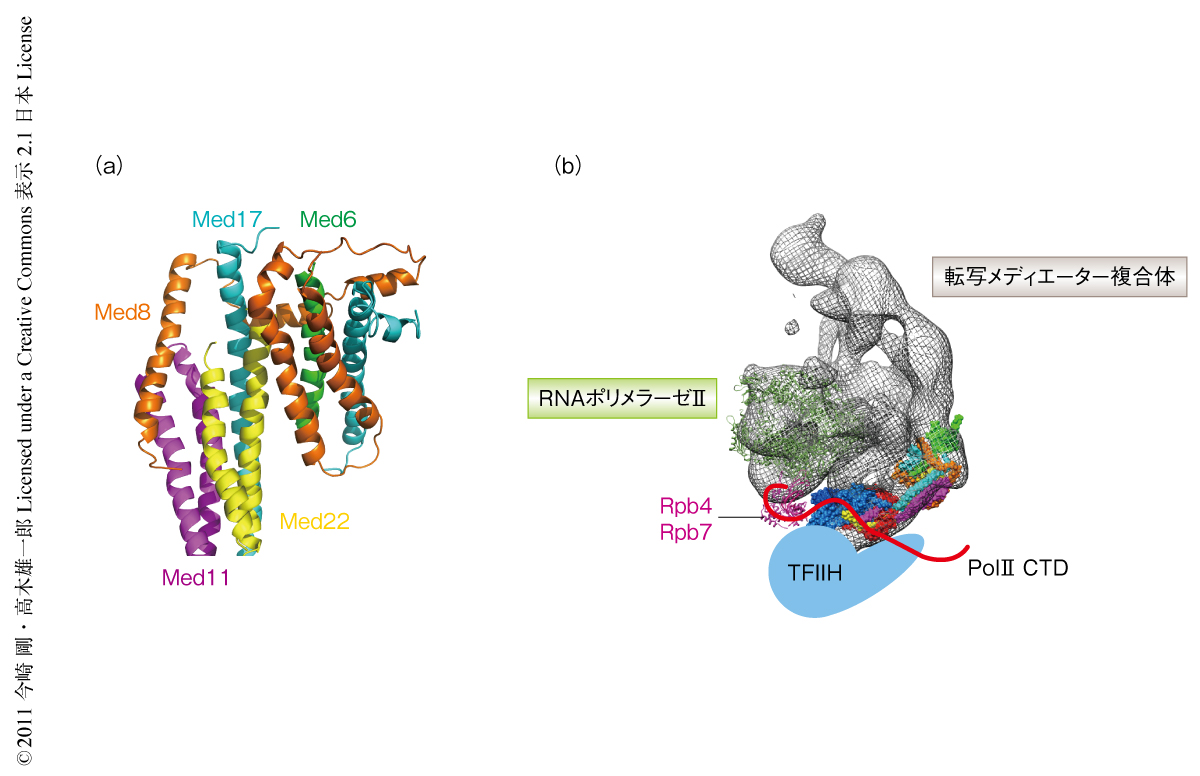
近年,転写の過程において転写開始から鎖伸長に切り替わる過程が幹細胞の自己複製に重要な役割をはたしていることが明らかになった9).この過程では,RNAポリメラーゼIIのもつC末端ドメインが基本転写因子TFIIHによりリン酸化をうけることが必要であり,このリン酸化は転写メディエーター複合体のRNAポリメラーゼIIからの解離も促進し,転写の過程が転写開始から鎖伸長へと切り替わるのに重要な役割をはたしている10).さらに,転写メディエーター複合体はTFIIHによるRNAポリメラーゼIIのC末端ドメインSer5のリン酸化を促進することが報告されている11).しかし,RNAポリメラーゼIIのC末端ドメイン,転写メディエーター複合体,そして,TFIIHとのあいだの相互作用の機構は謎につつまれていた.この研究で得られた転写メディエーター複合体の構造情報と過去の研究報告とをあわせると,ヘッドモジュールはRNAポリメラーゼIIとTFIIHをつなぐ架け橋となり,それぞれの位置を安定化し転写開始から鎖伸長への切り替えを促進しているのではないかと考えられた(図2b).
転写メディエーター複合体のヘッドモジュールの構造決定は想像以上に困難であったが,最先端のタンパク質発現技術とX線構造解析法とを駆使することでこれに成功し,構造生物学の分野にブレークスルーをもたらすことができた.驚くべきことに,転写メディエーター複合体のヘッドモジュールは7つのサブユニットが複雑にからみあった構造を形成することで安定性と柔軟性とをあわせもつという,過去に観測されたことのない新規な構造をとっていた.このような構造はそれぞれのサブユニットをターゲットとした従来の構造解析法により解明することは不可能であり,超分子複合体の全体をターゲットとしたことではじめて明らかになった成果である.この研究は,超分子複合体は複合体として研究しなければその実態はわからないという好例であった.細胞には数多くの超分子複合体が存在し重要なはたらきをしており,これら超分子複合体の構造および機能の研究は今後の重要課題である.また,超分子複合体の構造研究が基礎研究のみならず,超分子複合体をターゲットにしたドラッグデザインなどの応用に重要な役割をはたすことは明らかであろう.
米国Indiana大学:http://communications.medicine.iu.edu/newsroom/stories/2011/researchers-decipher-protein-structure-of-key-molecule-in-dna-tr
Human Frontier Science Program:http://www.hfsp.org/frontier-science/awardees-articles/architecture-mediator-head-module-revealed
略歴:2007年 横浜市立大学大学院国際総合科学研究科 修了,同年より米国Indiana大学School of Medicine博士研究員.
研究テーマ:転写メディエーター複合体の構造科学.
抱負:超分子複合体の構造および機能を明らかにしていきたい.
高木 雄一郎(Yuichiro Takagi)
略歴:1997年 米国Oklahoma大学にてPh.D.取得,1998年 米国Stanford大学 博士研究員,2003年 同 研究助手を経て,2006年より米国Indiana大学School of MedicineにてAssistant Professor.
研究テーマ:真核生物における転写の制御機構.
抱負:転写メディエーター複合体の全体構造の解明,また,ほかの超分子複合体の構造解明をめざしたい.
研究室URL:http://www.biochemistry.iu.edu/pe/primary-faculty/yuichiro-takagi-ph-d
© 2011 今崎 剛・高木雄一郎 Licensed under CC 表示 2.1 日本
(米国Indiana大学School of Medicine,Department of Biochemistry and Molecular Biology)
email:今崎 剛,高木雄一郎
DOI: 10.7875/first.author.2011.119
Architecture of the Mediator head module.
Tsuyoshi Imasaki, Guillermo Calero, Gang Cai, Kuang-Lei Tsai, Kentaro Yamada, Francesco Cardelli, Hediye Erdjument-Bromage, Paul Tempst, Imre Berger, Guy Lorch Kornberg, Francisco J. Asturias, Roger D. Kornberg, Yuichiro Takagi
Nature, 475, 240-243 (2011)
要 約
転写メディエーター複合体は遺伝子の発現機構において重要な役割をはたし,エンハンサーからの情報をRNAポリメラーゼIIへと伝達するのがその役割である.その構造および機能は酵母からヒトまで保存されており,酵母では21個のサブユニットからなる超分子複合体を形成し,ヘッドモジュール,ミドル/アームモジュール,テイルモジュールと名づけられた3つの独立したモジュールから構成されている.転写機構の解明において転写メディエーター複合体の機能および構造の解析は必須であるが,転写メディエーター複合体は巨大かつ複雑であるため研究は困難をきたしていた.今回,筆者らは,転写メディエーター複合体において主要な機能をつかさどるヘッドモジュールのX線結晶構造解析を行い,分解能4.3Åでの構造の決定に成功した.このヘッドモジュールは7つのサブユニットが複雑にからみあうというまったく新規な構造をとっていた.そのことから3つのドメインからなる柔軟性と安定性とをもちあわせ,RNAポリメラーゼIIおよび基本転写因子との複雑な相互作用を可能にしていることを解明した.
はじめに
転写は遺伝子の発現においてもっとも重要な生命現象のひとつであり,その分子機構の解明は生命科学における重要な課題である.真核生物ではRNAポリメラーゼIIがすべてのmRNAの合成を行っている.このRNAポリメラーゼIIは遺伝子の上流の領域に結合するエンハンサー,あるいは,基本転写因子などさまざまな因子により制御されている.超分子複合体である転写メディエーター複合体はこれらを機能的に結びつけており,転写を理解するうえでもっとも重要な因子のひとつである1).転写メディエーター複合体は酵母からヒトにいたるまで機能的および構造的に保存されている.出芽酵母では21個のサブユニットからなり,ヘッドモジュール,ミドル/アームモジュール,テイルモジュールと名づけられた3つの独立したモジュールから構成されている2).しかし,多数のサブユニットから構成されるという複雑さ,細胞における発現量の少なさ,1 MDaをこえる大きさから構造研究は難航しており,分子機構についての詳細はほとんどわかっていない.
筆者らは,数々の最新技術を導入することにより,転写メディエーター複合体の主要な機能をつかさどるヘッドモジュール3) の構造決定に成功した.驚くべきことに,このヘッドモジュールは7つのサブユニットが複雑にからみあい3つのドメインを形成して安定性と柔軟性とをあわせもつという構造をとっていた.構造解析,生化学的な実験,遺伝学的な解析,そして,過去の研究報告から,転写メディエーター複合体のヘッドモジュールはRNAポリメラーゼIIや基本転写因子と相互作用し,それぞれを適切な位置に配置する調整役として機能する機構を明らかにした.
1.超分子複合体の発現と構造決定
超分子複合体の構造解析はその大きさ,複雑さ,結晶化試料の大量調製のむずかしさ,高品質な結晶調製の困難さ,などさまざまな問題があることから難解である.現在までに構造解析された超分子複合体は,リボソーム,RNAポリメラーゼIIを含め,天然に発現したタンパク質から調製した試料を用いた例がほとんどである.しかし,転写メディエーター複合体の発現量は出芽酵母を100リットル培養しても精製ののち数mgと,この方法で30 mg程度は得られるRNAポリメラーゼIIなどと比べ非常に低く,結晶構造解析には組換えタンパク質による大量発現法を用いることが必須であった.そこで,バキュロウイルス-昆虫細胞を用いた組換えタンパク質発現系を用い,転写メディエーター複合体の主要な機能をつかさどるヘッドモジュールを発現させた.超分子複合体の発現のため開発されたMultiBacシステム4) と,複数の遺伝子を同時にクローニングすることのできるSLIC(sequence and ligation independent iloning)法5) とを組み合わせ,高効率な超分子複合体の発現系を実現した.この発現系を最大限に利用することで,64種類にものぼるコンストラクトについてスクリーニングを行った結果,結晶構造解析に適した結晶を得ることに成功し,それを用いてX線結晶構造解析を行った.不運にもこの結晶は単結晶ではなく双晶(merohedral twin)であったため,それをできるだけ単結晶に修正する条件の検索に多大な時間を費やすことになった.
構造解析は重原子クラスターを用いた初期位相の決定とセレノメチオニン置換タンパク質の単波長異常分散法とを組み合わせ,初期電子密度を分解能4.3Åで得ることに成功した.この低い分解能のため,低分解能でもモデル構築のしやすい領域のモデル構築をまず行い,さらにセレノメチオニンに由来するメチオニン残基の位置,2次構造の予測,サイズの大きい芳香族側鎖の位置を目印にアミノ酸残基の位置決定を行った.その結果,7つのサブユニットからなる超分子複合体,転写メディエーター複合体のヘッドモジュールの結晶構造を分解能4.3Åで決定することに成功した(図1,PDB ID:3RJ1).
2.転写メディエーター複合体のヘッドモジュールの全体構造
ヘッドモジュールの全体構造は,以前,筆者らのグループが報告した電子線顕微鏡による構造解析の結果と同じく,アルファベットのYのようなかたちをしていた6)(図1).7つのサブユニットが複雑にからみあい3つの独立したドメインを形成しており,ドメインどうしは柔軟なリンカー領域でつながれていた.この3つのドメインをNeckドメイン,Fixed Jawドメイン,Movable Jawドメインと名づけた.柔軟なリンカーでつながれていることは,電子線顕微鏡による構造解析においてMovable Jawドメインがいくつかの異なったコンフォメーションをとることを非常によく説明することができた.
3.転写メディエーター複合体のヘッドモジュールにおける複合体の形成機構
複合体の形成機構の謎はNeckドメインのユニークな構造にかくされていた.Neckドメインは5種類のサブユニットからなる10本のαヘリカルバンドルをコアとした非常にユニークな構造をとっていた(図2a).αヘリカルバンドル構造は膜タンパク質などによくみられそれ自体に目新しいことはない.ところが驚くべきことに,Neckドメインの場合,10本のαヘリカルバンドルが5種類のまったく違ったサブユニットから構成されていた.この構造をさらにくわしくみると,NeckドメインにおいてはまずMed11-Med17-Med22からなる5本の長いαヘリカルバンドル構造が安定な相互作用をつくり,そこにMed6-Med8がからみつくようにして複合体のあいだの相互作用を形成していた.また,転写メディエーター複合体のヘッドモジュールは基本転写因子TFIIDのサブユニットのひとつ,TBP(TATA binding protein)やTFIIHとの相互作用が報告されている7,8).これらの相互作用の領域はNeckドメインにあり,このことから,Neckドメインは複合体の形成のみならずタンパク質間相互作用にも重要な役割をもつことが示唆された.
4.転写におけるヘッドモジュールの役割
近年,転写の過程において転写開始から鎖伸長に切り替わる過程が幹細胞の自己複製に重要な役割をはたしていることが明らかになった9).この過程では,RNAポリメラーゼIIのもつC末端ドメインが基本転写因子TFIIHによりリン酸化をうけることが必要であり,このリン酸化は転写メディエーター複合体のRNAポリメラーゼIIからの解離も促進し,転写の過程が転写開始から鎖伸長へと切り替わるのに重要な役割をはたしている10).さらに,転写メディエーター複合体はTFIIHによるRNAポリメラーゼIIのC末端ドメインSer5のリン酸化を促進することが報告されている11).しかし,RNAポリメラーゼIIのC末端ドメイン,転写メディエーター複合体,そして,TFIIHとのあいだの相互作用の機構は謎につつまれていた.この研究で得られた転写メディエーター複合体の構造情報と過去の研究報告とをあわせると,ヘッドモジュールはRNAポリメラーゼIIとTFIIHをつなぐ架け橋となり,それぞれの位置を安定化し転写開始から鎖伸長への切り替えを促進しているのではないかと考えられた(図2b).
おわりに
転写メディエーター複合体のヘッドモジュールの構造決定は想像以上に困難であったが,最先端のタンパク質発現技術とX線構造解析法とを駆使することでこれに成功し,構造生物学の分野にブレークスルーをもたらすことができた.驚くべきことに,転写メディエーター複合体のヘッドモジュールは7つのサブユニットが複雑にからみあった構造を形成することで安定性と柔軟性とをあわせもつという,過去に観測されたことのない新規な構造をとっていた.このような構造はそれぞれのサブユニットをターゲットとした従来の構造解析法により解明することは不可能であり,超分子複合体の全体をターゲットとしたことではじめて明らかになった成果である.この研究は,超分子複合体は複合体として研究しなければその実態はわからないという好例であった.細胞には数多くの超分子複合体が存在し重要なはたらきをしており,これら超分子複合体の構造および機能の研究は今後の重要課題である.また,超分子複合体の構造研究が基礎研究のみならず,超分子複合体をターゲットにしたドラッグデザインなどの応用に重要な役割をはたすことは明らかであろう.
この研究に関係するPress Releases
米国Indiana大学:http://communications.medicine.iu.edu/newsroom/stories/2011/researchers-decipher-protein-structure-of-key-molecule-in-dna-tr
Human Frontier Science Program:http://www.hfsp.org/frontier-science/awardees-articles/architecture-mediator-head-module-revealed
文 献
- Kornberg, R. G.: Mediator and the mechanism of transcriptional activation. Trends Biochem. Sci., 30, 235-239 (2005)[PubMed]
- Cai, G., Imasaki, T., Takagi, Y. et al.: Mediator structural conservation and implications for the regulation mechanism. Structure, 17, 559-567 (2009)[PubMed]
- Takagi, Y., Calero, G., Komori, H. et al.: Head module control of mediator interactions. Mol. Cell, 23, 355-64 (2006)[PubMed]
- Fitzgerald, D. J., Berger, P., Schaffitzel, C. et al.: Protein complex expression by using multigene baculoviral vectors. Nat. Methods, 12, 1021-1032 (2006)[PubMed]
- Li, M. Z. & Elledge, S. J.: Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods, 3, 251-256 (2007)[PubMed]
- Cai, G., Imasaki, T., Yamada, K. et al.: Mediator head module strucutre and functional interactions. Nat. Struct. Mol. Biol., 17, 273-279 (2010)[PubMed]
- Lariviere, L. Gelger, S., Hoeppner, S. et al.: Structure and TBP binding of the Mediator head subcomplex Med8-Med18-Med20. Nat. Struct. Mol. Biol., 13, 895-901 (2006)[PubMed]
- Esnault, C., Chavi-Helm, Y., Brun, S. et al.: Mediator-dependent recruitment of TFIIH modules in preinitiation complex. Mol. Cell, 31, 337-346 (2008)[PubMed]
- Boyer, L. A., Lee, T. I., Cole, M. F. et al.: Core transcriptional regulatory circuitry in human embryonic stem cells. Cell, 122, 947-956 (2005)[PubMed]
- Payne, J. M., Laybourn, P. J. & Dahmus, M. E.: The transition of RNA polymerase II from initiation to elongation is associated with phosphorylation of the carboxyl-terminal domain of subunit IIa. J. Biol. Chem., 264, 19621-19629 (1989)[PubMed]
- Kim, Y. J., Bjorklund, S., Li, Y. et al.: A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II. Cell, 77, 599-608 (1994)[PubMed]
著者プロフィール
略歴:2007年 横浜市立大学大学院国際総合科学研究科 修了,同年より米国Indiana大学School of Medicine博士研究員.
研究テーマ:転写メディエーター複合体の構造科学.
抱負:超分子複合体の構造および機能を明らかにしていきたい.
高木 雄一郎(Yuichiro Takagi)
略歴:1997年 米国Oklahoma大学にてPh.D.取得,1998年 米国Stanford大学 博士研究員,2003年 同 研究助手を経て,2006年より米国Indiana大学School of MedicineにてAssistant Professor.
研究テーマ:真核生物における転写の制御機構.
抱負:転写メディエーター複合体の全体構造の解明,また,ほかの超分子複合体の構造解明をめざしたい.
研究室URL:http://www.biochemistry.iu.edu/pe/primary-faculty/yuichiro-takagi-ph-d
© 2011 今崎 剛・高木雄一郎 Licensed under CC 表示 2.1 日本
