E4BP4はT細胞におけるインターロイキン10の産生を制御する鍵となる転写因子である
本村泰隆・久保允人
(東京理科大学生命科学研究所 分子病態学研究部門)
email:本村泰隆,久保允人
DOI: 10.7875/first.author.2011.078
The transcription factor E4BP4 regulates the production of IL-10 and IL-13 in CD4+T cells.
Yasutaka Motomura, Hiroshi Kitamura, Atsushi Hijikata, Yuko Matsunaga, Koichiro Matsumoto, Hiromasa Inoue, Koji Atarashi, Shohei Hori, Hiroshi Watarai, Jinfang Zhu, Masaru Taniguchi, Masato Kubo
Nature Immunology, 12, 450-459 (2011)
われわれの身体に備わった免疫系は炎症性サイトカインを誘導することにより免疫反応を増強させている.しかしながら,この反応が過剰になると自己の正常な組織までも攻撃し自己免疫疾患にいたる.こうした過剰な攻撃を抑制するため免疫系は反応を抑制する分子機構ももちあわせている.インターロイキン10は炎症反応に対し抑制的にはたらくサイトカインとして知られ,さまざまなT細胞サブセットのなかでおもにTh2細胞から産生される.しかしながら,Th1細胞,Th17細胞やナチュラルキラーT細胞でも,炎症が慢性化するなど細胞のおかれた環境の変化によりインターロイキン10の産生能を獲得する.ところが,これらの細胞でインターロイキン10の発現がどのように制御されているのか,その分子機構についてはこれまでほとんど明らかにされていなかった.今回の研究で筆者らは,これまで,時計遺伝子のレギュレーターとして同定されていた基本ロイシンジッパー転写因子E4BP4が,T細胞におけるインターロイキン10の産生に必須の転写因子であることを世界にさきがけて証明した.また,E4BP4の欠損はインターロイキン10の産生低下にともない自己免疫性の炎症性大腸炎を誘導することから,自己免疫性の炎症におけるE4BP4の生理学的な意義の重要性がうかがえた.
免疫系はさまざまな病原性細菌やウイルスによる感染から生体を守るため,炎症性サイトカインを誘導することで生体防御反応を増強する一方,これらの反応にいきすぎのないよう,そして,腸に生息する細菌や食物といった異物に対し過剰に反応することがないよう,これを抑制する分子機構も兼ね備えている.この免疫応答のバランスを適切に保つ過程はサイトカインにより制御されている.これらのサイトカインはおもにヘルパーT細胞より産生され,ヘルパーT細胞は産生するサイトカインによりTh1細胞,Th2細胞,Th17細胞などのサブセットに分類されている.これらのヘルパーT細胞はいずれもかつて抗原刺激をうけたことのないナイーブT細胞が特定のサイトカイン環境において抗原刺激をうけることにより誘導されることから,分化という観点でとらえられている.そのため,ヘルパーT細胞はいちど獲得したサイトカインの産生能を変化させることなく維持しメモリーT細胞として残るか,ある一定の期間にわたり増殖したのち死んでいくことにより反応を終焉に導いているものと考えられ,分化の終着点にいると考えられていた.つまり従来は,いちどTh1細胞へと分化したT細胞はインターフェロンγやIL-2(IL:interleukin,インターロイキン)を産生するが,Th2細胞やTh17細胞の産生するサイトカインを産生することはないと考えられていた.
ところが,寄生虫感染の慢性化したマウスにおいてTh1細胞がIL-10を産生すること1-3),IL-27により刺激されたTh1細胞およびTh17細胞はIL-10を産生できるようになること4,5),IL-18による刺激はTh1細胞からIL-13の産生を誘導すること6),などの報告が蓄積されるにしたがい,Th1細胞,Th2細胞,Th17細胞は必ずしも分化の終着点ではなく,これまで産生のみられなかったサイトカインを産生する余地を残しているという“可塑性”の概念が浮上してきた.しかしながら,この可塑性を制御する分子機構についてはこれまでほとんど研究対象とされることもなく,“本来はIL-10やIL-13を産生しないTh1細胞がどのようにしてIL-10の産生能を獲得するのか?”という疑問については未解決のまま残されてきた.とくに,IL-10の産生制御機構はその抗炎症性サイトカインとしてのはたらきから炎症性疾患や自己免疫疾患の抑制的な制御を考察するうえでも重要な課題といえる.
筆者らは,概日リズムを制御する時計遺伝子のレギュレーターとして知られていた転写因子E4BP4がTh1細胞におけるIL-10およびIL-13の可塑的な発現を制御するタンパク質であったという発見を起点として,じつは,このE4BP4がT細胞におけるIL-10の産生を制御するうえで必須のタンパク質であることを証明した.ここでは,発見にいたる経緯などを含め,IL-10の産生におけるE4BP4の重要性を紹介する.
炎症に広くかかわるサイトカインであるインターフェロンγを産生するTh1細胞の可塑性を検討する目的で,Th1細胞に1週間の間隔で4回の抗原刺激をくり返すことで慢性的な抗原刺激をあたえ可塑性が誘導されるかどうか検討した.慢性的な抗原刺激はTh1細胞において,通常では産生されないIL-10およびIL-13を誘導することを見い出した.そこで,IL-10およびIL-13を産生しないTh1細胞と産生するようになったTh1細胞とのあいだで遺伝子の発現パターンを比較することで,この可塑性を制御するタンパク質の同定を試みた.この比較解析から浮上した16の候補タンパク質をTh1細胞に発現させたところ,時計遺伝子の転写因子として知られるE4BP4を発現させることによりTh1細胞はIL-13だけでなくIL-10も同時に産生した.また,E4BP4ノックアウトマウスに由来するT細胞に慢性的な抗原刺激をくわえることにより可塑的な状況をあたえたTh1細胞では,IL-10およびIL-13の産生がまったくみられなくなった.これらのことから,慢性的な抗原刺激により誘導されるE4BP4は,Th1細胞においてIL-13およびIL-10の産生を可塑的に制御するタンパク質であることが証明された.
E4BP4は松果体細胞において24時間の周期で発現の変動する転写因子であり,生体リズムを制御する概日時計の進行を負に制御することが知られていた.E4BP4は松果体において“時計の針”にあたる機能を担うタンパク質Periodのはたらきを抑えることにより時間合わせをするタンパク質として報告された7).そのため,概日リズムにおけるはたらきに長いあいだ注目が集まっていた.一方,免疫系においても同一のタンパク質がNfil3という別名で示されていた.Nfil3は最初に,T細胞においてIL-3遺伝子のプロモーターに結合しその発現を誘導するものとして同定された8).また,プロB細胞においてもその発現はIL-3により誘導され,Raf/MAPキナーゼ経路あるいはPI3キナーゼ経路を介してアポトーシスを抑制する機能をもつ9).ヘルパーT細胞においてはTh2細胞に特異的に発現の亢進するタンパク質であることがマイクロアレイ解析により既知の事実となっていた10,11).Th2細胞におけるE4BP4の発現は抗IL-4抗体およびSTAT6ノックアウトマウスに由来するT細胞において阻害されていることから,その発現はIL-4/STAT6シグナル経路により制御されていることまでは明らかになった.しかしながら,その生理学的な意義についてはこれまでまったく明らかにされてこなかった.一方,最近の知見では,E4BP4がナチュラルキラー細胞の分化の過程に重要なはたらきをもつことが示されている12,13).マウスの自然免疫系ではE4BP4はIL-15により誘導されナチュラルキラー細胞の分化に必須のId2の発現を誘導しているため,E4BP4ノックアウトマウスではId2の発現が誘導されずナチュラルキラー細胞も分化できない.このように,E4BP4は生体維持機構のなかのさまざまな局面ではたらく多機能な転写因子であることがわかっていた.
さきに述べたように,E4BP4はTh1細胞により可塑的に産生されるIL-10およびIL-13の産生にかかわることは明らかになったが,これら2つのサイトカインは通常はおもにTh2細胞から産生される.そこで,E4BP4ノックアウトマウスに由来するTh2細胞におけるIL-10およびIL-13の産生能を解析した.驚いたことに,そこで得られた結果はTh1細胞における結果とは大きく異なるものであった.E4BP4を欠損したTh2細胞では,IL-13の産生の低下は認められなかったにもかかわらず,IL-10の産生はほぼ完全に消失していた.このことから,E4BP4はIL-13の産生に関してTh2細胞とTh1細胞とで異なる役割をもつこと,一方,IL-10の産生に関してはTh1細胞とTh2細胞とのあいだで共通で,その産生系において鍵となる重要な転写因子であることが明らかとなった.また,従来からE4BP4の発現はTh2細胞において特異的に高いことが知られていたわけだが,同様に,Th2細胞はIL-10のおもな産生細胞としても知られている.このことは,Th2細胞におけるE4BP4の生理学的な意義はIL-10の産生制御にあることを示唆していた.
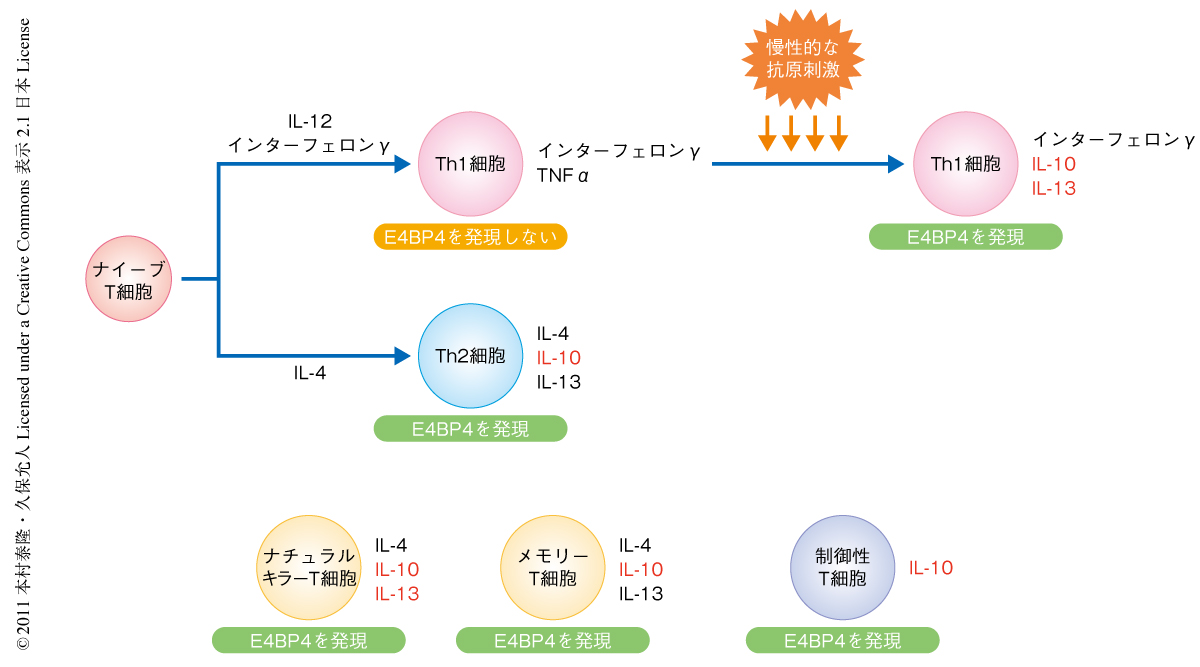
IL-10はさまざまなT細胞サブセットからも産生される.そこで,E4BP4ノックアウトマウスより単離したナチュラルキラーT細胞,メモリーT細胞,制御性T細胞におけるIL-10の産生を検討した.ナチュラルキラーT細胞のなかでIL-17RB(IL-25に対する受容体)を発現するサブセットはIL-25により刺激をうけさまざまなサイトカインを産生することが報告されている14).αガラクトシルセラミドおよびIL-25で刺激したナチュラルキラーT細胞はIL-10を産生するが,この産生はE4BP4ノックアウトマウスより単離した細胞では顕著に低下していた.また,メモリーT細胞や制御性T細胞においてもE4BP4の欠損にともないIL-10の発現は低下傾向にあった.以上の結果から,E4BP4は広義にT細胞におけるIL-10の産生を制御する転写因子として機能していることが明らかになった(図1).
IL-10は炎症反応に対し抑制的にはたらくことが知られている.このことは,IL-10ノックアウトマウスが炎症性大腸炎を自然発症することからも証明されている15).一方,E4BP4ノックアウトマウスは下痢をともなう軽度の炎症性大腸炎様の症状を示す.このことから,E4BP4は生体においてIL-10の産生を介して炎症反応を抑制していることが考えられた.生体におけるE4BP4の役割を明らかにするため,ナイーブCD4陽性T細胞をRAG-1ノックアウトマウスに移入することで誘導される炎症性大腸炎のモデルマウスを用いて解析を行った.野生型のT細胞を移入した炎症性大腸炎のモデルマウスよりもE4BP4を欠失したT細胞を移入したモデルマウスのほうが炎症性大腸炎の特徴的な症状である体重減少や下痢が顕著に増悪し,大腸における上皮の損傷および陰窩構造の破壊が進行していた.また,炎症部位におけるCD4陽性T細胞からのIL-10の産生がE4BP4の欠失により顕著に低下していた.このことから,E4BP4は大腸炎における炎症反応をIL-10の産生を介して抑制していることが示唆された.
さらに,多発性硬化症のモデルマウスである実験的自己免疫性脳脊髄炎においてもE4BP4の役割を検討した.ミエリン蛋白質を免疫することでE4BP4ノックアウトマウスに実験的自己免疫性脳脊髄炎を誘導したところ,その発症時期は野生型マウスと差が認められなかったが,E4BP4ノックアウトマウスは野生型マウスに比べ症状が徐々に悪化していき慢性化していった.そのとき,E4BP4ノックアウトマウスではリンパ節のCD4陽性T細胞からのIL-10の産生が顕著に低下していた.したがって,E4BP4はIL-10を介して実験的自己免疫性脳脊髄炎における慢性的な炎症反応を抑制していることが示唆された.この2つのモデルマウスを用いた実験結果から,E4BP4は炎症性疾患や自己免疫疾患といった病態において炎症反応を沈静化させる役割をもつことが明らかになった.
Th1細胞においてIL-10およびIL-13の可塑的な産生を制御するタンパク質として発見されたE4BP4は,T細胞に共通のIL-10レギュレーターとしての役割のあることが明らかになった.しかしながら最近になり,IL-27の刺激によりIL-10を産生する細胞傷害性T細胞や制御性T細胞におけるIL-10の産生にはBlimp-1やIRF4の関与していることが報告されている16,17).CD4陽性CD25陽性LAG3陽性制御性T細胞など特殊な制御性T細胞の集団ではEgr-2がIL-10のレギュレーターとしてはたらくという報告もある18).したがって,これまでまったく明らかにされてこなかったIL-10の産生について急速に新しい知見が得られていることはまちがいないが,E4BP4とほかのIL-10レギュレーターとの関係については筆者らが答えていかなければならない新しい課題として残されている.これらの課題に応えていくことにより,IL-10の産生機構の理解を深め,過剰な炎症反応をひき起こすさまざまな生活習慣病や自己免疫疾患に対して新しい視点からの治療の実現や根本的な予防を提案することが可能となることを期待する.
略歴:2010年 東京医科歯科大学大学院生命情報科学教育部博士課程(理学)修了,同年より東京理科大学生命科学研究所 ポストドクトラル研究員.
研究テーマ:T細胞におけるサイトカイン遺伝子の制御機構の解明.
久保 允人(Masato Kubo)
東京理科大学生命科学研究所 教授,理化学研究所免疫アレルギー科学総合研究センター チームリーダー.
研究室URL:http://www.rs.noda.tus.ac.jp/~ribsjm/kubolab/
© 2011 本村泰隆・久保允人 Licensed under CC 表示 2.1 日本
(東京理科大学生命科学研究所 分子病態学研究部門)
email:本村泰隆,久保允人
DOI: 10.7875/first.author.2011.078
The transcription factor E4BP4 regulates the production of IL-10 and IL-13 in CD4+T cells.
Yasutaka Motomura, Hiroshi Kitamura, Atsushi Hijikata, Yuko Matsunaga, Koichiro Matsumoto, Hiromasa Inoue, Koji Atarashi, Shohei Hori, Hiroshi Watarai, Jinfang Zhu, Masaru Taniguchi, Masato Kubo
Nature Immunology, 12, 450-459 (2011)
要 約
われわれの身体に備わった免疫系は炎症性サイトカインを誘導することにより免疫反応を増強させている.しかしながら,この反応が過剰になると自己の正常な組織までも攻撃し自己免疫疾患にいたる.こうした過剰な攻撃を抑制するため免疫系は反応を抑制する分子機構ももちあわせている.インターロイキン10は炎症反応に対し抑制的にはたらくサイトカインとして知られ,さまざまなT細胞サブセットのなかでおもにTh2細胞から産生される.しかしながら,Th1細胞,Th17細胞やナチュラルキラーT細胞でも,炎症が慢性化するなど細胞のおかれた環境の変化によりインターロイキン10の産生能を獲得する.ところが,これらの細胞でインターロイキン10の発現がどのように制御されているのか,その分子機構についてはこれまでほとんど明らかにされていなかった.今回の研究で筆者らは,これまで,時計遺伝子のレギュレーターとして同定されていた基本ロイシンジッパー転写因子E4BP4が,T細胞におけるインターロイキン10の産生に必須の転写因子であることを世界にさきがけて証明した.また,E4BP4の欠損はインターロイキン10の産生低下にともない自己免疫性の炎症性大腸炎を誘導することから,自己免疫性の炎症におけるE4BP4の生理学的な意義の重要性がうかがえた.
はじめに
免疫系はさまざまな病原性細菌やウイルスによる感染から生体を守るため,炎症性サイトカインを誘導することで生体防御反応を増強する一方,これらの反応にいきすぎのないよう,そして,腸に生息する細菌や食物といった異物に対し過剰に反応することがないよう,これを抑制する分子機構も兼ね備えている.この免疫応答のバランスを適切に保つ過程はサイトカインにより制御されている.これらのサイトカインはおもにヘルパーT細胞より産生され,ヘルパーT細胞は産生するサイトカインによりTh1細胞,Th2細胞,Th17細胞などのサブセットに分類されている.これらのヘルパーT細胞はいずれもかつて抗原刺激をうけたことのないナイーブT細胞が特定のサイトカイン環境において抗原刺激をうけることにより誘導されることから,分化という観点でとらえられている.そのため,ヘルパーT細胞はいちど獲得したサイトカインの産生能を変化させることなく維持しメモリーT細胞として残るか,ある一定の期間にわたり増殖したのち死んでいくことにより反応を終焉に導いているものと考えられ,分化の終着点にいると考えられていた.つまり従来は,いちどTh1細胞へと分化したT細胞はインターフェロンγやIL-2(IL:interleukin,インターロイキン)を産生するが,Th2細胞やTh17細胞の産生するサイトカインを産生することはないと考えられていた.
ところが,寄生虫感染の慢性化したマウスにおいてTh1細胞がIL-10を産生すること1-3),IL-27により刺激されたTh1細胞およびTh17細胞はIL-10を産生できるようになること4,5),IL-18による刺激はTh1細胞からIL-13の産生を誘導すること6),などの報告が蓄積されるにしたがい,Th1細胞,Th2細胞,Th17細胞は必ずしも分化の終着点ではなく,これまで産生のみられなかったサイトカインを産生する余地を残しているという“可塑性”の概念が浮上してきた.しかしながら,この可塑性を制御する分子機構についてはこれまでほとんど研究対象とされることもなく,“本来はIL-10やIL-13を産生しないTh1細胞がどのようにしてIL-10の産生能を獲得するのか?”という疑問については未解決のまま残されてきた.とくに,IL-10の産生制御機構はその抗炎症性サイトカインとしてのはたらきから炎症性疾患や自己免疫疾患の抑制的な制御を考察するうえでも重要な課題といえる.
筆者らは,概日リズムを制御する時計遺伝子のレギュレーターとして知られていた転写因子E4BP4がTh1細胞におけるIL-10およびIL-13の可塑的な発現を制御するタンパク質であったという発見を起点として,じつは,このE4BP4がT細胞におけるIL-10の産生を制御するうえで必須のタンパク質であることを証明した.ここでは,発見にいたる経緯などを含め,IL-10の産生におけるE4BP4の重要性を紹介する.
1.Th1細胞の可塑性を制御するタンパク質E4BP4の同定
炎症に広くかかわるサイトカインであるインターフェロンγを産生するTh1細胞の可塑性を検討する目的で,Th1細胞に1週間の間隔で4回の抗原刺激をくり返すことで慢性的な抗原刺激をあたえ可塑性が誘導されるかどうか検討した.慢性的な抗原刺激はTh1細胞において,通常では産生されないIL-10およびIL-13を誘導することを見い出した.そこで,IL-10およびIL-13を産生しないTh1細胞と産生するようになったTh1細胞とのあいだで遺伝子の発現パターンを比較することで,この可塑性を制御するタンパク質の同定を試みた.この比較解析から浮上した16の候補タンパク質をTh1細胞に発現させたところ,時計遺伝子の転写因子として知られるE4BP4を発現させることによりTh1細胞はIL-13だけでなくIL-10も同時に産生した.また,E4BP4ノックアウトマウスに由来するT細胞に慢性的な抗原刺激をくわえることにより可塑的な状況をあたえたTh1細胞では,IL-10およびIL-13の産生がまったくみられなくなった.これらのことから,慢性的な抗原刺激により誘導されるE4BP4は,Th1細胞においてIL-13およびIL-10の産生を可塑的に制御するタンパク質であることが証明された.
2.基本ロイシンジッパー転写因子E4BP4
E4BP4は松果体細胞において24時間の周期で発現の変動する転写因子であり,生体リズムを制御する概日時計の進行を負に制御することが知られていた.E4BP4は松果体において“時計の針”にあたる機能を担うタンパク質Periodのはたらきを抑えることにより時間合わせをするタンパク質として報告された7).そのため,概日リズムにおけるはたらきに長いあいだ注目が集まっていた.一方,免疫系においても同一のタンパク質がNfil3という別名で示されていた.Nfil3は最初に,T細胞においてIL-3遺伝子のプロモーターに結合しその発現を誘導するものとして同定された8).また,プロB細胞においてもその発現はIL-3により誘導され,Raf/MAPキナーゼ経路あるいはPI3キナーゼ経路を介してアポトーシスを抑制する機能をもつ9).ヘルパーT細胞においてはTh2細胞に特異的に発現の亢進するタンパク質であることがマイクロアレイ解析により既知の事実となっていた10,11).Th2細胞におけるE4BP4の発現は抗IL-4抗体およびSTAT6ノックアウトマウスに由来するT細胞において阻害されていることから,その発現はIL-4/STAT6シグナル経路により制御されていることまでは明らかになった.しかしながら,その生理学的な意義についてはこれまでまったく明らかにされてこなかった.一方,最近の知見では,E4BP4がナチュラルキラー細胞の分化の過程に重要なはたらきをもつことが示されている12,13).マウスの自然免疫系ではE4BP4はIL-15により誘導されナチュラルキラー細胞の分化に必須のId2の発現を誘導しているため,E4BP4ノックアウトマウスではId2の発現が誘導されずナチュラルキラー細胞も分化できない.このように,E4BP4は生体維持機構のなかのさまざまな局面ではたらく多機能な転写因子であることがわかっていた.
3.E4BP4はT細胞における共通のIL-10レギュレーターである
さきに述べたように,E4BP4はTh1細胞により可塑的に産生されるIL-10およびIL-13の産生にかかわることは明らかになったが,これら2つのサイトカインは通常はおもにTh2細胞から産生される.そこで,E4BP4ノックアウトマウスに由来するTh2細胞におけるIL-10およびIL-13の産生能を解析した.驚いたことに,そこで得られた結果はTh1細胞における結果とは大きく異なるものであった.E4BP4を欠損したTh2細胞では,IL-13の産生の低下は認められなかったにもかかわらず,IL-10の産生はほぼ完全に消失していた.このことから,E4BP4はIL-13の産生に関してTh2細胞とTh1細胞とで異なる役割をもつこと,一方,IL-10の産生に関してはTh1細胞とTh2細胞とのあいだで共通で,その産生系において鍵となる重要な転写因子であることが明らかとなった.また,従来からE4BP4の発現はTh2細胞において特異的に高いことが知られていたわけだが,同様に,Th2細胞はIL-10のおもな産生細胞としても知られている.このことは,Th2細胞におけるE4BP4の生理学的な意義はIL-10の産生制御にあることを示唆していた.
IL-10はさまざまなT細胞サブセットからも産生される.そこで,E4BP4ノックアウトマウスより単離したナチュラルキラーT細胞,メモリーT細胞,制御性T細胞におけるIL-10の産生を検討した.ナチュラルキラーT細胞のなかでIL-17RB(IL-25に対する受容体)を発現するサブセットはIL-25により刺激をうけさまざまなサイトカインを産生することが報告されている14).αガラクトシルセラミドおよびIL-25で刺激したナチュラルキラーT細胞はIL-10を産生するが,この産生はE4BP4ノックアウトマウスより単離した細胞では顕著に低下していた.また,メモリーT細胞や制御性T細胞においてもE4BP4の欠損にともないIL-10の発現は低下傾向にあった.以上の結果から,E4BP4は広義にT細胞におけるIL-10の産生を制御する転写因子として機能していることが明らかになった(図1).
4.炎症性大腸炎および実験的自己免疫性脳脊髄炎におけるE4BP4の役割
IL-10は炎症反応に対し抑制的にはたらくことが知られている.このことは,IL-10ノックアウトマウスが炎症性大腸炎を自然発症することからも証明されている15).一方,E4BP4ノックアウトマウスは下痢をともなう軽度の炎症性大腸炎様の症状を示す.このことから,E4BP4は生体においてIL-10の産生を介して炎症反応を抑制していることが考えられた.生体におけるE4BP4の役割を明らかにするため,ナイーブCD4陽性T細胞をRAG-1ノックアウトマウスに移入することで誘導される炎症性大腸炎のモデルマウスを用いて解析を行った.野生型のT細胞を移入した炎症性大腸炎のモデルマウスよりもE4BP4を欠失したT細胞を移入したモデルマウスのほうが炎症性大腸炎の特徴的な症状である体重減少や下痢が顕著に増悪し,大腸における上皮の損傷および陰窩構造の破壊が進行していた.また,炎症部位におけるCD4陽性T細胞からのIL-10の産生がE4BP4の欠失により顕著に低下していた.このことから,E4BP4は大腸炎における炎症反応をIL-10の産生を介して抑制していることが示唆された.
さらに,多発性硬化症のモデルマウスである実験的自己免疫性脳脊髄炎においてもE4BP4の役割を検討した.ミエリン蛋白質を免疫することでE4BP4ノックアウトマウスに実験的自己免疫性脳脊髄炎を誘導したところ,その発症時期は野生型マウスと差が認められなかったが,E4BP4ノックアウトマウスは野生型マウスに比べ症状が徐々に悪化していき慢性化していった.そのとき,E4BP4ノックアウトマウスではリンパ節のCD4陽性T細胞からのIL-10の産生が顕著に低下していた.したがって,E4BP4はIL-10を介して実験的自己免疫性脳脊髄炎における慢性的な炎症反応を抑制していることが示唆された.この2つのモデルマウスを用いた実験結果から,E4BP4は炎症性疾患や自己免疫疾患といった病態において炎症反応を沈静化させる役割をもつことが明らかになった.
おわりに
Th1細胞においてIL-10およびIL-13の可塑的な産生を制御するタンパク質として発見されたE4BP4は,T細胞に共通のIL-10レギュレーターとしての役割のあることが明らかになった.しかしながら最近になり,IL-27の刺激によりIL-10を産生する細胞傷害性T細胞や制御性T細胞におけるIL-10の産生にはBlimp-1やIRF4の関与していることが報告されている16,17).CD4陽性CD25陽性LAG3陽性制御性T細胞など特殊な制御性T細胞の集団ではEgr-2がIL-10のレギュレーターとしてはたらくという報告もある18).したがって,これまでまったく明らかにされてこなかったIL-10の産生について急速に新しい知見が得られていることはまちがいないが,E4BP4とほかのIL-10レギュレーターとの関係については筆者らが答えていかなければならない新しい課題として残されている.これらの課題に応えていくことにより,IL-10の産生機構の理解を深め,過剰な炎症反応をひき起こすさまざまな生活習慣病や自己免疫疾患に対して新しい視点からの治療の実現や根本的な予防を提案することが可能となることを期待する.
文 献
- Shaw, M. H., Freeman, G. J., Scott, M. F. et al.: Tyk2 negatively regulates adaptive Th1 immunity by mediating IL-10 signaling and promoting IFN-γ-dependent IL-10 reactivation. J. Immunol., 176, 7263-7271 (2006)[PubMed]
- Jankovic, D., Kullberg, M. C., Feng, C. G. et al.: Conventional T-bet+Foxp3- Th1 cells are the major source of host-protective regulatory IL-10 during intracellular protozoan infection. J. Exp. Med., 204, 273-283 (2007)[PubMed]
- Anderson, C. F., Oukka, M., Kuchroo, V. J. et al.: CD4+CD25-Foxp3- Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J. Exp. Med., 204, 285-297 (2007)[PubMed]
- Batten, M., Kljavin, N. M., Li, J. et al.: Cutting edge: IL-27 is a potent inducer of IL-10 but not FoxP3 in murine T cells. J. Immunol., 180, 2752-2756 (2008)[PubMed]
- Stumhofer, J. S., Silver, J. S., Laurence, A. et al.: Interleukins 27 and 6 induce STAT3-mediated T cell production of interleukin 10. Nat. Immunol., 8, 1363-1371 (2007)[PubMed]
- Hayashi, N., Yoshimoto, T., Izuhara, K. et al.: T helper 1 cells stimulated with ovalbumin and IL-18 induce airway hyperresponsiveness and lung fibrosis by IFN-γand IL-13 production. Proc. Natl. Acad. Sci. USA, 104, 14765-14770 (2007)[PubMed]
- Doi, M., Okano, T., Yujnovsky, I. et al.: Negative control of circadian clock regulator E4BP4 by casein kinase Iε-mediated phosphorylation. Curr. Biol., 14, 975-980 (2004)[PubMed]
- Zhang, W., Zhang, J., Kornuc, M. et al.: Molecular cloning and characterization of NF-IL3A, a transcriptional activator of the human interleukin-3 promoter. Mol.Cell. Biol., 15, 6055-6063 (1995)[PubMed]
- Ikushima, S., Inukai, T., Inaba, T. et al.: Pivotal role for the NFIL3/E4BP4 transcription factor in interleukin 3-mediated survival of pro-B lymphocytes. Proc. Natl. Acad. Sci. USA, 94, 2609-2614 (1997)[PubMed]
- Lund, R., Aittokallio, T., Nevalainen, O. et al.: Identification of novel genes regulated by IL-12, IL-4, or TGF-β during the early polarization of CD4+ lymphocytes. J. Immunol., 171, 5328-5336 (2003)[PubMed]
- Lund, R., Ahlfors, H., Kainonen, E. et al.: Identification of genes involved in the initiation of human Th1 or Th2 cell commitment. Eur. J. Immunol., 35, 3307-3319 (2005)[PubMed]
- Gascoyne, D. M., Long, E., Veiga-Fernandes, H. et al.: The basic leucine zipper transcription factor E4BP4 is essential for natural killer cell development. Nat. Immunol., 10, 1118-1124 (2009)[PubMed]
- Kamizono, S., Duncan, G. S., Seidel, M. G. et al.: Nfil3/E4bp4 is required for the development and maturation of NK cells in vivo. J. Exp. Med., 206, 2977-2986 (2009)[PubMed]
- Terashima, A., Watarai, H., Inoue, S. et al.: A novel subset of mouse NKT cells bearing the IL-17 receptor B responds to IL-25 and contributes to airway hyperreactivity. J. Exp. Med., 205, 2727-2733 (2008)[PubMed]
- Kuhn, R., Lohler, J., Rennick, D., Rajewsky, K. et al.: Interleukin-10-deficient mice develop chronic enterocolitis. Cell, 75, 263-274 (1993)[PubMed]
- Sun, J., Dodd, H., Moser, E. K. et al.: CD4+ T cell help and innate-derived IL-27 induce Blimp-1-dependent IL-10 production by antiviral CTLs. Nat. Immunol., 12, 327-334 (2011)[PubMed]
- Cretney, E., Xin, A., Shi, W. et al.: The transcription factors Blimp-1 and IRF4 jointly control the differentiation and function of effector regulatory T cells. Nat. Immunol., 12, 304-311 (2011)[PubMed]
- Okamura, T., Fujio, K., Shibuya, M. et al.: CD4+CD25-LAG3+ regulatory T cells controlled by the transcription factor Egr-2. Proc. Natl. Acad. Sci. USA, 106, 13974-13979 (2009)[PubMed]
著者プロフィール
略歴:2010年 東京医科歯科大学大学院生命情報科学教育部博士課程(理学)修了,同年より東京理科大学生命科学研究所 ポストドクトラル研究員.
研究テーマ:T細胞におけるサイトカイン遺伝子の制御機構の解明.
久保 允人(Masato Kubo)
東京理科大学生命科学研究所 教授,理化学研究所免疫アレルギー科学総合研究センター チームリーダー.
研究室URL:http://www.rs.noda.tus.ac.jp/~ribsjm/kubolab/
© 2011 本村泰隆・久保允人 Licensed under CC 表示 2.1 日本
