脳の固有のシナプス層への視神経の軸索の投射において2つの膜タンパク質が協調している
鈴木(羽毛田)聡子・鈴木崇之
(ドイツMax Planck Institute of Neurobiology,Axonal Guidance and Neuronal Connectivity)
email:鈴木崇之
DOI: 10.7875/first.author.2011.033
Golden Goal collaborates with Flamingo in conferring synaptic-layer specificity in the visual system.
Satoko Hakeda-Suzuki, Sandra Berger-Müller, Tatiana Tomasi, Tadao Usui, Shin-ya Horiuchi, Tadashi Uemura, Takashi Suzuki
Nature Neuroscience, 14, 314-323 (2011)
脳神経組織において相同の機能をもつニューロンは決まった1層においてシナプスを形成し,それが集まって何重もの層状の構造を形成することが多い.ショウジョウバエのR7視細胞とR8視細胞は空間の同一な点からの異なった波長の光を感知し,脳(視覚中枢)の異なったシナプス層に軸索を伸ばしつなぎとめている.ニューロンの軸索がどのように固有のシナプス層を認識し選択をするのか,その分子機構はよくわかっていない.筆者らは,視神経の軸索の投射を制御する2つの細胞表面タンパク質,GogoとFmiがR8視細胞の軸索において機能的に相互作用することを見い出した.Gogo単独のはたらきによりR8視細胞の軸索を一過的にM1層にとどめ,そののち,GogoとFmiとが協調してR8視細胞の軸索を最終的な標的であるM3層へと導いていた.構造機能解析の結果,GogoはFmiと協調する際に細胞内シグナルを伝達する役割を担っていることが示唆された.さらに,R8視細胞の軸索の投射においてFmiは脳の一部のニューロンで必要とされていることがわかり,このことは,R8視細胞のFmiと脳の標的細胞のFmiとがホモ接触結合することによりR8視細胞のM3層への投射の起こることを示唆していた.この研究によって,Gogoは神経軸索の投射においてFmiの機能的なパートナーであり,これら2つのタンパク質の相互作用をダイナミックに制御することにより視神経の軸索のシナプス層の特異的な選択が正確に行われていることが示唆された.
ニューロンどうしの正確なシナプスの形成は脳における情報演算処理にとり非常に重要である.シナプスはしばしば機能的な意味あいを反映する構造を形成する.たとえば,視覚系ではとなりあった視細胞は脳の標的領域においてもとなりあった柱状構造に軸索を投射しシナプスを接続する.この特徴的な構造により外の視覚世界からの視覚信号と脳に映し出される映像信号との位置関係を維持している1).この柱状の網膜脳相関維持(retinotopy)構造にくわえ層状の構造もとっており,外界からの映像を異なった成分に分解しそれぞれの成分を別々のシナプス層で情報処理することも行われている.これらのような特徴的な柱状かつ層状の構造は複雑な神経組織において広くみられるものである2).それでは,このシナプスの層状構造はどのような分子機構により形成されているのだろうか.ニューロンの軸索はどのように固有のシナプス層を認識しそこに投射するのか,その分子機構はよくわかっていない.脊椎動物の視覚系では,SidekicksやDscamsなど,いくつかのホモ結合接着分子がそれどうしの接着の選択性を利用して視細胞におけるシナプス層の特異性を決めていることが知られている1).
ショウジョウバエの視覚系も層状の構造をとっており,シナプスの結合に特異性のあることもよく知られている.ショウジョウバエの複眼は約800個の個眼からできており,1つの個眼には8個の視細胞(R1~R8)がある.R1~R6の視細胞は物体の動きを感知し,軸索は最初の視神経節であるラミナに投射する.色覚をつかさどるR7視細胞とR8視細胞は2番目の視神経節であるメダラに軸索を投射する.1つの個眼においてR7視細胞とR8視細胞のペアは空間の同一の点から色信号を受け取り,メダラの同一の柱状の構造に軸索を投射する.興味深いことに,このR7視細胞とR8視細胞とはそれぞれ別々の波長の色信号を感知しているのだが,軸索を別々のメダラ層(R8視細胞はM3層,R7視細胞はM6層に)に投射してシナプスを形成する(図1).このようにして,個々の機能モジュールは別々のシナプス層でプロセスされている.このようなことから,ショウジョウバエの視覚系はシナプス層の特異的な選択機構を研究する格好のモデル系であるといえる1,3).

ショウジョウバエではいくつかの膜貫通型タンパク質がシナプス層に特異的な投射を制御していることが知られている4).そのような例として,カドヘリンの一種であるN-カドヘリンとFlamingo 5-7)(Fmi),受容体型チロシンキナーゼであるLARとPtp69D,接着分子であるCapricious,受容体と予想されるGolden Goal 8)(Gogo)などが知られている.それぞれ個々の機能的な役割は報告されているが,これらのタンパク質が総合的にどのようにかかわりあってシナプス層の特異性を決定しているのかという全体像はよくわかっていない.
筆者らは,R8視細胞に着目して研究を進めた9).このR8視細胞ではFmiが非常に重要な役割を担っていることがわかった.Fmiは,カドヘリンがそうであるように,試験管内では強くホモ結合接着(同じ分子種どうしが接着する)することが知られている5).しかし,生体ではFmiはすべての種類の視細胞に発現しており,しかも,脳の標的領域であるメダラ層においても何層にも広範に発現している.このことは,ほかのタンパク質がFmiの接着活性をタイミングよく制御してR8視細胞の投射を制御していることを示唆していた.筆者らは,Gogoの変異体がFmiの変異体と似たR8視細胞の表現型を示すことから,Fmiの機能的なパートナーはGogoではないかと考えそれを実証した.
筆者らがすでに発表した研究により,gogo変異体はR8視細胞の軸索の投射においてfmi変異体6,7) と似た表現型を示すことが知られていた8).それにくわえて,この研究では,gogo変異体がR1~R6の視細胞のラミナでのシナプスの選択においてfmi変異体と同様の間違いを起こすことを見い出した.さらに,gogo変異体は胚の末梢神経系の樹状突起においてもfmi変異体と同様に10),伸びすぎて背中線をこえるという表現型をみせた.GogoとFmiはタンパク質の発現や局在パターンでも相当に重なる部分の多いこともわかった6).これらのことは,この2つのタンパク質が協調的にはたらいていることを示唆していた.
GogoとFmiとが機能的に協調してはたらくことを示すため,遺伝学的な相互作用をみた.具体的には,Fmiの不完全な変異体(ハイポモルフ)について,それにくわえてGogoの遺伝子を半減したときに異常が強くなるかどうかをみた.その結果,R8視細胞の軸索の投射にくわえて,GogoとFmiとは成体になるまでの生存率や樹状突起網の形成において遺伝学的な相互作用があった.これは,GogoとFmiとの協調的なはたらきが視神経だけではなく広く神経系の全般において起こっていることを示唆していた.
また逆に,野生型ではGogoとFmiが機能していないR7視細胞にこの2つのタンパク質を強制的に共発現してみた.すると,本来はR7視細胞の軸索はM6層に投射するのだが,R8視細胞が投射するM3層に投射するようになることがわかった.このことから,GogoとFmiとを組み合わせることによりR7視細胞の軸索はR8視細胞の軸索の形質を獲得しそのようにふるまうことがわかった.GogoとFmiとはR8視細胞の軸索のシナプス層への投射に必要かつ十分であることが示された.
GogoとFmiとが物理的な相互作用をするかどうかの示唆を得るため,培養細胞においてGogoとFmiの局在を調べた.その結果,細胞凝集団において細胞のあいだの接触面にFmiが集積すると,Gogoも必ず集積することがわかった.この共集積はGogoの細胞外ドメインを介して起こることが,Gogoの細胞内ドメインの欠損変異体なども共集積することから示唆された.
では,生体でもGogoとFmiとは相互作用するのだろうか? 発生途中の翅の細胞を用いて検証した.野生型の翅の細胞ではFmiは発現し細胞の頂端側に特徴的に局在するのだが,Gogoは発現していない.この状況でGogoを強制発現させると,FmiはGogoにひき連れられて細胞の横面にも局在するようになった.GogoとFmiとがシスあるいはトランスに相互作用するのかを調べるため,Gogoの強制発現とfmi変異体の細胞クローンとをランダムに組み合わせ翅においてモザイク状とした.その細胞クローンの境界を調べた結果,このGogoによるFmiの局在のシフトはGogoとFmiとが同じ細胞に存在するときだけみられた.そこで,GogoとFmiとはシスに相互作用することが示唆された(図2a).
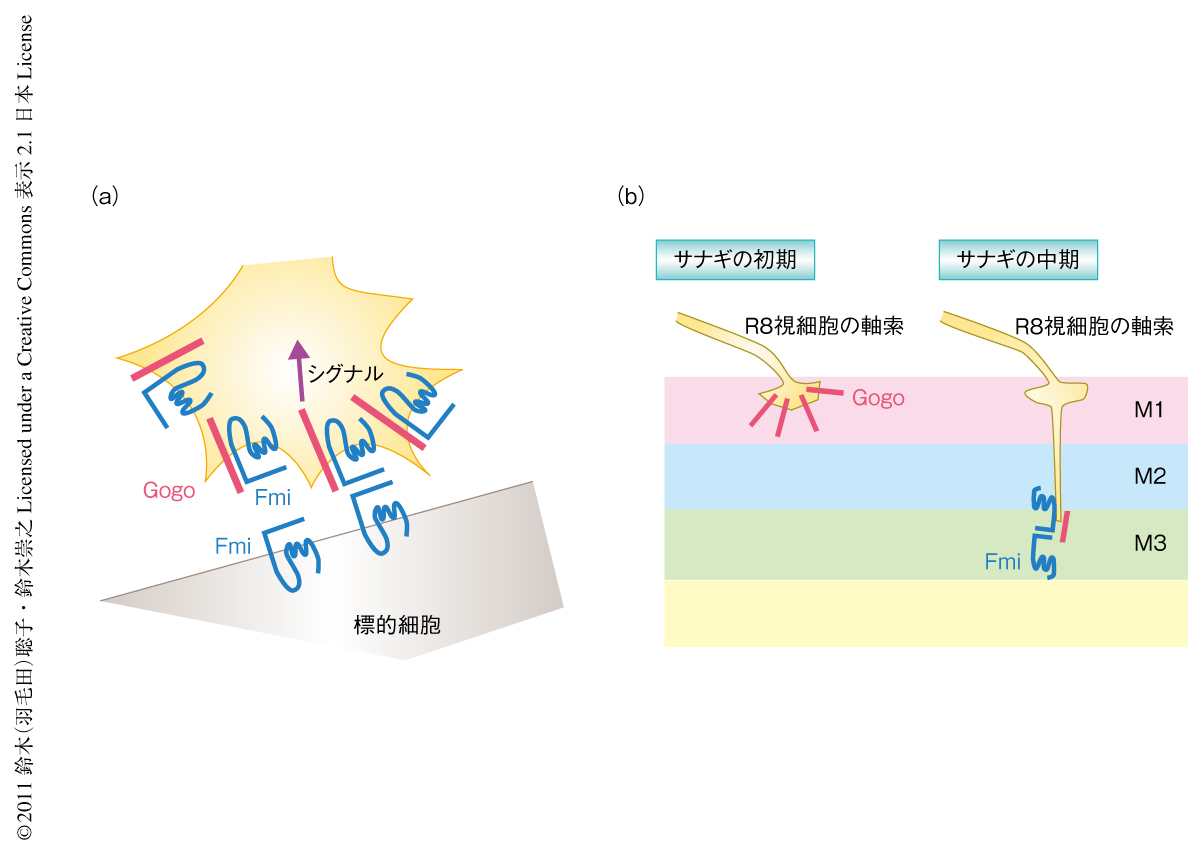
では,GogoはFmiと協調したとき,R8視細胞の軸索の投射においてどのような役割を担っているのだろうか? 以前の研究から,Gogoの細胞内ドメインを欠損させるとgogo変異体の表現型をレスキューできないことがわかっていた8).しかしながら,Fmiの場合はその細胞内ドメインを欠損させてもfmi変異体の表現型をレスキューできた.これらのことから,GogoはFmiとその細胞外ドメインで相互作用し,Gogoの細胞内ドメインを介して投射シグナルをR8視細胞の軸索に伝達しているのではないかと予想できた.これを検証するため,さきのR7視細胞でのGogoとFmiとの共強制発現の系を使い,生体におけるそれらの細胞内ドメインの影響力をみた.FmiとGogoの全長または細胞内ドメインを欠損させた遺伝子を組み合わせた結果,Gogoの細胞内ドメインがR7視細胞の投射先を(本来はR8視細胞の標的である)M3層に変更させるのに必要かつ十分であることがわかった.このことは,投射シグナルをR8視細胞の軸索に伝達することがFmiと協調したときのGogoの重要な役割のひとつであることを示唆していた(図2a).
脳のFmiがR8視細胞の軸索のリガンドになっているかどうかは不明であった.その原因として,脳にfmi変異体の大きな細胞集団クローンを作製するのがむずかしいことがあげられた.そこで,細胞のタイプに特異的なコンディショナルノックアウト細胞を作製するため新しい系を開発して脳にfmi変異体の大きな細胞集団クローンを作製した.脳のラミナニューロンからFmiを取り除くとR8視細胞の軸索の投射が相当な割合で異常となり,M1層でとまってしまうことがわかった.このことから,脳のFmiがR8視細胞の軸索の投射に必要であり,脳のFmiとR8視細胞のFmiとが相互作用をしていることが示唆された(図2a).
すでに報告されたGogoの過剰発現では,R8視細胞はM1層に強く接着する表現型を示すことが知られていた8).今回の研究では,Gogoは(Fmiと協調して)M3層に投射するのを助けていた.この2つの実験結果は矛盾せずに説明できるのだろうか? Gogoの過剰発現ではR8視細胞の軸索の途中(M1層)に大きな軸索塊ができた.この状況でFmiを不完全な変異体にすると軸索塊がより大きくなり,Fmiを強制発現すると軸索塊の形成が抑制された.このことから,FmiはM1層の付近ではGogoと拮抗するようにはたらくことがわかった.
以上のことをまとめると,サナギの初期ではGogo単独でM1層に接着し,サナギの中期にR8視細胞がM3層に軸索を伸ばしはじめるときにGogoとFmiとが協調してR8視細胞をM3層に誘導するものと考えられた(図2b).ちなみに,M3層におけるリガンドはFmiだと推察しているが,M1層におけるリガンドは現時点では不明である.
この研究によって,細胞の表面にあるタンパク質の活動あるいは活性の組合せを細胞特異的かつ経時的に変化させることによりニューロンのシナプス層に特異的な投射の制御されていることが示唆された.この分子機構は,視神経にかぎらずあらゆる脳神経組織の発生段階においてあてはまる普遍原理的なものである可能性がある.これを理解することは,われわれの五感・意識・思索・行動などのみなもとである脳神経組織の形成の分子機序を理解するうえで非常に重要である.
また,近年,幹細胞の移植などにより神経再生医療の道が開けようとしている.神経系の機能的な回復には,いうまでもなく,神経回路網の再構築が必要である.この研究のように,神経回路網の形成の分子機構を理解することによって,将来は神経再生医療にも役立つ可能性があると考えている.
略歴:2000年 東京大学大学院理学系研究科 修了,同年 オーストリアResearch Institute of Molecular Pathology博士研究員を経て,2005年よりドイツMax Planck Institute of Neurobiology博士研究員.
研究テーマ:神経回路の形成,軸索の投射,樹状突起網の形成,アクチン骨格系.
鈴木 崇之(Takashi Suzuki)
略歴:2000年 東京大学大学院理学系研究科 修了,同年 オーストリアResearch Institute of Molecular Pathology博士研究員を経て,2005年よりドイツMax Planck Institute of Neurobiologyグループリーダー.
研究テーマ:神経回路の形成,シナプス形成の決定機構.
抱負:ショウジョウバエには生まれながらに高度な神経活動を遂行できるような神経回路網が備わっている.ヒトと違って神経活動に非依存的なのは驚くべきことだ.その形成機構にせまりたい.
研究室URL:http://www.neuro.mpg.de/english/junior/axguide/index.html
© 2011 鈴木(羽毛田)聡子・鈴木崇之 Licensed under CC 表示 2.1 日本
(ドイツMax Planck Institute of Neurobiology,Axonal Guidance and Neuronal Connectivity)
email:鈴木崇之
DOI: 10.7875/first.author.2011.033
Golden Goal collaborates with Flamingo in conferring synaptic-layer specificity in the visual system.
Satoko Hakeda-Suzuki, Sandra Berger-Müller, Tatiana Tomasi, Tadao Usui, Shin-ya Horiuchi, Tadashi Uemura, Takashi Suzuki
Nature Neuroscience, 14, 314-323 (2011)
要 約
脳神経組織において相同の機能をもつニューロンは決まった1層においてシナプスを形成し,それが集まって何重もの層状の構造を形成することが多い.ショウジョウバエのR7視細胞とR8視細胞は空間の同一な点からの異なった波長の光を感知し,脳(視覚中枢)の異なったシナプス層に軸索を伸ばしつなぎとめている.ニューロンの軸索がどのように固有のシナプス層を認識し選択をするのか,その分子機構はよくわかっていない.筆者らは,視神経の軸索の投射を制御する2つの細胞表面タンパク質,GogoとFmiがR8視細胞の軸索において機能的に相互作用することを見い出した.Gogo単独のはたらきによりR8視細胞の軸索を一過的にM1層にとどめ,そののち,GogoとFmiとが協調してR8視細胞の軸索を最終的な標的であるM3層へと導いていた.構造機能解析の結果,GogoはFmiと協調する際に細胞内シグナルを伝達する役割を担っていることが示唆された.さらに,R8視細胞の軸索の投射においてFmiは脳の一部のニューロンで必要とされていることがわかり,このことは,R8視細胞のFmiと脳の標的細胞のFmiとがホモ接触結合することによりR8視細胞のM3層への投射の起こることを示唆していた.この研究によって,Gogoは神経軸索の投射においてFmiの機能的なパートナーであり,これら2つのタンパク質の相互作用をダイナミックに制御することにより視神経の軸索のシナプス層の特異的な選択が正確に行われていることが示唆された.
はじめに
ニューロンどうしの正確なシナプスの形成は脳における情報演算処理にとり非常に重要である.シナプスはしばしば機能的な意味あいを反映する構造を形成する.たとえば,視覚系ではとなりあった視細胞は脳の標的領域においてもとなりあった柱状構造に軸索を投射しシナプスを接続する.この特徴的な構造により外の視覚世界からの視覚信号と脳に映し出される映像信号との位置関係を維持している1).この柱状の網膜脳相関維持(retinotopy)構造にくわえ層状の構造もとっており,外界からの映像を異なった成分に分解しそれぞれの成分を別々のシナプス層で情報処理することも行われている.これらのような特徴的な柱状かつ層状の構造は複雑な神経組織において広くみられるものである2).それでは,このシナプスの層状構造はどのような分子機構により形成されているのだろうか.ニューロンの軸索はどのように固有のシナプス層を認識しそこに投射するのか,その分子機構はよくわかっていない.脊椎動物の視覚系では,SidekicksやDscamsなど,いくつかのホモ結合接着分子がそれどうしの接着の選択性を利用して視細胞におけるシナプス層の特異性を決めていることが知られている1).
ショウジョウバエの視覚系も層状の構造をとっており,シナプスの結合に特異性のあることもよく知られている.ショウジョウバエの複眼は約800個の個眼からできており,1つの個眼には8個の視細胞(R1~R8)がある.R1~R6の視細胞は物体の動きを感知し,軸索は最初の視神経節であるラミナに投射する.色覚をつかさどるR7視細胞とR8視細胞は2番目の視神経節であるメダラに軸索を投射する.1つの個眼においてR7視細胞とR8視細胞のペアは空間の同一の点から色信号を受け取り,メダラの同一の柱状の構造に軸索を投射する.興味深いことに,このR7視細胞とR8視細胞とはそれぞれ別々の波長の色信号を感知しているのだが,軸索を別々のメダラ層(R8視細胞はM3層,R7視細胞はM6層に)に投射してシナプスを形成する(図1).このようにして,個々の機能モジュールは別々のシナプス層でプロセスされている.このようなことから,ショウジョウバエの視覚系はシナプス層の特異的な選択機構を研究する格好のモデル系であるといえる1,3).
ショウジョウバエではいくつかの膜貫通型タンパク質がシナプス層に特異的な投射を制御していることが知られている4).そのような例として,カドヘリンの一種であるN-カドヘリンとFlamingo 5-7)(Fmi),受容体型チロシンキナーゼであるLARとPtp69D,接着分子であるCapricious,受容体と予想されるGolden Goal 8)(Gogo)などが知られている.それぞれ個々の機能的な役割は報告されているが,これらのタンパク質が総合的にどのようにかかわりあってシナプス層の特異性を決定しているのかという全体像はよくわかっていない.
筆者らは,R8視細胞に着目して研究を進めた9).このR8視細胞ではFmiが非常に重要な役割を担っていることがわかった.Fmiは,カドヘリンがそうであるように,試験管内では強くホモ結合接着(同じ分子種どうしが接着する)することが知られている5).しかし,生体ではFmiはすべての種類の視細胞に発現しており,しかも,脳の標的領域であるメダラ層においても何層にも広範に発現している.このことは,ほかのタンパク質がFmiの接着活性をタイミングよく制御してR8視細胞の投射を制御していることを示唆していた.筆者らは,Gogoの変異体がFmiの変異体と似たR8視細胞の表現型を示すことから,Fmiの機能的なパートナーはGogoではないかと考えそれを実証した.
1.gogo変異体とfmi変異体は神経系で似た表現型を示す
筆者らがすでに発表した研究により,gogo変異体はR8視細胞の軸索の投射においてfmi変異体6,7) と似た表現型を示すことが知られていた8).それにくわえて,この研究では,gogo変異体がR1~R6の視細胞のラミナでのシナプスの選択においてfmi変異体と同様の間違いを起こすことを見い出した.さらに,gogo変異体は胚の末梢神経系の樹状突起においてもfmi変異体と同様に10),伸びすぎて背中線をこえるという表現型をみせた.GogoとFmiはタンパク質の発現や局在パターンでも相当に重なる部分の多いこともわかった6).これらのことは,この2つのタンパク質が協調的にはたらいていることを示唆していた.
2.gogo変異体とfmi変異体は遺伝学的な相互作用を示す
GogoとFmiとが機能的に協調してはたらくことを示すため,遺伝学的な相互作用をみた.具体的には,Fmiの不完全な変異体(ハイポモルフ)について,それにくわえてGogoの遺伝子を半減したときに異常が強くなるかどうかをみた.その結果,R8視細胞の軸索の投射にくわえて,GogoとFmiとは成体になるまでの生存率や樹状突起網の形成において遺伝学的な相互作用があった.これは,GogoとFmiとの協調的なはたらきが視神経だけではなく広く神経系の全般において起こっていることを示唆していた.
また逆に,野生型ではGogoとFmiが機能していないR7視細胞にこの2つのタンパク質を強制的に共発現してみた.すると,本来はR7視細胞の軸索はM6層に投射するのだが,R8視細胞が投射するM3層に投射するようになることがわかった.このことから,GogoとFmiとを組み合わせることによりR7視細胞の軸索はR8視細胞の軸索の形質を獲得しそのようにふるまうことがわかった.GogoとFmiとはR8視細胞の軸索のシナプス層への投射に必要かつ十分であることが示された.
3.GogoとFmiとは培養細胞の接触面で共局在する
GogoとFmiとが物理的な相互作用をするかどうかの示唆を得るため,培養細胞においてGogoとFmiの局在を調べた.その結果,細胞凝集団において細胞のあいだの接触面にFmiが集積すると,Gogoも必ず集積することがわかった.この共集積はGogoの細胞外ドメインを介して起こることが,Gogoの細胞内ドメインの欠損変異体なども共集積することから示唆された.
4.GogoとFmiは互いの細胞内局在に影響をあたえうる
では,生体でもGogoとFmiとは相互作用するのだろうか? 発生途中の翅の細胞を用いて検証した.野生型の翅の細胞ではFmiは発現し細胞の頂端側に特徴的に局在するのだが,Gogoは発現していない.この状況でGogoを強制発現させると,FmiはGogoにひき連れられて細胞の横面にも局在するようになった.GogoとFmiとがシスあるいはトランスに相互作用するのかを調べるため,Gogoの強制発現とfmi変異体の細胞クローンとをランダムに組み合わせ翅においてモザイク状とした.その細胞クローンの境界を調べた結果,このGogoによるFmiの局在のシフトはGogoとFmiとが同じ細胞に存在するときだけみられた.そこで,GogoとFmiとはシスに相互作用することが示唆された(図2a).
5.Gogoの細胞内ドメインはFmiと協調したときシグナルを伝達する役割を担う
では,GogoはFmiと協調したとき,R8視細胞の軸索の投射においてどのような役割を担っているのだろうか? 以前の研究から,Gogoの細胞内ドメインを欠損させるとgogo変異体の表現型をレスキューできないことがわかっていた8).しかしながら,Fmiの場合はその細胞内ドメインを欠損させてもfmi変異体の表現型をレスキューできた.これらのことから,GogoはFmiとその細胞外ドメインで相互作用し,Gogoの細胞内ドメインを介して投射シグナルをR8視細胞の軸索に伝達しているのではないかと予想できた.これを検証するため,さきのR7視細胞でのGogoとFmiとの共強制発現の系を使い,生体におけるそれらの細胞内ドメインの影響力をみた.FmiとGogoの全長または細胞内ドメインを欠損させた遺伝子を組み合わせた結果,Gogoの細胞内ドメインがR7視細胞の投射先を(本来はR8視細胞の標的である)M3層に変更させるのに必要かつ十分であることがわかった.このことは,投射シグナルをR8視細胞の軸索に伝達することがFmiと協調したときのGogoの重要な役割のひとつであることを示唆していた(図2a).
6.R8視細胞の軸索の標的側のリガンドはFmiである
脳のFmiがR8視細胞の軸索のリガンドになっているかどうかは不明であった.その原因として,脳にfmi変異体の大きな細胞集団クローンを作製するのがむずかしいことがあげられた.そこで,細胞のタイプに特異的なコンディショナルノックアウト細胞を作製するため新しい系を開発して脳にfmi変異体の大きな細胞集団クローンを作製した.脳のラミナニューロンからFmiを取り除くとR8視細胞の軸索の投射が相当な割合で異常となり,M1層でとまってしまうことがわかった.このことから,脳のFmiがR8視細胞の軸索の投射に必要であり,脳のFmiとR8視細胞のFmiとが相互作用をしていることが示唆された(図2a).
7.R8視細胞はGogo単独ではM1層にGogoとFmiの両方ではM3層に誘導される
すでに報告されたGogoの過剰発現では,R8視細胞はM1層に強く接着する表現型を示すことが知られていた8).今回の研究では,Gogoは(Fmiと協調して)M3層に投射するのを助けていた.この2つの実験結果は矛盾せずに説明できるのだろうか? Gogoの過剰発現ではR8視細胞の軸索の途中(M1層)に大きな軸索塊ができた.この状況でFmiを不完全な変異体にすると軸索塊がより大きくなり,Fmiを強制発現すると軸索塊の形成が抑制された.このことから,FmiはM1層の付近ではGogoと拮抗するようにはたらくことがわかった.
以上のことをまとめると,サナギの初期ではGogo単独でM1層に接着し,サナギの中期にR8視細胞がM3層に軸索を伸ばしはじめるときにGogoとFmiとが協調してR8視細胞をM3層に誘導するものと考えられた(図2b).ちなみに,M3層におけるリガンドはFmiだと推察しているが,M1層におけるリガンドは現時点では不明である.
おわりに
この研究によって,細胞の表面にあるタンパク質の活動あるいは活性の組合せを細胞特異的かつ経時的に変化させることによりニューロンのシナプス層に特異的な投射の制御されていることが示唆された.この分子機構は,視神経にかぎらずあらゆる脳神経組織の発生段階においてあてはまる普遍原理的なものである可能性がある.これを理解することは,われわれの五感・意識・思索・行動などのみなもとである脳神経組織の形成の分子機序を理解するうえで非常に重要である.
また,近年,幹細胞の移植などにより神経再生医療の道が開けようとしている.神経系の機能的な回復には,いうまでもなく,神経回路網の再構築が必要である.この研究のように,神経回路網の形成の分子機構を理解することによって,将来は神経再生医療にも役立つ可能性があると考えている.
文 献
- Sanes, J. R. & Zipursky, S. L.: Design principles of insect and vertebrate visual systems. Neuron, 66, 15-36 (2010)[PubMed]
- Huberman, A. D., Clandinin, T. R., Baier, H.: Molecular and cellular mechanisms of lamina-specific axon targeting. Cold Spring Harb. Perspect. Biol., 2, a001743 (2010)[PubMed]
- Takemura, S. Y., Lu, Z. & Meinertzhagen, I. A.: Synaptic circuits of the Drosophila optic lobe: the input terminals to the medulla. J. Comp. Neurol., 509, 493-513 (2008)[PubMed]
- Berger, J., Senti, K. A., Senti, G. et al.: Systematic identification of genes that regulate neuronal wiring in the Drosophila visual system. PLoS Genet., 4, e1000085 (2008)[PubMed]
- Usui, T., Shima, Y., Shimada, Y. et al.: Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell, 98, 585-595 (1999)[PubMed]
- Lee, R. C., Clandinin, T. R., Lee C. H. et al.: The protocadherin Flamingo is required for axon target selection in the Drosophila visual system. Nat. Neurosci., 6, 557-563 (2003)[PubMed]
- Senti, K. A., Usui, T., Boucke, K. et al.: Flamingo regulates R8 axon-axon and axon-target interactions in the Drosophila visual system. Curr. Biol., 13, 828-832 (2003)[PubMed]
- Tomasi, T., Hakeda-Suzuki, S., Ohler, S. et al.: The transmembrane protein Golden goal regulates R8 photoreceptor axon-axon and axon-target interactions. Neuron, 57, 691-704 (2008)[PubMed]
- Ting, C. Y., Yonekura, S., Chung, P. et al.: Drosophila N-cadherin functions in the first stage of the two-stage layer-selection process of R7 photoreceptor afferents. Development, 132, 953-963 (2005)[PubMed]
- Gao, F. B., Kohwi, M., Brenman, J. E. et al.: Control of dendritic field formation in Drosophila: the roles of flamingo and competition between homologous neurons. Neuron, 28, 91-101 (2000)[PubMed]
著者プロフィール
略歴:2000年 東京大学大学院理学系研究科 修了,同年 オーストリアResearch Institute of Molecular Pathology博士研究員を経て,2005年よりドイツMax Planck Institute of Neurobiology博士研究員.
研究テーマ:神経回路の形成,軸索の投射,樹状突起網の形成,アクチン骨格系.
鈴木 崇之(Takashi Suzuki)
略歴:2000年 東京大学大学院理学系研究科 修了,同年 オーストリアResearch Institute of Molecular Pathology博士研究員を経て,2005年よりドイツMax Planck Institute of Neurobiologyグループリーダー.
研究テーマ:神経回路の形成,シナプス形成の決定機構.
抱負:ショウジョウバエには生まれながらに高度な神経活動を遂行できるような神経回路網が備わっている.ヒトと違って神経活動に非依存的なのは驚くべきことだ.その形成機構にせまりたい.
研究室URL:http://www.neuro.mpg.de/english/junior/axguide/index.html
© 2011 鈴木(羽毛田)聡子・鈴木崇之 Licensed under CC 表示 2.1 日本
