分泌タンパク質Rspoファミリーの新たな分子機構とその役割
大河原 美静
(名古屋大学大学院医学系研究科 神経遺伝情報学)
email:大河原美静
DOI: 10.7875/first.author.2011.061
Rspo3 binds syndecan 4 and induces Wnt/PCP signaling via clathrin-mediated endocytosis to promote morphogenesis.
Bisei Ohkawara, Andrei Glinka, Christof Niehrs
Developmental Cell, 20, 303-314 (2011)
分泌タンパク質RspoファミリーはWnt分泌タンパク質と協調してWntシグナル伝達経路を活性化し,発生過程の多くの現象や疾患の発症に関与していることが知られている.しかし,その受容体が明確でなくどのようにWntシグナル伝達経路を制御しているのかは不明であった.この論文では,Rspo3がSdc4と結合しWnt/PCPシグナル伝達経路ではたらくキナーゼJNKを活性化していることを明らかにした.アフリカツメガエルの初期胚ではWnt/PCPシグナル伝達経路を必要とする原腸陥入と頭部の軟骨形成においてRspo3とSdc4が必須であることも示した.さらに,Rspo3およびSdc4はWnt5aとその受容体であるFzd7とDvlを介してJNKを活性化し,この活性化にはRspo3が誘導するクラスリンを介したエンドサイトーシスが必要であることも証明した.これらのことから,Rspo3はSdc4を介してWnt5a-Fzd7複合体のエンドサイトーシスを促進することによりWnt/PCPシグナル伝達経路を制御していることが示唆された.
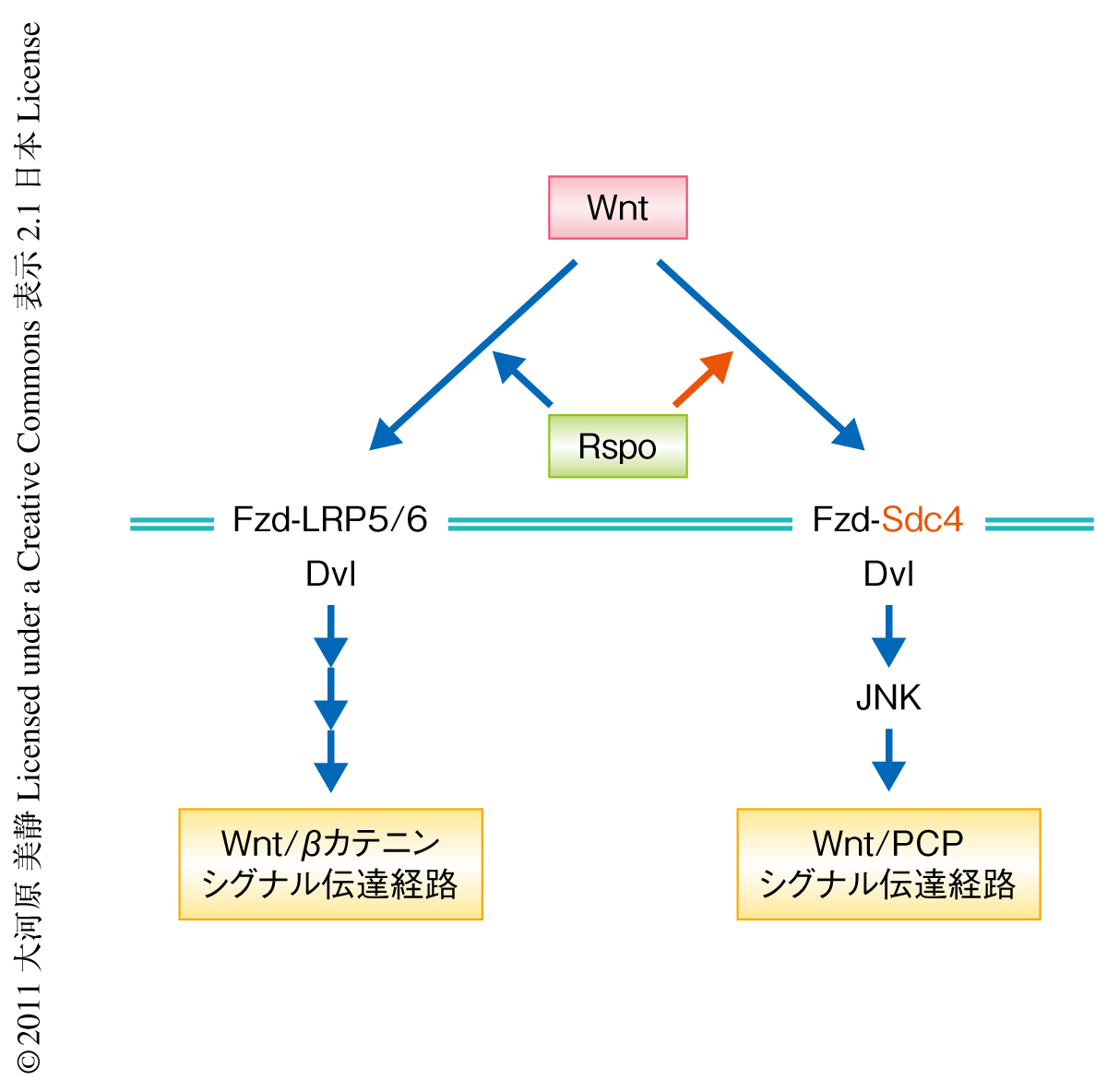
Wnt分泌タンパク質はさまざまな生命現象に関与するシグナル分泌タンパク質で,転写のコアクチベーターであるβカテニンにより仲介されるWnt/βカテニンシグナル伝達経路と,それに依存しないWnt/PCPシグナル伝達経路という2つのシグナル伝達経路を活性化する(図1).R-spondin(Rspo)はWntと同様に細胞外に放出されるシグナル分泌タンパク質であり,Wntシグナル伝達経路によりその発現が誘導されこのシグナル伝達経路を増強および維持する正のフィードバック制御タンパク質である1).Wntがその受容体であるFrizzled(Fzd)やLRP5あるいはLRP6に結合し細胞におけるシグナル伝達タンパク質であるDshevelled(Dvl)やAxinの局在を変えることでシグナル伝達経路を活性化しているのに対し2),Rspoファミリーはその受容体が明確にされておらず,細胞外の情報を細胞内に伝える分子機構もわかっていなかった.また,WntがWnt/PCPシグナル伝達経路をも活性化することがわかっている一方で,Wntと同時に機能するRspoファミリーがWnt/PCPシグナル伝達経路を活性化するかどうかということも検討されていなかった.
膜貫通型タンパク質Syndecan(Sdc)はその細胞外ドメインがヘパラン硫酸という糖鎖により修飾されているプロテオグリカンで,Wntを含むさまざまな分泌タンパク質と細胞膜の表面で結合することによりそれら分泌タンパク質の誘導するシグナル伝達経路の活性を制御する補助受容体として機能している3).アフリカツメガエルの初期胚ではSdc4がWnt/PCPシグナル伝達経路に必須であるという報告があった4).
筆者らは,アフリカツメガエルの初期胚においてRspo3をノックダウンするとさまざまな部位で形態形成の不全がみられ,このことはRspo3がWnt/βカテニンシグナル伝達経路のみを制御していることでは説明のつかないこと,Rspo3ノックダウンによる表現型の一部がSdc4やWnt5aのノックダウンによる表現型と類似していること,Rspoファミリーにはプロテオグリカンに強い親和性をもつトロンボスポンジンI型ドメインがありその活性に大きく影響していることから,Rspo3とSdc4の役割について検討した.
Rspo3の受容体の候補としてWntシグナル伝達経路に関与する膜タンパク質が考えられた.その探索のためRspo3とアルカリホスファターゼとの融合タンパク質を用いて細胞膜における結合実験を行った.その結果,Rspo3はほかの膜タンパク質に比べプロテオグリカンであるSdc4やGly4に強く結合することがわかった.また,それぞれのSdcファミリーとの結合の強さはRspoファミリーのそれぞれの活性の強さと相関があり5),Rspoファミリーはプロテオグリカンのヘパラン硫酸糖鎖に結合していること,Sdc4とRspo3との結合定数は生物学的に有意であることが示された.
Sdc4はアフリカツメガエルの初期胚においてWnt/PCPシグナル伝達経路を活性化し原腸陥入のときの細胞移動に必須である4).Rspo3はWntに依存してWnt/βカテニンシグナル伝達経路を活性化することから1),Wntの関与する別のシグナル伝達経路にも機能することを予想してRspo3のWnt/PCPシグナル伝達経路に対する役割を検討した.その結果,Rspo3はアフリカツメガエルの初期胚でWnt/PCPシグナル伝達経路を制御しているWnt5aやSdc4とともに原腸陥入運動に関与していること,細胞レベルでの観察より細胞が移動する際に特徴的に表われる突出部(protrusion)の形成に必須であること,Wnt/PCPシグナル伝達経路で機能するキナーゼJNKのリン酸化を上昇させること,がわかった.
実験を進めていく過程で,Rspo3ノックダウンによる表現型はノックダウンする部位により異なっていることがわかった6).このことから,Rspo3遺伝子の発現が発生過程においていろいろな組織あるいは部位でオン/オフをくり返すことでさまざまな現象に関与していると考え,Rspo3ノックダウンによる表現型とRspo3の発現部位とを詳細に比べた.その結果,新たにRspo3が頭部軟骨細胞に発現し頭部骨格の形成に関与していることを見い出した.この表現型はアフリカツメガエルの胚では深く検討されていなかったが,ゼブラフィッシュの胚におけるWnt5aあるいはプロテオグリカンであるGly4のノックダウン表現型7,8) と類似していた.そこで,アフリカツメガエルの胚で頭部軟骨組織を観察したところコンドロサイト細胞の並び方に異常が見られた.同様の異常がWnt5aノックダウン個体においてもみられたことから,Rspo3が頭部骨格の形成においてもWnt5aと同様に機能していることが証明された.
Rspo3の分子機構を探る際,これまではWnt/βカテニンシグナル伝達経路のみが注目されていた.今回,新たにWnt/PCPシグナル伝達経路への関与が示されたことから,Wnt/βカテニンシグナル伝達経路とWnt/PCPシグナル伝達経路の両方に共通するタンパク質がRspo3の機能に関与することが予想された.そこで,Rspo3がWnt/PCPシグナル伝達経路を活性化する際にどのタンパク質に依存しているかを検討した.Wnt/PCPシグナル伝達経路ではRspo3結合タンパク質であるSdc4,リガンドであるWnt5a,その受容体であるFzd7,Fzd7の細胞内ドメインに結合しシグナルを伝達するDvlが,Rspo3の機能に必須であることがわかった.また,Wnt/βカテニンシグナル伝達経路に必須であるLRP6はRspo3によるWnt/PCPシグナル伝達経路の誘導には関与しないことがわかった.これらのことより,Rspo3は細胞外でSdc4を介してWnt5a-Fzd7複合体になんらかのはたらきかけをしているものと示唆された.
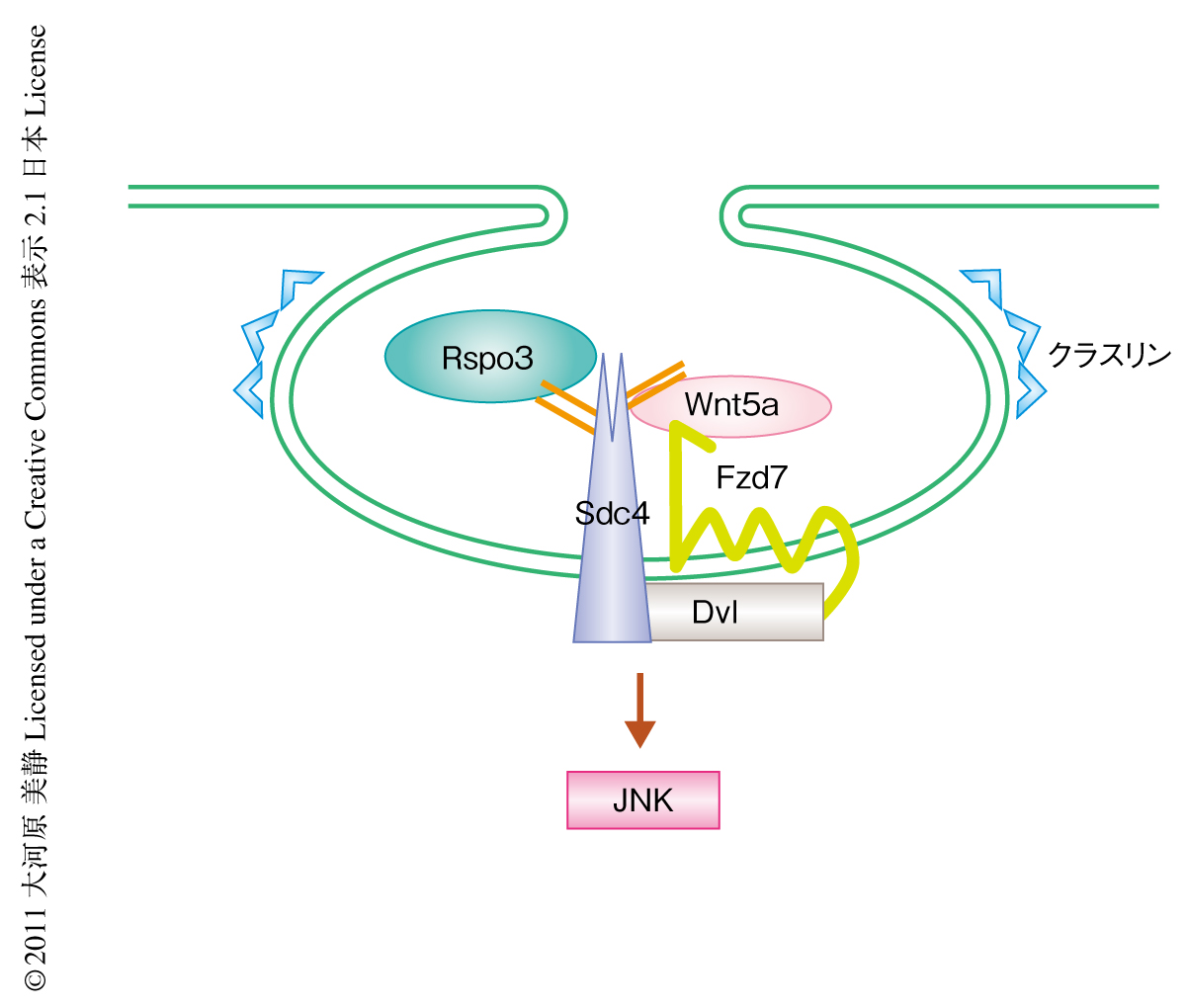
Sdc4はシグナル分泌タンパク質であるFGFの補助受容体としてはたらくときFGFを細胞に取り込むこと(エンドサイトーシス)が知られている3).Wntの存在によりその受容体が細胞に取り込まれるようすがみられていること9,10),エンドサイトーシスに機能するクラスリンやカベオリンがWntシグナル伝達経路に必須であることから10),現在では,エンドサイトーシスによりWntとその受容体複合体が細胞に取り込まれることがWntシグナル伝達経路の活性化に必須であると考えられている.そこで,Rspo3がエンドサイトーシスを介し下流のタンパク質を活性化しているのかどうか検討した.その結果,Rspo3はSdc4,Wnt5aやFzd7に依存して細胞に取り込まれること,それによりFzd7と結合するDvlの局在の変化することがわかった.さらに,このエンドサイトーシスはクラスリンに依存的であり,Wntシグナル伝達経路の下流タンパク質であるJNKのリン酸化に必要であることもわかった.
これらの結果から,Rspo3はWnt/βカテニン伝達経路だけでなくWnt/PCPシグナル伝達経路も活性化すること,そのとき,Sdc4に結合しリガンド-受容体複合体であるWnt5a-Fzd7複合体と協同で機能すること,また,Wntシグナル伝達経路の活性化にはWnt5a-Fzd7複合体のエンドサイトーシスが必要であることがわかった(図2).この知見は,この論文では注目しなかったWnt/βカテニンシグナル伝達経路におけるRspoファミリーの役割を調べるうえでも参考になるものと考えている.Wntシグナル伝達経路はがんなどさまざまな疾患に関与しているが,その経路で機能するタンパク質の多くは生体のさまざまな場面で機能をもつため,これらを標的とした薬剤の開発は困難である.Rspoファミリーはその機能がWntシグナル伝達経路に特異的であること,がん細胞において高い発現がみられること,分泌タンパク質であるなど,創薬において多くの利点のあることから薬剤の標的としても注目される.
略歴:2002年 総合研究大学院大学生命科学研究科 修了,同年 東京医科歯科大学難治疾患研究所 研究員,2005年 ドイツGerman Cancer Research Center(DKFZ)研究員を経て,2010年より名古屋大学大学院医学系研究科 グローバルCOE特任助教.
研究テーマ:細胞間シグナル伝達,Wntシグナル伝達経路.
関心事:生体内においてとなりあった細胞が分泌タンパク質を介して何をひそひそおしゃべりしているのか聞いてみたい.
© 2011 大河原 美静 Licensed under CC 表示 2.1 日本
(名古屋大学大学院医学系研究科 神経遺伝情報学)
email:大河原美静
DOI: 10.7875/first.author.2011.061
Rspo3 binds syndecan 4 and induces Wnt/PCP signaling via clathrin-mediated endocytosis to promote morphogenesis.
Bisei Ohkawara, Andrei Glinka, Christof Niehrs
Developmental Cell, 20, 303-314 (2011)
要 約
分泌タンパク質RspoファミリーはWnt分泌タンパク質と協調してWntシグナル伝達経路を活性化し,発生過程の多くの現象や疾患の発症に関与していることが知られている.しかし,その受容体が明確でなくどのようにWntシグナル伝達経路を制御しているのかは不明であった.この論文では,Rspo3がSdc4と結合しWnt/PCPシグナル伝達経路ではたらくキナーゼJNKを活性化していることを明らかにした.アフリカツメガエルの初期胚ではWnt/PCPシグナル伝達経路を必要とする原腸陥入と頭部の軟骨形成においてRspo3とSdc4が必須であることも示した.さらに,Rspo3およびSdc4はWnt5aとその受容体であるFzd7とDvlを介してJNKを活性化し,この活性化にはRspo3が誘導するクラスリンを介したエンドサイトーシスが必要であることも証明した.これらのことから,Rspo3はSdc4を介してWnt5a-Fzd7複合体のエンドサイトーシスを促進することによりWnt/PCPシグナル伝達経路を制御していることが示唆された.
はじめに
Wnt分泌タンパク質はさまざまな生命現象に関与するシグナル分泌タンパク質で,転写のコアクチベーターであるβカテニンにより仲介されるWnt/βカテニンシグナル伝達経路と,それに依存しないWnt/PCPシグナル伝達経路という2つのシグナル伝達経路を活性化する(図1).R-spondin(Rspo)はWntと同様に細胞外に放出されるシグナル分泌タンパク質であり,Wntシグナル伝達経路によりその発現が誘導されこのシグナル伝達経路を増強および維持する正のフィードバック制御タンパク質である1).Wntがその受容体であるFrizzled(Fzd)やLRP5あるいはLRP6に結合し細胞におけるシグナル伝達タンパク質であるDshevelled(Dvl)やAxinの局在を変えることでシグナル伝達経路を活性化しているのに対し2),Rspoファミリーはその受容体が明確にされておらず,細胞外の情報を細胞内に伝える分子機構もわかっていなかった.また,WntがWnt/PCPシグナル伝達経路をも活性化することがわかっている一方で,Wntと同時に機能するRspoファミリーがWnt/PCPシグナル伝達経路を活性化するかどうかということも検討されていなかった.
膜貫通型タンパク質Syndecan(Sdc)はその細胞外ドメインがヘパラン硫酸という糖鎖により修飾されているプロテオグリカンで,Wntを含むさまざまな分泌タンパク質と細胞膜の表面で結合することによりそれら分泌タンパク質の誘導するシグナル伝達経路の活性を制御する補助受容体として機能している3).アフリカツメガエルの初期胚ではSdc4がWnt/PCPシグナル伝達経路に必須であるという報告があった4).
筆者らは,アフリカツメガエルの初期胚においてRspo3をノックダウンするとさまざまな部位で形態形成の不全がみられ,このことはRspo3がWnt/βカテニンシグナル伝達経路のみを制御していることでは説明のつかないこと,Rspo3ノックダウンによる表現型の一部がSdc4やWnt5aのノックダウンによる表現型と類似していること,Rspoファミリーにはプロテオグリカンに強い親和性をもつトロンボスポンジンI型ドメインがありその活性に大きく影響していることから,Rspo3とSdc4の役割について検討した.
1.Rspo3はSdc4と結合する
Rspo3の受容体の候補としてWntシグナル伝達経路に関与する膜タンパク質が考えられた.その探索のためRspo3とアルカリホスファターゼとの融合タンパク質を用いて細胞膜における結合実験を行った.その結果,Rspo3はほかの膜タンパク質に比べプロテオグリカンであるSdc4やGly4に強く結合することがわかった.また,それぞれのSdcファミリーとの結合の強さはRspoファミリーのそれぞれの活性の強さと相関があり5),Rspoファミリーはプロテオグリカンのヘパラン硫酸糖鎖に結合していること,Sdc4とRspo3との結合定数は生物学的に有意であることが示された.
2.Sdc4と同様にRspo3も原腸陥入のときの細胞移動に関与する
Sdc4はアフリカツメガエルの初期胚においてWnt/PCPシグナル伝達経路を活性化し原腸陥入のときの細胞移動に必須である4).Rspo3はWntに依存してWnt/βカテニンシグナル伝達経路を活性化することから1),Wntの関与する別のシグナル伝達経路にも機能することを予想してRspo3のWnt/PCPシグナル伝達経路に対する役割を検討した.その結果,Rspo3はアフリカツメガエルの初期胚でWnt/PCPシグナル伝達経路を制御しているWnt5aやSdc4とともに原腸陥入運動に関与していること,細胞レベルでの観察より細胞が移動する際に特徴的に表われる突出部(protrusion)の形成に必須であること,Wnt/PCPシグナル伝達経路で機能するキナーゼJNKのリン酸化を上昇させること,がわかった.
3.Rspo3は頭部骨格の形成に関与する
実験を進めていく過程で,Rspo3ノックダウンによる表現型はノックダウンする部位により異なっていることがわかった6).このことから,Rspo3遺伝子の発現が発生過程においていろいろな組織あるいは部位でオン/オフをくり返すことでさまざまな現象に関与していると考え,Rspo3ノックダウンによる表現型とRspo3の発現部位とを詳細に比べた.その結果,新たにRspo3が頭部軟骨細胞に発現し頭部骨格の形成に関与していることを見い出した.この表現型はアフリカツメガエルの胚では深く検討されていなかったが,ゼブラフィッシュの胚におけるWnt5aあるいはプロテオグリカンであるGly4のノックダウン表現型7,8) と類似していた.そこで,アフリカツメガエルの胚で頭部軟骨組織を観察したところコンドロサイト細胞の並び方に異常が見られた.同様の異常がWnt5aノックダウン個体においてもみられたことから,Rspo3が頭部骨格の形成においてもWnt5aと同様に機能していることが証明された.
4.Rspo3はSdc4,Wnt5a,Fzd7を介してWnt/PCPシグナル伝達経路を活性化する
Rspo3の分子機構を探る際,これまではWnt/βカテニンシグナル伝達経路のみが注目されていた.今回,新たにWnt/PCPシグナル伝達経路への関与が示されたことから,Wnt/βカテニンシグナル伝達経路とWnt/PCPシグナル伝達経路の両方に共通するタンパク質がRspo3の機能に関与することが予想された.そこで,Rspo3がWnt/PCPシグナル伝達経路を活性化する際にどのタンパク質に依存しているかを検討した.Wnt/PCPシグナル伝達経路ではRspo3結合タンパク質であるSdc4,リガンドであるWnt5a,その受容体であるFzd7,Fzd7の細胞内ドメインに結合しシグナルを伝達するDvlが,Rspo3の機能に必須であることがわかった.また,Wnt/βカテニンシグナル伝達経路に必須であるLRP6はRspo3によるWnt/PCPシグナル伝達経路の誘導には関与しないことがわかった.これらのことより,Rspo3は細胞外でSdc4を介してWnt5a-Fzd7複合体になんらかのはたらきかけをしているものと示唆された.
5.Rspo3はエンドサイトーシスされる
Sdc4はシグナル分泌タンパク質であるFGFの補助受容体としてはたらくときFGFを細胞に取り込むこと(エンドサイトーシス)が知られている3).Wntの存在によりその受容体が細胞に取り込まれるようすがみられていること9,10),エンドサイトーシスに機能するクラスリンやカベオリンがWntシグナル伝達経路に必須であることから10),現在では,エンドサイトーシスによりWntとその受容体複合体が細胞に取り込まれることがWntシグナル伝達経路の活性化に必須であると考えられている.そこで,Rspo3がエンドサイトーシスを介し下流のタンパク質を活性化しているのかどうか検討した.その結果,Rspo3はSdc4,Wnt5aやFzd7に依存して細胞に取り込まれること,それによりFzd7と結合するDvlの局在の変化することがわかった.さらに,このエンドサイトーシスはクラスリンに依存的であり,Wntシグナル伝達経路の下流タンパク質であるJNKのリン酸化に必要であることもわかった.
おわりに
これらの結果から,Rspo3はWnt/βカテニン伝達経路だけでなくWnt/PCPシグナル伝達経路も活性化すること,そのとき,Sdc4に結合しリガンド-受容体複合体であるWnt5a-Fzd7複合体と協同で機能すること,また,Wntシグナル伝達経路の活性化にはWnt5a-Fzd7複合体のエンドサイトーシスが必要であることがわかった(図2).この知見は,この論文では注目しなかったWnt/βカテニンシグナル伝達経路におけるRspoファミリーの役割を調べるうえでも参考になるものと考えている.Wntシグナル伝達経路はがんなどさまざまな疾患に関与しているが,その経路で機能するタンパク質の多くは生体のさまざまな場面で機能をもつため,これらを標的とした薬剤の開発は困難である.Rspoファミリーはその機能がWntシグナル伝達経路に特異的であること,がん細胞において高い発現がみられること,分泌タンパク質であるなど,創薬において多くの利点のあることから薬剤の標的としても注目される.
文 献
- Kazanskaya, O., Glinka, A., del Barco Barrantes, I. et al.: R-Spondin2 is a secreted activator of Wnt/β-catenin signaling and is required for Xenopus myogenesis. Dev. Cell, 4, 525-34 (2004)[PubMed]
- Bilic, J., Huang, Y. L., Davidson, G. et al.: Wnt induces LRP6 signalosomes and promotes disheveled-dependent LRP6 phosphorylation. Science, 316, 1619-1622 (2007)[PubMed]
- Tkachenko, E., Rhodes, J. M. & Simons, M.: Syndecans: new kids on the signaling block. Circ. Res., 96, 488-500 (2005)[PubMed]
- Munoz, R., Moreno, M., Oliva, C. et al.: Syndecan-4 regulates non-canonical Wnt signalling and is essential for convergent and extension movements in Xenopus embryos. Nat. Cell Biol., 5, 492-500 (2006)[PubMed]
- Kim, K. A., Wagle, M., Tran, K. et al.: R-spondin family members regulate the Wnt pathway by a common mechanism. Mol. Biol. Cell, 6, 2588-2596 (2008)[PubMed]
- Kazanskaya, O., Ohkawara, B., Heroult, M. et al.: The Wnt signaling regulator R-spondin 3 promotes angioblast and vascular development. Development, 22, 3655-3564 (2008)[PubMed]
- Piotrowski, T., Schilling, T. F., Brand, M. et al.: Jaw and branchial arch mutants in zebrafish II: anterior arches and cartilage differentiation. Development, 123, 345-356 (1996)[PubMed]
- Topczewski, J., Sepich, D. S., Myers, D. C. et al.: The zebrafish glypican knypek controls cell polarity during gastrulation movements of convergent extension. Dev. Cell, 2, 251-264 (2001)[PubMed]
- Cruciat, C. M., Ohkawara, B., Acebron, S. P. et al.: Requirement of prorenin receptor and vacuolar H+-ATPase-mediated acidification for Wnt signaling. Science, 327, 459-463 (2010)[PubMed]
- Kikuchi, A., Yamamoto, H. & Sato, A.: Selective activation mechanisms of Wnt signaling pathways. Trends Cell Biol., 3, 119-129 (2009)[PubMed]
著者プロフィール
略歴:2002年 総合研究大学院大学生命科学研究科 修了,同年 東京医科歯科大学難治疾患研究所 研究員,2005年 ドイツGerman Cancer Research Center(DKFZ)研究員を経て,2010年より名古屋大学大学院医学系研究科 グローバルCOE特任助教.
研究テーマ:細胞間シグナル伝達,Wntシグナル伝達経路.
関心事:生体内においてとなりあった細胞が分泌タンパク質を介して何をひそひそおしゃべりしているのか聞いてみたい.
© 2011 大河原 美静 Licensed under CC 表示 2.1 日本
