逐次的なシナプスの興奮と抑制により随意行動の準備活動が形成される
加賀谷勝史・高畑雅一
(北海道大学大学院理学研究院 生物科学部門)
email:加賀谷勝史
DOI: 10.7875/first.author.2011.081
Sequential synaptic excitation and inhibition shape readiness discharge for voluntary behavior.
Katsushi Kagaya, Masakazu Takahata
Science, 332, 365-368 (2011)
動物はどのように随意行動を開始するのか? 神経科学において鍵となる現象が脳の特定領域で起こる準備神経活動である.しかし,その活動を媒介する特定のニューロンおよびシナプス機構については不明であった.筆者らがアメリカザリガニの脳において発見した準備発射活動は,逐次的なシナプスの興奮と抑制により形成されることが判明した.準備発射ニューロンは軸索の側枝を脳に伸ばし,回帰性の神経回路網を形成する局所介在ニューロンを賦活すると考えられた.したがって,筆者らは,下位神経節からのフィードバック信号なしに逐次的なシナプスの賦活により準備発射が形成されているという回帰性の神経回路網を提案した.この神経回路網は自己生成的な随意行動の開始の信号処理に適している.
随意行動の開始に先行して脳の特定領域で特徴的な神経活動の変化が起こる.活動電位として記録される準備発射は霊長類や齧歯類において多くの報告がなされてきた1).しかし,準備発射を形成する脳内ニューロンの樹状突起における膜電位のダイナミクスについては不明であった.アメリカザリガニ(Procambarus clarkii)の脳における準備発射は随意行動の開始のための準備活動の生成における細胞の機構を理解する手がかりをあたえると考えられる.ザリガニでは準備発射は囲食道縦連合において記録される2)(図1a).囲食道縦連合には脳内ニューロンの軸索が走行しており,ザリガニの脳の活動はこの囲食道縦連合を介して下行性ニューロンにより胸部へと伝えられる(図1b).胸部には歩行パターンを生成する中枢パターン発生器が存在する3).準備発射は行動開始の数秒まえに現われ歩行が開始すると消える2).この一過的なスパイク活動は準備発射に共通する特徴である.しかし,どのようなシナプス機構およびニューロンがこの活動の形成にかかわるのかは不明であった.
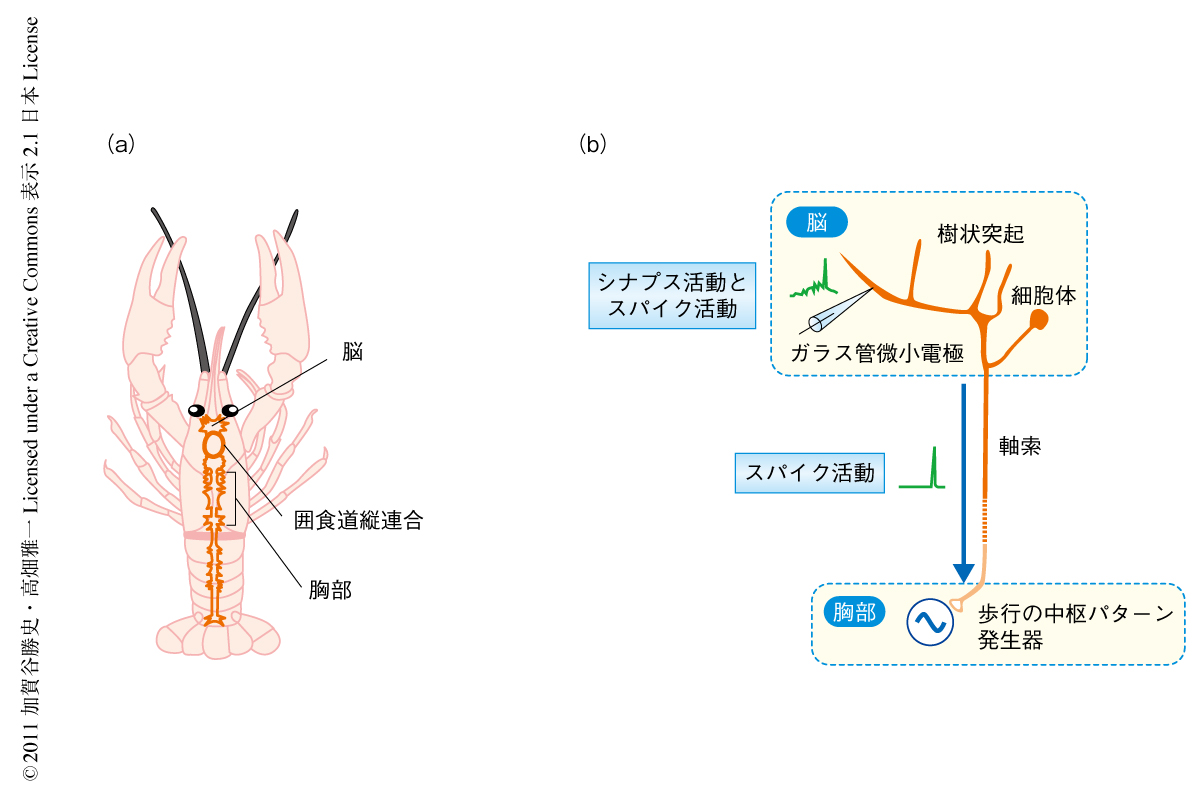
随意歩行の開始の神経生理学的な機構を明らかにする目的で,球形のトレッドミルを歩行するザリガニの脳の単一細胞からの細胞内記録を行った.外部からの感覚刺激はあたえられず,したがって,ザリガニは自ら歩行を開始,維持,停止することができる.実験の結果,合計で69個のニューロンからの細胞内記録および細胞内染色に成功した.同定されたニューロンの機能的な分類は歩行の開始と停止のタイミングにもとづいて行った.ザリガニの歩脚は8本ある.歩脚の筋活動はほぼ同時に(65.89±6.40ミリ秒の時間幅のなかで)活動の上がることが判明している4).そのため,1本の歩脚の長腕節屈筋から記録をとり筋活動の変化から歩行の開始と停止のタイミングを決定した.シナプス活動およびスパイク活動が歩行の開始に先行すること500ミリ秒以上である場合を“先行する”と定義した.形態的には下行性ニューロンと局所介在ニューロンに分類した.下行性ニューロンは囲食道縦連合をとおり胸部へと軸索を伸ばす.局所介在ニューロンは突起の投射を脳に限局している.昆虫では歩行制御の脳内部位として中心複合体が関連すると考えられている5,6).ザリガニの脳における相同部位7) のなかで随意歩行の制御にかかわる部位がどこかを明らかにする目的で脳内ニューロンの突起投射のパターンを調査した.細胞内染色は細胞内記録を行ったあとにガラス管微小電極に充填した蛍光色素Lucifer yellowにより行った.
行動を遂行しているザリガニの準備発射ニューロンからシナプス活動を記録することに成功した.ザリガニは多様な歩行パターンを示したが,準備発射ニューロンの膜電位は歩行パターンとは関係なく,歩行の開始前には脱分極性の変化,開始後には過分極性の変化を示した.さらに,電流注入実験により,この逐次的な膜電位の変化は直接的な興奮性シナプス入力および抑制性シナプス入力によることが判明した.また,膜コンダクタンス変化の解析から,この抑制性の入力は行動開始の直後と数秒後とで樹状突起の異なる入力部位において受け取られる可能性を示した.また,脱分極のときにはシナプスノイズの増加も認められた.これらの結果は,興奮性シナプス入力および2つ以上の抑制性シナプス入力を前シナプスニューロンから受け取ることで準備発射が生成されており,個々のニューロンにおける内発的な活動電位の発生という機構によるものではないことを示していた.
準備発射ニューロンは細胞体を前大脳の腹側前方部にもち樹状突起を前方および後方の内側ニューロピルに投射することが判明した.このニューロンは下行性ニューロンであり軸索は囲食道縦連合のWiersma領域70に伸びていた8).この領域には電気刺激をくわえると歩行をひき起こすユニットの含まれていることが過去の研究でわかっている2,9).また,この領域には準備活動ユニットが含まれ,軸索束の切断断端から蛍光色素を注入して脳における細胞を染色したところ細胞体の位置や樹状突起の形態などの形態的な特徴の一致するニューロンの存在することが判明した2).さらに,後大脳の食道ニューロピルに軸索の側枝の投射がみられた.これは,胸部だけでなく脳にも準備発射をうけるニューロンの存在することを示し,下位神経節を介さずとも脳において歩行の開始の情報が得られることを示していた.
筆者らは,以前の細胞外記録法を用いた報告で,準備発射活動にくわえて歩行の維持および停止にかかわる下行性の活動の存在することを示した.今回,細胞内記録および細胞内染色を用いてそれぞれを代表する下行性ニューロンの同定にも成功した.準備活動ニューロンと共通した特徴として,歩行の維持および停止にかかわるニューロンも後大脳の食道ニューロピルに軸索の側枝をもっていることが判明した.このことは,脳においては下位神経節からのフィードバックなしに行動の開始,維持,停止という情報を得ている可能性を示していた.
スパイク活動から分類した歩行の開始,維持,停止の3つのタイプにくわえて,脳のどの領域でシナプスの賦活が起こっているのかを調べるため,シナプス活動の変化が歩行の開始の前後で観察された45個の下行性ニューロンを同定した.先行したシナプス活動の変化を示すものを先行型ニューロン,ほぼ同時に変化を示すものを随伴型ニューロン,遅れてあるいは変化を示さないものを遅延型ニューロンとして分類し,それぞれの脳における樹状突起の投射部位を調べた.その結果,先行型ニューロンと随伴型ニューロンは樹状突起を後大脳あるいは中大脳に比べ前大脳に多くもつ傾向を示した.一方で,遅延型ニューロンは中大脳と後大脳に樹状突起を多くもつ傾向があった.したがって,前大脳の内側部に限局された投射とシナプス活動から随意歩行の開始にかかわるシナプスの賦活は前大脳の内側部で起こっているものと考えられた.
今回,発見された準備発射ニューロンは興奮性シナプス入力のあとに抑制性シナプス入力をうけていることが判明した.ひとつの可能性として,この抑制性シナプス入力が継続ニューロンによりひき起こされていることが考えられた.準備発射ニューロンおよび継続ニューロンは脳に軸索の側枝をもっており,それぞれの活動を脳において出力するものと考えられた.準備発射ニューロンの樹状突起と継続ニューロンの軸索の側枝の出力領域は重なってはいなかった.多シナプス接続を介して信号が伝えられている,つまり,脳において局所介在ニューロンにより並列的かつ逐次的に下行性回路を賦活しているものと考えられた(図2).
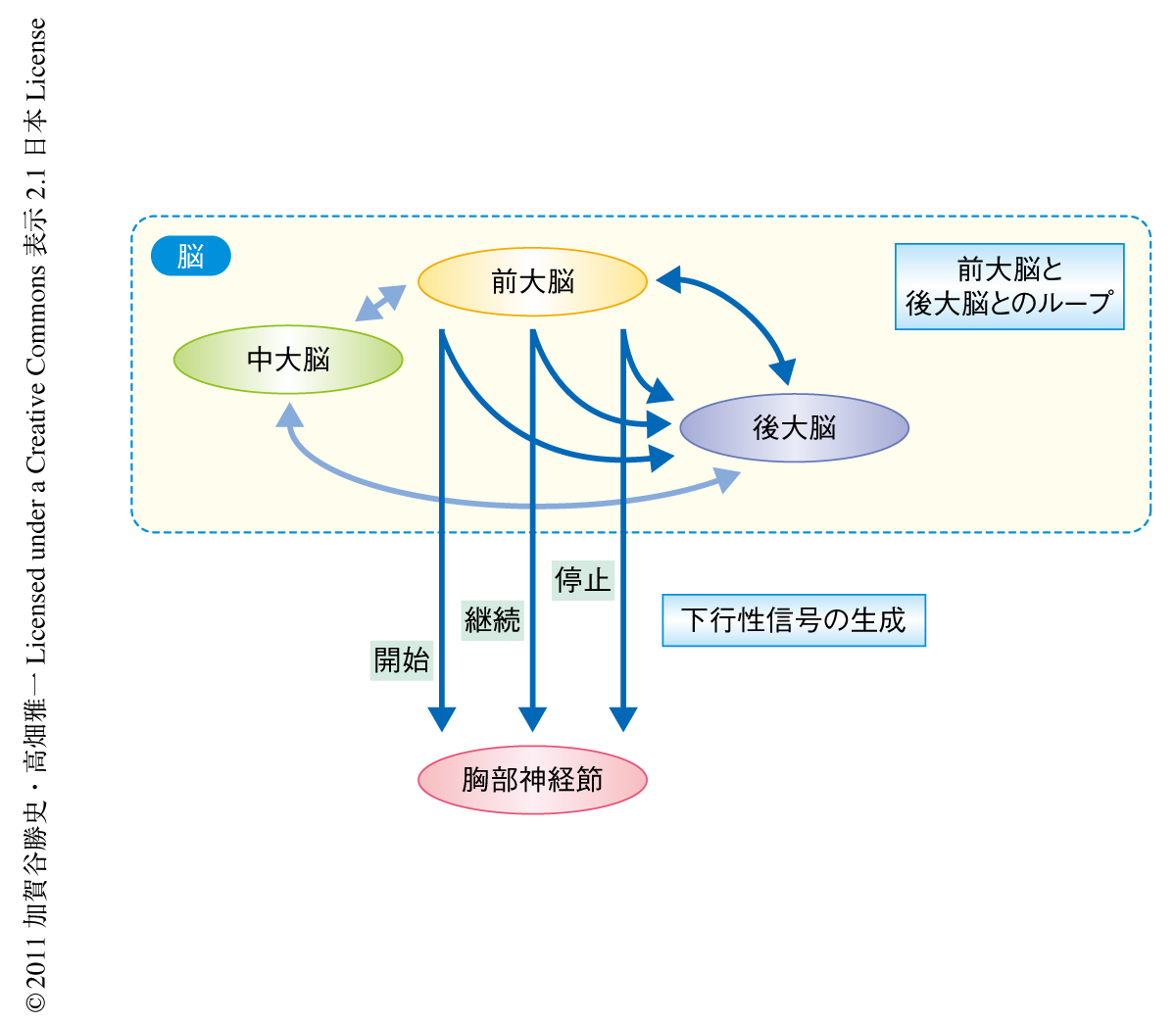
さらに,スパイク発生型およびスパイク非発生型の局所介在ニューロンからの細胞内記録および細胞内染色に成功し,後大脳の食道ニューロピルと前大脳の内側部に突起をもつニューロンを見い出した.これらのシナプスの接続関係については不明であるが,作業仮説として,これら局所介在ニューロンと下行性ニューロンとが前大脳と後大脳においてループを構成するという回帰性の神経回路網が存在すると考えた(図2).
いずれにしても,筆者らのデータは,ザリガニにおいて自己生成的な随意歩行に関連した脳内ニューロンの生理学的および形態学的な特徴を明らかにし,それらが個々の内発的な機構によってではなく,前シナプスニューロンにより賦活されていることを示した.
従来,動物の行動の発現機構,とりわけ無脊椎動物における行動の発現機構は“感覚刺激に対する応答としての行動”という視点により実験的に解析されて多大な知見が得られてきた.そこでは“感覚入力が司令ニューロンに収斂し運動出力へといたる”という一方向性の並列階層的な機構が想定され研究されてきた.一方で,多くの動物は明確に定義された外部からの感覚刺激がなくても自発的に行動を発現し制御している.しかし,自発性の行動発現の単一細胞レベルでの神経回路の動作機構となると無脊椎動物ではほとんど報告がなく,また,そのシナプス機構となると脊椎動物でもまったく未解明である.
随意行動の発現機構を研究する場合,この研究のように運動出力の側からさかのぼってその機構を明らかにしていくのが近道であるが,従来の“感覚刺激と応答としての行動”でとらえられた一方向性の並列階層的な構造を想定すると無限に自発性の起源をさかのぼる必要が生じてしまう.しかし,この研究で,準備活動を生成する回帰性の神経回路網の脳に存在する可能性が比較的単純な神経系をもつと考えられてきたザリガニで示された.ザリガニは行動を遂行しているときでも単一ニューロンレベルでシナプス機構を追究することが可能であり,神経生理学的な過去の知見も豊富である.今後,回帰性の神経回路網の動作機構が単一細胞レベルにおいて明らかになることが期待される.
略歴:2010年 北海道大学大学院理学研究科博士課程 修了,同年より同 学術研究員(ポスドク).
研究テーマ:動物生理学,神経生物学,自発性の行動発現の神経機構.
抱負:動物の適応的な行動発現のしくみを比較生物学的に明らかにしていきたい.
高畑 雅一(Masakazu Takahata)
北海道大学大学院理学研究院 教授.
研究室URL:http://crayfish3.sci.hokudai.ac.jp/index.html
© 2011 加賀谷勝史・高畑雅一 Licensed under CC 表示 2.1 日本
(北海道大学大学院理学研究院 生物科学部門)
email:加賀谷勝史
DOI: 10.7875/first.author.2011.081
Sequential synaptic excitation and inhibition shape readiness discharge for voluntary behavior.
Katsushi Kagaya, Masakazu Takahata
Science, 332, 365-368 (2011)
要 約
動物はどのように随意行動を開始するのか? 神経科学において鍵となる現象が脳の特定領域で起こる準備神経活動である.しかし,その活動を媒介する特定のニューロンおよびシナプス機構については不明であった.筆者らがアメリカザリガニの脳において発見した準備発射活動は,逐次的なシナプスの興奮と抑制により形成されることが判明した.準備発射ニューロンは軸索の側枝を脳に伸ばし,回帰性の神経回路網を形成する局所介在ニューロンを賦活すると考えられた.したがって,筆者らは,下位神経節からのフィードバック信号なしに逐次的なシナプスの賦活により準備発射が形成されているという回帰性の神経回路網を提案した.この神経回路網は自己生成的な随意行動の開始の信号処理に適している.
はじめに
随意行動の開始に先行して脳の特定領域で特徴的な神経活動の変化が起こる.活動電位として記録される準備発射は霊長類や齧歯類において多くの報告がなされてきた1).しかし,準備発射を形成する脳内ニューロンの樹状突起における膜電位のダイナミクスについては不明であった.アメリカザリガニ(Procambarus clarkii)の脳における準備発射は随意行動の開始のための準備活動の生成における細胞の機構を理解する手がかりをあたえると考えられる.ザリガニでは準備発射は囲食道縦連合において記録される2)(図1a).囲食道縦連合には脳内ニューロンの軸索が走行しており,ザリガニの脳の活動はこの囲食道縦連合を介して下行性ニューロンにより胸部へと伝えられる(図1b).胸部には歩行パターンを生成する中枢パターン発生器が存在する3).準備発射は行動開始の数秒まえに現われ歩行が開始すると消える2).この一過的なスパイク活動は準備発射に共通する特徴である.しかし,どのようなシナプス機構およびニューロンがこの活動の形成にかかわるのかは不明であった.
1.歩行しているザリガニの脳の単一ニューロンからの細胞内記録および細胞内染色
随意歩行の開始の神経生理学的な機構を明らかにする目的で,球形のトレッドミルを歩行するザリガニの脳の単一細胞からの細胞内記録を行った.外部からの感覚刺激はあたえられず,したがって,ザリガニは自ら歩行を開始,維持,停止することができる.実験の結果,合計で69個のニューロンからの細胞内記録および細胞内染色に成功した.同定されたニューロンの機能的な分類は歩行の開始と停止のタイミングにもとづいて行った.ザリガニの歩脚は8本ある.歩脚の筋活動はほぼ同時に(65.89±6.40ミリ秒の時間幅のなかで)活動の上がることが判明している4).そのため,1本の歩脚の長腕節屈筋から記録をとり筋活動の変化から歩行の開始と停止のタイミングを決定した.シナプス活動およびスパイク活動が歩行の開始に先行すること500ミリ秒以上である場合を“先行する”と定義した.形態的には下行性ニューロンと局所介在ニューロンに分類した.下行性ニューロンは囲食道縦連合をとおり胸部へと軸索を伸ばす.局所介在ニューロンは突起の投射を脳に限局している.昆虫では歩行制御の脳内部位として中心複合体が関連すると考えられている5,6).ザリガニの脳における相同部位7) のなかで随意歩行の制御にかかわる部位がどこかを明らかにする目的で脳内ニューロンの突起投射のパターンを調査した.細胞内染色は細胞内記録を行ったあとにガラス管微小電極に充填した蛍光色素Lucifer yellowにより行った.
2.準備活動ニューロンのシナプス活動およびスパイク活動
行動を遂行しているザリガニの準備発射ニューロンからシナプス活動を記録することに成功した.ザリガニは多様な歩行パターンを示したが,準備発射ニューロンの膜電位は歩行パターンとは関係なく,歩行の開始前には脱分極性の変化,開始後には過分極性の変化を示した.さらに,電流注入実験により,この逐次的な膜電位の変化は直接的な興奮性シナプス入力および抑制性シナプス入力によることが判明した.また,膜コンダクタンス変化の解析から,この抑制性の入力は行動開始の直後と数秒後とで樹状突起の異なる入力部位において受け取られる可能性を示した.また,脱分極のときにはシナプスノイズの増加も認められた.これらの結果は,興奮性シナプス入力および2つ以上の抑制性シナプス入力を前シナプスニューロンから受け取ることで準備発射が生成されており,個々のニューロンにおける内発的な活動電位の発生という機構によるものではないことを示していた.
3.準備活動ニューロンの脳における投射パターン
準備発射ニューロンは細胞体を前大脳の腹側前方部にもち樹状突起を前方および後方の内側ニューロピルに投射することが判明した.このニューロンは下行性ニューロンであり軸索は囲食道縦連合のWiersma領域70に伸びていた8).この領域には電気刺激をくわえると歩行をひき起こすユニットの含まれていることが過去の研究でわかっている2,9).また,この領域には準備活動ユニットが含まれ,軸索束の切断断端から蛍光色素を注入して脳における細胞を染色したところ細胞体の位置や樹状突起の形態などの形態的な特徴の一致するニューロンの存在することが判明した2).さらに,後大脳の食道ニューロピルに軸索の側枝の投射がみられた.これは,胸部だけでなく脳にも準備発射をうけるニューロンの存在することを示し,下位神経節を介さずとも脳において歩行の開始の情報が得られることを示していた.
4.歩行の維持および停止において動員される下行性ニューロン
筆者らは,以前の細胞外記録法を用いた報告で,準備発射活動にくわえて歩行の維持および停止にかかわる下行性の活動の存在することを示した.今回,細胞内記録および細胞内染色を用いてそれぞれを代表する下行性ニューロンの同定にも成功した.準備活動ニューロンと共通した特徴として,歩行の維持および停止にかかわるニューロンも後大脳の食道ニューロピルに軸索の側枝をもっていることが判明した.このことは,脳においては下位神経節からのフィードバックなしに行動の開始,維持,停止という情報を得ている可能性を示していた.
5.随意歩行の開始に関連する下行性ニューロンの樹状突起の投射パターン
スパイク活動から分類した歩行の開始,維持,停止の3つのタイプにくわえて,脳のどの領域でシナプスの賦活が起こっているのかを調べるため,シナプス活動の変化が歩行の開始の前後で観察された45個の下行性ニューロンを同定した.先行したシナプス活動の変化を示すものを先行型ニューロン,ほぼ同時に変化を示すものを随伴型ニューロン,遅れてあるいは変化を示さないものを遅延型ニューロンとして分類し,それぞれの脳における樹状突起の投射部位を調べた.その結果,先行型ニューロンと随伴型ニューロンは樹状突起を後大脳あるいは中大脳に比べ前大脳に多くもつ傾向を示した.一方で,遅延型ニューロンは中大脳と後大脳に樹状突起を多くもつ傾向があった.したがって,前大脳の内側部に限局された投射とシナプス活動から随意歩行の開始にかかわるシナプスの賦活は前大脳の内側部で起こっているものと考えられた.
6.並列した下行性経路を賦活する回帰性の神経回路網
今回,発見された準備発射ニューロンは興奮性シナプス入力のあとに抑制性シナプス入力をうけていることが判明した.ひとつの可能性として,この抑制性シナプス入力が継続ニューロンによりひき起こされていることが考えられた.準備発射ニューロンおよび継続ニューロンは脳に軸索の側枝をもっており,それぞれの活動を脳において出力するものと考えられた.準備発射ニューロンの樹状突起と継続ニューロンの軸索の側枝の出力領域は重なってはいなかった.多シナプス接続を介して信号が伝えられている,つまり,脳において局所介在ニューロンにより並列的かつ逐次的に下行性回路を賦活しているものと考えられた(図2).
さらに,スパイク発生型およびスパイク非発生型の局所介在ニューロンからの細胞内記録および細胞内染色に成功し,後大脳の食道ニューロピルと前大脳の内側部に突起をもつニューロンを見い出した.これらのシナプスの接続関係については不明であるが,作業仮説として,これら局所介在ニューロンと下行性ニューロンとが前大脳と後大脳においてループを構成するという回帰性の神経回路網が存在すると考えた(図2).
いずれにしても,筆者らのデータは,ザリガニにおいて自己生成的な随意歩行に関連した脳内ニューロンの生理学的および形態学的な特徴を明らかにし,それらが個々の内発的な機構によってではなく,前シナプスニューロンにより賦活されていることを示した.
おわりに
従来,動物の行動の発現機構,とりわけ無脊椎動物における行動の発現機構は“感覚刺激に対する応答としての行動”という視点により実験的に解析されて多大な知見が得られてきた.そこでは“感覚入力が司令ニューロンに収斂し運動出力へといたる”という一方向性の並列階層的な機構が想定され研究されてきた.一方で,多くの動物は明確に定義された外部からの感覚刺激がなくても自発的に行動を発現し制御している.しかし,自発性の行動発現の単一細胞レベルでの神経回路の動作機構となると無脊椎動物ではほとんど報告がなく,また,そのシナプス機構となると脊椎動物でもまったく未解明である.
随意行動の発現機構を研究する場合,この研究のように運動出力の側からさかのぼってその機構を明らかにしていくのが近道であるが,従来の“感覚刺激と応答としての行動”でとらえられた一方向性の並列階層的な構造を想定すると無限に自発性の起源をさかのぼる必要が生じてしまう.しかし,この研究で,準備活動を生成する回帰性の神経回路網の脳に存在する可能性が比較的単純な神経系をもつと考えられてきたザリガニで示された.ザリガニは行動を遂行しているときでも単一ニューロンレベルでシナプス機構を追究することが可能であり,神経生理学的な過去の知見も豊富である.今後,回帰性の神経回路網の動作機構が単一細胞レベルにおいて明らかになることが期待される.
文 献
- Roskies, A. L.: How does neuroscience affect our conception of volition? Annu. Rev. Neurosci., 33, 109-130 (2010)[PubMed]
- Kagaya, K. & Takahata, M.: Readiness discharge for spontaneous initiation of walking in crayfish. J. Neurosci., 30, 1348-1362 (2010)[PubMed]
- Chrachri, A. & Clarac, F.: Fictive locomotion in the fourth thoracic ganglion of the crayfish, Procambarus clarkii. J. Neurosci., 10, 707-719 (1990)[PubMed]
- Chikamoto, K., Kagaya K., Takahata, M.: Electromyographic characterization of walking behavior initiated spontaneously in crayfish. Zoolog. Sci., 25, 783-792 (2008)[PubMed]
- Martin, J. R., Raabe, T., Heisenberg M.: Central complex substructures are required for the maintenance of locomotor activity in Drosophila melanogaster. J. Comp. Physiol. A, 185, 277-288 (1999)[PubMed]
- Bender, J. A., Pollack, A. J., Ritzmann, R. E.: Neural activity in the central complex of the insect brain is linked to locomotor changes. Curr. Biol., 20, 921-926 (2010)[PubMed]
- Sandeman, D., Sandeman, R., Derby, C. et al.: Morphology of the brain of crayfish, crabs, and spiny lobsters: A common nomenclature for homologous structures. Biol. Bull., 183, 304-326 (1992)
- Wiersma, C. A. G.: On the functional connections of single units in the central nervous system of the crayfish, Procambarus clarkii Girard. J. Comp. Neurol., 110, 421-471 (1958)[PubMed]
- Bowerman, R. F. & Larimer, J. L.: Command fibres in the circumoesophageal connectives of crayfish II phasic fibres. J. Exp. Biol., 60, 119-134 (1974)
著者プロフィール
略歴:2010年 北海道大学大学院理学研究科博士課程 修了,同年より同 学術研究員(ポスドク).
研究テーマ:動物生理学,神経生物学,自発性の行動発現の神経機構.
抱負:動物の適応的な行動発現のしくみを比較生物学的に明らかにしていきたい.
高畑 雅一(Masakazu Takahata)
北海道大学大学院理学研究院 教授.
研究室URL:http://crayfish3.sci.hokudai.ac.jp/index.html
© 2011 加賀谷勝史・高畑雅一 Licensed under CC 表示 2.1 日本
