マウス胚の前後方向を決定する遠位臓側内胚葉の起源と役割
高岡勝吉・濱田博司
(大阪大学大学院生命機能研究科 発生遺伝学研究室)
email:高岡勝吉
DOI: 10.7875/first.author.2011.094
Origin and role of distal visceral endoderm, a group of cells that determines anterior-posterior polarity of the mouse embryo.
Katsuyoshi Takaoka, Masamichi Yamamoto, Hiroshi Hamada
Nature Cell Biology, 13, 743-752 (2011)
マウス胚の前後軸に関する研究は,前後方向を決定する遠位臓側内胚葉の標識遺伝子であるHex遺伝子を中心に解析が進められてきた.しかし今回,筆者らは,別の遠位臓側内胚葉の標識遺伝子であるLefty1遺伝子に着目し,その発現細胞の系譜と細胞分化に注目して解析を行った.その結果,受精後5.5日胚において形成されると考えられてきた遠位臓側内胚葉は受精後4.0日胚においてすでに運命決定していること,従来は同じであると考えられてきた遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることを明らかにした.また,遠位臓側内胚葉の遺伝的な除去実験を行うことで,遠位臓側内胚葉は前後方向を決定する役割をもち,頭部を誘導する前方臓側内胚葉を前側へとガイドしていることが明らかになった.以上の成果より,前後軸の形成における従来のモデルをくつがえし,新たなモデルを提唱するにいたった.
ショウジョウバエを含む多くの生物はbicoid遺伝子に代表される母性効果遺伝子のmRNAが非対称に分布することにより,卵子の段階ですでに分子レベルの非対称情報を獲得している.そして,この非対称情報がのちの体軸形成のための情報を提供する1).それに対して,ヒトやマウスといった哺乳類は分裂期の胚が高い操作性や適応能を備えていることから,卵子や受精卵の時期には分子レベルの非対称情報は獲得しておらず,そののちの発生段階における“非対称性の起源”をへて体軸形成のための情報を獲得するのであろうと考えられている2,3).では,“非対称性の起源”はいつ,どのようにして起こるのだろうか? 筆者らは,この命題を解明すべくマウス胚においてもっとも早期に形成される前後軸の初期形成における分子機構に着目してきた.
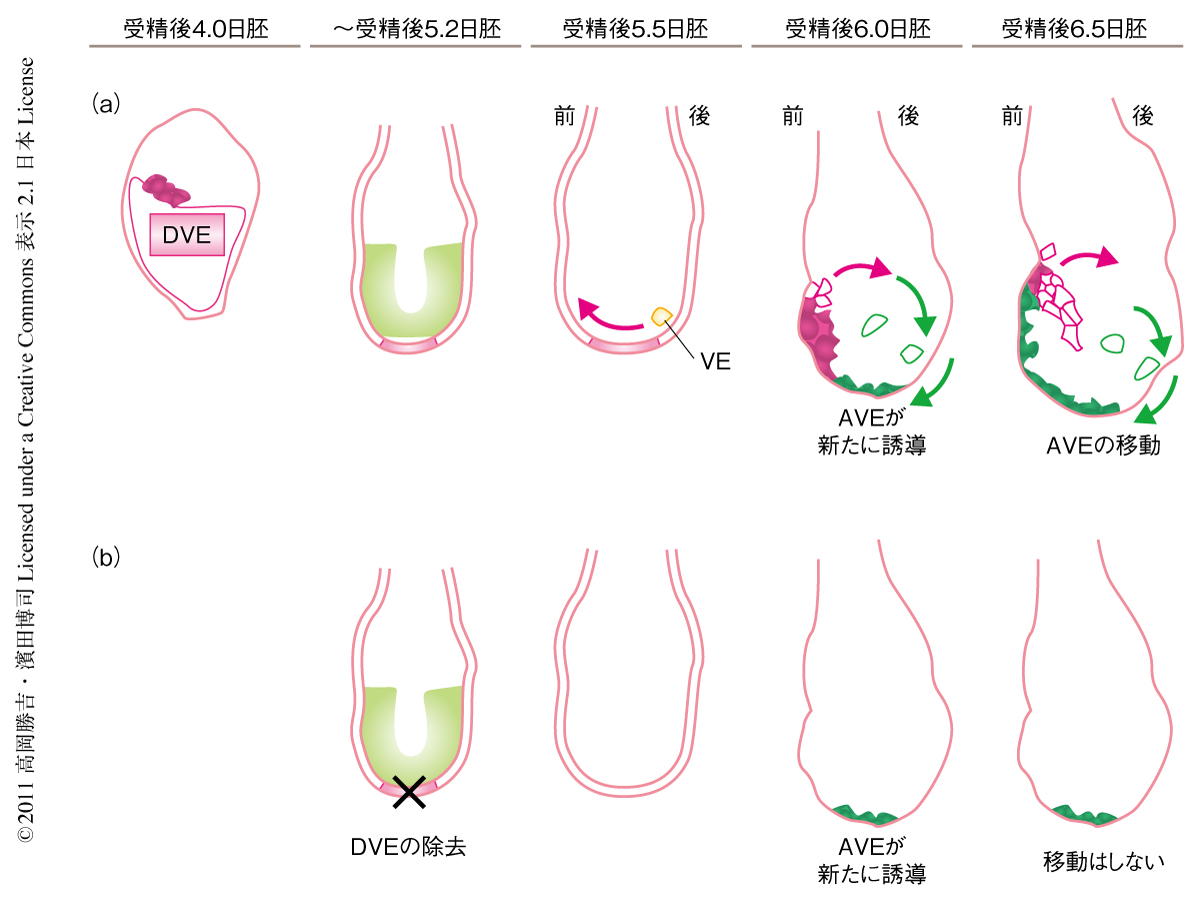
前後軸に関する研究は,遠位臓側内胚葉(distal visceral endoderm,DVE)の標識遺伝子であるHex遺伝子を中心に解析が進められてきた2,4,5).これまでの研究によれば,まず,受精後5.5日胚の遠位端にさまざまな遺伝子の発現する遠位臓側内胚葉が形成される.この遠位臓側内胚葉は受精後5.7日胚において将来の前側になる部分へと移動し,前方臓側内胚葉(anterior visceral endoderm:AVE)と名称を変える.そして,前方臓側内胚葉が将来の体になる胚体の部分に後方化抑制シグナルをあたえることにより頭部ができ,形態的な前後軸が形成されるとされてきた2-5)(図1a).

筆者らはこれまでに,遠位臓側内胚葉の標識遺伝子のひとつであるLefty1遺伝子は受精後5.5日胚よりまえの受精後3.5日胚の一部の細胞で発現を開始していることを見い出していた6,7).この発見により,遠位臓側内胚葉はもっと早期に運命決定している可能性が示唆された.そこで,受精後5.5日胚よりまえの胚のLefty1遺伝子陽性細胞が遠位臓側内胚葉に運命決定しているかどうかを調べるため,Lefty1遺伝子陽性細胞をタモキシフェン誘導性CreERT2で発現するβガラクトシダーゼにより標識したトランスジェニック胚を用いて,Lefty1遺伝子陽性細胞の細胞系譜を詳細に解析した.その結果,受精後3.5日胚のLefty1遺伝子陽性細胞は受精後5.5日胚のエピブラストに寄与し,受精後4.2日胚のLefty1遺伝子陽性細胞は遠位臓側内胚葉に寄与していることが明らかになった.
また,Lefty1遺伝子陽性細胞を膜移行型の蛍光タンパク質Venusにより標識したトランスジェニック胚を受精後3.7日胚から受精後4.2日胚まで経時観察した結果,将来,遠位臓側内胚葉となる部分におけるLefty1遺伝子の発現は,受精後4.0日胚の付近でエピブラストに分化するLefty1遺伝子陽性細胞とは異なる細胞に,遅れて誘導されていることが明らかになった.
以上の結果より,遠位臓側内胚葉は受精後4.0日胚においてすでに運命決定していることが明らかになった(図1b).
従来は,遠位臓側内胚葉と前方臓側内胚葉とは同じ細胞系譜であると考えられてきた2,3)(図1a).しかし,Lefty1遺伝子陽性細胞をβガラクトシダーゼにより標識したトランスジェニック胚を用いて受精後5.5日胚よりのちのLefty1遺伝子陽性細胞の細胞系譜を解析した結果,遠位臓側内胚葉は受精後6.5日胚では前方臓側内胚葉とは異なる細胞に寄与しており,遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることが明らかになった(図1b).また,別の遠位臓側内胚葉の標識遺伝子であるCerl遺伝子の発現細胞の系譜を解析した結果も,Lefty1遺伝子の結果と同様に,遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることを示していた.
受精後5.5日から受精後6.5日胚における遠位臓側内胚葉と前方臓側内胚葉のそれぞれの形成過程と位置関係を明らかにするため,すべての臓側内胚葉の細胞膜を赤色蛍光タンパク質Tomatoにより標識したトランスジェニック胚と,遠位臓側内胚葉および前方臓側内胚葉を緑色蛍光タンパク質Venusにより標識したトランスジェニック胚を用いて,経時観察を行った.2種類の蛍光を経時的に観察することで,すべての臓側内胚葉と遠位臓側内胚葉および前方臓側内胚葉の細胞の動きを1細胞レベルで追跡することが可能になった.受精後5.5日胚から受精後6.5日胚まで観察した結果,遠位臓側内胚葉の移動にともない,胚の遠位側でそれまでLefty1遺伝子の発現のない臓側内胚葉の細胞において新たにLefty1遺伝子の発現が誘導されており,この新たに誘導されたLefty1遺伝子発現細胞は前側へと移動し,受精後6.5日胚において前方臓側内胚葉を形成していることがわかった.
また,Venusはタンパク質自体が安定であるため,受精後6.5日胚においても遠位臓側内胚葉の子孫細胞と前方臓側内胚葉の両方で蛍光を検出することができた.そこで,遠位臓側内胚葉および前方臓側内胚葉をVenusにより標識したトランスジェニック胚を用いて受精後6.5日胚において蛍光領域と内在性のLefty1遺伝子の発現領域とを比較した.その結果,受精後6.5日胚の遠位臓側内胚の子孫細胞では内在性のLefty1遺伝子は発現していないことがわかった.別の遠位臓側内胚葉の標識遺伝子であるCerl遺伝子の発現細胞の細胞膜をVenusにより標識したトランスジェニック胚を用いて蛍光領域と内在性のCerl遺伝子の発現領域とを比較した結果もLefty1遺伝子の結果と同様であり,受精後6.5日胚の遠位臓側内胚葉の子孫細胞では内在性のCerl遺伝子は発現していないことがわかった.
以上の結果より,胚の遠位側では遠位臓側内胚葉が移動するにともない前方臓側内胚葉が新たに誘導されること,胚の近位側では遠位臓側内胚葉の移動にともない遠位臓側内胚葉の標識遺伝子の発現が消失していることがわかった.
遠位臓側内胚葉は前方臓側内胚葉とは異なる細胞系譜であることが明らかになったことにより,遠位臓側内胚葉の役割を再定義する必要が生じた.そこで,遠位臓側内胚葉を遺伝的に除去する実験を行った.その結果,受精後5.5日胚において遠位臓側内胚葉の除去ののちしばらくすると遠位端で前方臓側内胚葉が誘導されるが,これは前側へは移動せず遠位端にとどまったままであった.これまでの研究により前方臓側内胚葉は頭部を誘導する機能をもっているとされている8).この結果より,遠位臓側内胚葉は頭部を誘導する前方臓側内胚葉を前側にガイドする役割を担っているものと考えられた.
すべての臓側内胚葉の細胞膜をTomatoにより標識したトランスジェニック胚と遠位臓側内胚葉および前方臓側内胚葉をVenusにより標識したトランスジェニック胚を用いて,受精後5.2日胚から受精後6.5日胚において臓側内胚葉の全体の細胞の動きを経時観察した.すると,受精後5.2日から受精後5.5日胚においては臓側内胚葉はほとんど動いていないが,遠位臓側内胚葉が移動を開始する受精後5.5日胚よりのちの臓側内胚葉は,胚の左側からみたとき時計回りの運動をしていた.以上の観察より,受精後5.5日胚において臓側内胚葉の全体の動きが開始することがわかった.
では,臓側内胚葉の全体の動きの駆動力は何なのであろうか? 臓側内胚葉の全体が動くタイミングは,受精後5.5日胚よりのちの遠位臓側内胚葉が移動するタイミングであることから,遠位臓側内胚葉の移動が臓側内胚葉の全体の動きをひき起こしているという仮説をたてた.この仮説を証明するため,遠位臓側内胚葉を除去した胚における臓側内胚葉の全体の動きを観察したところ,動きはみられなかった.つまり,受精後5.5日胚において遠位臓側内胚葉の移動が開始することでこれが駆動力となり,臓側内胚葉の全体の時計回りの動き(胚の左側からみたとき)が生み出される.この動きによりLefty1遺伝子の発現のない臓側内胚葉が胚の遠位端に位置すると,新たにLefty1遺伝子が誘導され前方臓側内胚葉となる(図2).
これまでの常識にとらわれず,細胞の由来と系譜という従来とは異なる観点で解析した結果,新たな前後軸形成モデルを提唱するにいたった.以上の成果より,これまで前後軸が正常に形成されないと報告されている変異マウスの解析を再検証する必要がでてきた.たとえば,従来は遠位臓側内胚葉と前方臓側内胚葉とが区別されずに解析されていたが,この2つは異なる細胞系譜であるためそれぞれ区別して検証しなければならない.実際に,前方臓側内胚葉の形成は起こるが遠位臓側内胚葉の形成は起らないというマウス変異胚が報告されている9).
今後は,“非対称性の起源”にせまるべく,受精後4.0日胚より以前にさかのぼって解析をしたい.とくに,遠位臓側内胚葉のような特別な細胞がどのようにして均一な細胞のなかから選ばれているのかという疑問を明らかにしたい.ES細胞やiPS細胞から複雑な構造をもつ臓器をつくるためには,目的の臓器の細胞を誘導するだけでなく,細胞集団へ正確な位置情報をあたえ高度な構造をもたせることも重要である.この研究から得られた知見は,体の非対称性の起源という発生生物学の命題にせまるとともに,将来の再生医療の基盤となるものと期待している.
略歴:2009年 大阪大学大学院生命機能研究科一貫制博士課程 修了,同年より同 助教.
研究テーマ:マウス胚における体軸のはじまり.
抱負:“夢とロマン”のある研究をする.
濱田 博司(Hiroshi Hamada)
大阪大学大学院生命機能研究科 教授.
研究室URL:http://www.fbs.osaka-u.ac.jp/labs/hamada/
© 2011 高岡勝吉・濱田博司 Licensed under CC 表示 2.1 日本
(大阪大学大学院生命機能研究科 発生遺伝学研究室)
email:高岡勝吉
DOI: 10.7875/first.author.2011.094
Origin and role of distal visceral endoderm, a group of cells that determines anterior-posterior polarity of the mouse embryo.
Katsuyoshi Takaoka, Masamichi Yamamoto, Hiroshi Hamada
Nature Cell Biology, 13, 743-752 (2011)
要 約
マウス胚の前後軸に関する研究は,前後方向を決定する遠位臓側内胚葉の標識遺伝子であるHex遺伝子を中心に解析が進められてきた.しかし今回,筆者らは,別の遠位臓側内胚葉の標識遺伝子であるLefty1遺伝子に着目し,その発現細胞の系譜と細胞分化に注目して解析を行った.その結果,受精後5.5日胚において形成されると考えられてきた遠位臓側内胚葉は受精後4.0日胚においてすでに運命決定していること,従来は同じであると考えられてきた遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることを明らかにした.また,遠位臓側内胚葉の遺伝的な除去実験を行うことで,遠位臓側内胚葉は前後方向を決定する役割をもち,頭部を誘導する前方臓側内胚葉を前側へとガイドしていることが明らかになった.以上の成果より,前後軸の形成における従来のモデルをくつがえし,新たなモデルを提唱するにいたった.
はじめに
ショウジョウバエを含む多くの生物はbicoid遺伝子に代表される母性効果遺伝子のmRNAが非対称に分布することにより,卵子の段階ですでに分子レベルの非対称情報を獲得している.そして,この非対称情報がのちの体軸形成のための情報を提供する1).それに対して,ヒトやマウスといった哺乳類は分裂期の胚が高い操作性や適応能を備えていることから,卵子や受精卵の時期には分子レベルの非対称情報は獲得しておらず,そののちの発生段階における“非対称性の起源”をへて体軸形成のための情報を獲得するのであろうと考えられている2,3).では,“非対称性の起源”はいつ,どのようにして起こるのだろうか? 筆者らは,この命題を解明すべくマウス胚においてもっとも早期に形成される前後軸の初期形成における分子機構に着目してきた.
前後軸に関する研究は,遠位臓側内胚葉(distal visceral endoderm,DVE)の標識遺伝子であるHex遺伝子を中心に解析が進められてきた2,4,5).これまでの研究によれば,まず,受精後5.5日胚の遠位端にさまざまな遺伝子の発現する遠位臓側内胚葉が形成される.この遠位臓側内胚葉は受精後5.7日胚において将来の前側になる部分へと移動し,前方臓側内胚葉(anterior visceral endoderm:AVE)と名称を変える.そして,前方臓側内胚葉が将来の体になる胚体の部分に後方化抑制シグナルをあたえることにより頭部ができ,形態的な前後軸が形成されるとされてきた2-5)(図1a).
1.遠位臓側内胚葉は受精後4.0日胚においてすでに運命決定されている
筆者らはこれまでに,遠位臓側内胚葉の標識遺伝子のひとつであるLefty1遺伝子は受精後5.5日胚よりまえの受精後3.5日胚の一部の細胞で発現を開始していることを見い出していた6,7).この発見により,遠位臓側内胚葉はもっと早期に運命決定している可能性が示唆された.そこで,受精後5.5日胚よりまえの胚のLefty1遺伝子陽性細胞が遠位臓側内胚葉に運命決定しているかどうかを調べるため,Lefty1遺伝子陽性細胞をタモキシフェン誘導性CreERT2で発現するβガラクトシダーゼにより標識したトランスジェニック胚を用いて,Lefty1遺伝子陽性細胞の細胞系譜を詳細に解析した.その結果,受精後3.5日胚のLefty1遺伝子陽性細胞は受精後5.5日胚のエピブラストに寄与し,受精後4.2日胚のLefty1遺伝子陽性細胞は遠位臓側内胚葉に寄与していることが明らかになった.
また,Lefty1遺伝子陽性細胞を膜移行型の蛍光タンパク質Venusにより標識したトランスジェニック胚を受精後3.7日胚から受精後4.2日胚まで経時観察した結果,将来,遠位臓側内胚葉となる部分におけるLefty1遺伝子の発現は,受精後4.0日胚の付近でエピブラストに分化するLefty1遺伝子陽性細胞とは異なる細胞に,遅れて誘導されていることが明らかになった.
以上の結果より,遠位臓側内胚葉は受精後4.0日胚においてすでに運命決定していることが明らかになった(図1b).
2.遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜である
従来は,遠位臓側内胚葉と前方臓側内胚葉とは同じ細胞系譜であると考えられてきた2,3)(図1a).しかし,Lefty1遺伝子陽性細胞をβガラクトシダーゼにより標識したトランスジェニック胚を用いて受精後5.5日胚よりのちのLefty1遺伝子陽性細胞の細胞系譜を解析した結果,遠位臓側内胚葉は受精後6.5日胚では前方臓側内胚葉とは異なる細胞に寄与しており,遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることが明らかになった(図1b).また,別の遠位臓側内胚葉の標識遺伝子であるCerl遺伝子の発現細胞の系譜を解析した結果も,Lefty1遺伝子の結果と同様に,遠位臓側内胚葉と前方臓側内胚葉とは異なる細胞系譜であることを示していた.
3.前方臓側内胚葉は受精後5.5日胚よりのちに胚の遠位側で新たに誘導される
受精後5.5日から受精後6.5日胚における遠位臓側内胚葉と前方臓側内胚葉のそれぞれの形成過程と位置関係を明らかにするため,すべての臓側内胚葉の細胞膜を赤色蛍光タンパク質Tomatoにより標識したトランスジェニック胚と,遠位臓側内胚葉および前方臓側内胚葉を緑色蛍光タンパク質Venusにより標識したトランスジェニック胚を用いて,経時観察を行った.2種類の蛍光を経時的に観察することで,すべての臓側内胚葉と遠位臓側内胚葉および前方臓側内胚葉の細胞の動きを1細胞レベルで追跡することが可能になった.受精後5.5日胚から受精後6.5日胚まで観察した結果,遠位臓側内胚葉の移動にともない,胚の遠位側でそれまでLefty1遺伝子の発現のない臓側内胚葉の細胞において新たにLefty1遺伝子の発現が誘導されており,この新たに誘導されたLefty1遺伝子発現細胞は前側へと移動し,受精後6.5日胚において前方臓側内胚葉を形成していることがわかった.
また,Venusはタンパク質自体が安定であるため,受精後6.5日胚においても遠位臓側内胚葉の子孫細胞と前方臓側内胚葉の両方で蛍光を検出することができた.そこで,遠位臓側内胚葉および前方臓側内胚葉をVenusにより標識したトランスジェニック胚を用いて受精後6.5日胚において蛍光領域と内在性のLefty1遺伝子の発現領域とを比較した.その結果,受精後6.5日胚の遠位臓側内胚の子孫細胞では内在性のLefty1遺伝子は発現していないことがわかった.別の遠位臓側内胚葉の標識遺伝子であるCerl遺伝子の発現細胞の細胞膜をVenusにより標識したトランスジェニック胚を用いて蛍光領域と内在性のCerl遺伝子の発現領域とを比較した結果もLefty1遺伝子の結果と同様であり,受精後6.5日胚の遠位臓側内胚葉の子孫細胞では内在性のCerl遺伝子は発現していないことがわかった.
以上の結果より,胚の遠位側では遠位臓側内胚葉が移動するにともない前方臓側内胚葉が新たに誘導されること,胚の近位側では遠位臓側内胚葉の移動にともない遠位臓側内胚葉の標識遺伝子の発現が消失していることがわかった.
4.遠位臓側内胚葉は頭部を誘導する前方臓側内胚葉を前側にガイドする
遠位臓側内胚葉は前方臓側内胚葉とは異なる細胞系譜であることが明らかになったことにより,遠位臓側内胚葉の役割を再定義する必要が生じた.そこで,遠位臓側内胚葉を遺伝的に除去する実験を行った.その結果,受精後5.5日胚において遠位臓側内胚葉の除去ののちしばらくすると遠位端で前方臓側内胚葉が誘導されるが,これは前側へは移動せず遠位端にとどまったままであった.これまでの研究により前方臓側内胚葉は頭部を誘導する機能をもっているとされている8).この結果より,遠位臓側内胚葉は頭部を誘導する前方臓側内胚葉を前側にガイドする役割を担っているものと考えられた.
5.受精後5.5日胚において臓側内胚葉の全体の動きが開始する
すべての臓側内胚葉の細胞膜をTomatoにより標識したトランスジェニック胚と遠位臓側内胚葉および前方臓側内胚葉をVenusにより標識したトランスジェニック胚を用いて,受精後5.2日胚から受精後6.5日胚において臓側内胚葉の全体の細胞の動きを経時観察した.すると,受精後5.2日から受精後5.5日胚においては臓側内胚葉はほとんど動いていないが,遠位臓側内胚葉が移動を開始する受精後5.5日胚よりのちの臓側内胚葉は,胚の左側からみたとき時計回りの運動をしていた.以上の観察より,受精後5.5日胚において臓側内胚葉の全体の動きが開始することがわかった.
6.遠位臓側内胚葉は臓側内胚葉の全体の動きをひき起こすことで前方臓側内胚葉を前側へとガイドする
では,臓側内胚葉の全体の動きの駆動力は何なのであろうか? 臓側内胚葉の全体が動くタイミングは,受精後5.5日胚よりのちの遠位臓側内胚葉が移動するタイミングであることから,遠位臓側内胚葉の移動が臓側内胚葉の全体の動きをひき起こしているという仮説をたてた.この仮説を証明するため,遠位臓側内胚葉を除去した胚における臓側内胚葉の全体の動きを観察したところ,動きはみられなかった.つまり,受精後5.5日胚において遠位臓側内胚葉の移動が開始することでこれが駆動力となり,臓側内胚葉の全体の時計回りの動き(胚の左側からみたとき)が生み出される.この動きによりLefty1遺伝子の発現のない臓側内胚葉が胚の遠位端に位置すると,新たにLefty1遺伝子が誘導され前方臓側内胚葉となる(図2).
おわりに
これまでの常識にとらわれず,細胞の由来と系譜という従来とは異なる観点で解析した結果,新たな前後軸形成モデルを提唱するにいたった.以上の成果より,これまで前後軸が正常に形成されないと報告されている変異マウスの解析を再検証する必要がでてきた.たとえば,従来は遠位臓側内胚葉と前方臓側内胚葉とが区別されずに解析されていたが,この2つは異なる細胞系譜であるためそれぞれ区別して検証しなければならない.実際に,前方臓側内胚葉の形成は起こるが遠位臓側内胚葉の形成は起らないというマウス変異胚が報告されている9).
今後は,“非対称性の起源”にせまるべく,受精後4.0日胚より以前にさかのぼって解析をしたい.とくに,遠位臓側内胚葉のような特別な細胞がどのようにして均一な細胞のなかから選ばれているのかという疑問を明らかにしたい.ES細胞やiPS細胞から複雑な構造をもつ臓器をつくるためには,目的の臓器の細胞を誘導するだけでなく,細胞集団へ正確な位置情報をあたえ高度な構造をもたせることも重要である.この研究から得られた知見は,体の非対称性の起源という発生生物学の命題にせまるとともに,将来の再生医療の基盤となるものと期待している.
文 献
- Huynh, J. R. & St. Johnston, D.: The origin of asymmetry: early polarisation of the Drosophila germline cyst and oocyte. Curr. Biol., 14, 438-449 (2004)[PubMed]
- Beddington, R. S. & Robertson, E. J.: Axis development and early asymmetry in mammals. Cell, 96, 195-209 (1999)[PubMed]
- Rossant, J. & Tam, P. P.: Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. Development, 136, 701-713 (2009)[PubMed]
- Thomas, P. Q., Brown, A. & Beddington, R. S.: Hex: a homeobox gene revealing peri-implantation asymmetry in the mouse embryo and an early transient marker of endothelial cell precursors. Development, 125, 85-94 (1998)[PubMed]
- Srinivas, S., Rodriguez, T., Clements, M. et al.: Active cell migration drives the unilateral movements of the anterior visceral endoderm. Development, 131, 1157-1164 (2004)[PubMed]
- Takaoka, K., Yamamoto, M., Shiratori, H. et al.: The mouse embryo autonomously acquires anterior-posterior polarity at implantation. Dev. Cell, 10, 451-459 (2006)[PubMed]
- Yamamoto, M., Saijoh, Y., Perea-Gomez, A. et al.: Nodal antagonists regulate formation of the anteroposterior axis of the mouse embryo. Nature, 428, 387-392 (2004)[PubMed]
- Kimura, C., Yoshinaga, K., Tian, E. et al.: Visceral endoderm mediates forebrain development by suppressing posteriorizing signals. Dev. Biol., 225, 304-321 (2000)[PubMed]
- Chu, J. & Shen, M. M.: Functional redundancy of EGF-CFC genes in epiblast and extraembryonic patterning during early mouse embryogenesis. Dev. Biol., 342, 63-73 (2010)[PubMed]
著者プロフィール
略歴:2009年 大阪大学大学院生命機能研究科一貫制博士課程 修了,同年より同 助教.
研究テーマ:マウス胚における体軸のはじまり.
抱負:“夢とロマン”のある研究をする.
濱田 博司(Hiroshi Hamada)
大阪大学大学院生命機能研究科 教授.
研究室URL:http://www.fbs.osaka-u.ac.jp/labs/hamada/
© 2011 高岡勝吉・濱田博司 Licensed under CC 表示 2.1 日本
