生命の根幹にかかわるシグナル受容体のキネシンスーパーファミリーによる細胞内輸送
田中庸介・廣川信隆
(東京大学大学院医学系研究科 分子構造・動態学講座,細胞生物学・解剖学大講座)
email:田中庸介,廣川信隆
DOI: 10.7875/first.author.2011.025
KIF16B/Rab14 molecular motor complex is critical for early embryonic development by transporting FGF receptor.
Hitoshi Ueno, Xiao Huang, Yosuke Tanaka, Nobutaka Hirokawa
Developmental Cell, 20, 60-71 (2011)
FGFは細胞から分泌されるタンパク質で周囲の細胞の表面にあるFGF受容体に結合して分化や増殖のシグナルを伝達する.細胞質で新しく合成されたFGF受容体がどのようにして細胞の表面へと輸送されるのかはほとんど不明であったが,今回,筆者らのグループは,キネシンスーパーファミリーに属する新規の分子モーターKIF16Bの分子遺伝学的な解析をつうじてFGF受容体の輸送の分子機構とその生理学的な意義を解明した.FGF受容体は細胞の中心にあるゴルジ体から小胞によって細胞膜の直下の中継地点であるリサイクリングエンドソームにむかい輸送される.この小胞に含まれるGタンパク質Rab14が活性型になったとき,小胞はRab14を介して分子モーターKIF16Bと直接に結合しリサイクリングエンドソームにむかって微小管のレールを輸送されていくことが明らかとなった.一方,FGFによるシグナル伝達は胎児の発生初期,われわれの体のすべての細胞のおおもととなる内細胞塊の分化に必要である.ところが,KIF16BあるいはRab14の活性が失われたマウス初期胚では細胞表面のFGF受容体の欠乏によるシグナル伝達の低下の結果,内細胞塊がほとんど分化できず胎仔がまったく育たないという重篤な事象が生じた.KIF16BおよびRab14によるFGF受容体の輸送制御はこのように生命の根幹にかかわるものであり,さまざまな発生過程のパターン化をはじめ,幹細胞,成人病,神経機能などとの関連が示唆された.
キネシン分子モーターはすべての細胞において微小管をレールとしてオルガネラや分子複合体を輸送するATP分解酵素である.これまで哺乳類のゲノムには45種類の遺伝子からなるキネシンスーパーファミリーが同定されており1,2),生命の要となる種々の細胞機能に深くかかわっている3).今回,そのうちキネシン3ファミリーに属する新規の分子モーターKIF16Bの欠損胚をマウスにおいて作製したところ,これまでのなかでもっとも重篤な表現型を生じ着床期に死亡した.この表現型はさまざまな細胞現象のもととなるFGF(fibroblast growth factor,線維芽細胞栄養因子)シグナル伝達系4,5) との関係を示唆するものであり,実際に,FGF受容体2の細胞内輸送が障害されていた.そこで,このFGF受容体の細胞内輸送の分子機構を追求したところ,低分子量Gタンパク質Rab14がKIF16Bに直接に結合するアダプタータンパク質としてこの輸送をダイナミックに制御していることを発見した.これは,FGF受容体の翻訳後の細胞内輸送における分子機構のはじめての解明であり,さらに,分子モーターによるシグナル伝達の新しい制御機構を提唱するものであった.
筆者らは,今回,新規の分子モーターKIF16Bのin vivoでのはたらきを調べるため,遺伝子ターゲティング法を用いてKIF16Bの欠損したES細胞を作出し,これを四倍体胚レスキュー(tetraploid rescue)法によって100%がES細胞に由来する初期胚へと直接に発生させその表現型を調べた.野生型の胚は着床の前後において内細胞塊(inner cell mass)とよばれる胚盤胞(blastocyst)の中心的な細胞が胚盤葉上層(epiblast)と胚盤葉下層(primitive endoderm)とに分化し胚盤葉下層の細胞がFGF依存性に基底膜を分泌する(図1).この基底膜と直接に接触することによって胚盤葉上層が成体のすべての細胞を形成する外胚葉・中胚葉・内胚葉の源となるべく上皮化し円筒胚(egg cylinder)となることが知られている.
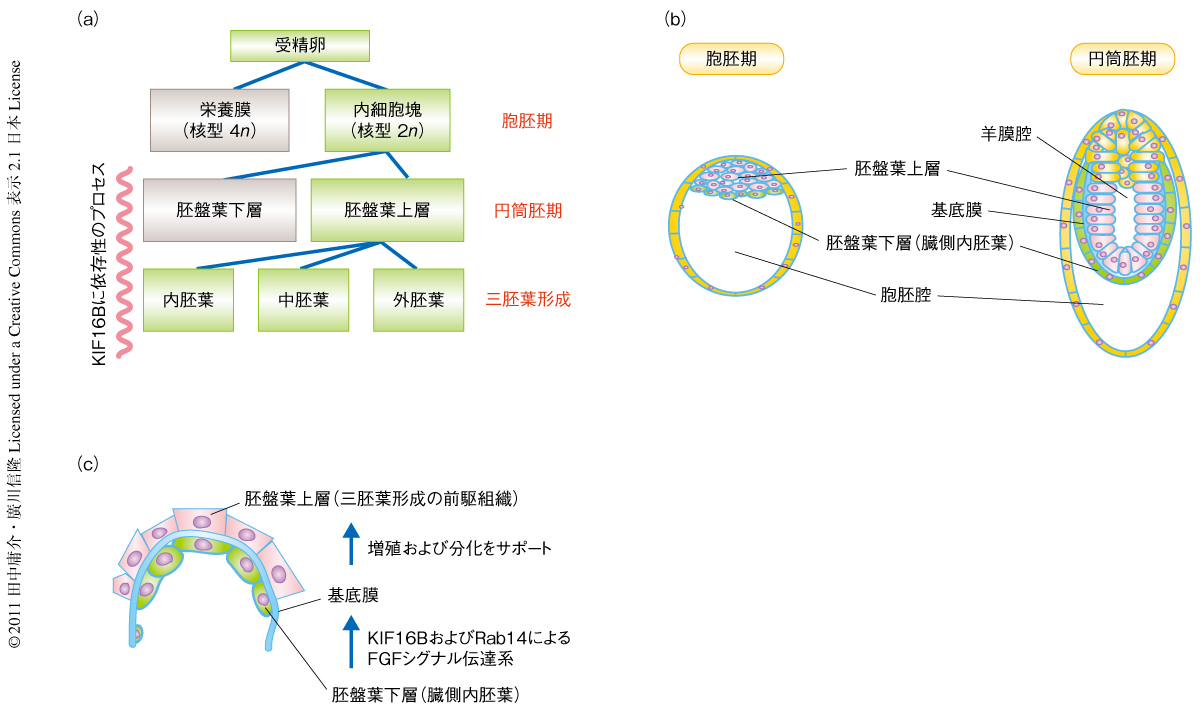
ところが,KIF16B欠損胚の5.5日胚においてはこの円筒胚が形成されず,形態的に胚盤葉上層と胚盤葉下層が不分明であり発生を継続することができなかった.着床子宮のパラフィン切片の免疫組織化学を行ったところ,基底膜のマーカーであるラミニンおよび胚盤葉下層の分化マーカーPemないしGATA6の発現が有意に低下していた.また,胚盤胞の免疫ブロッティングを行ったところ,このGATA6のほか,分化多能性のマーカーであるOct4とSox2の発現も有意に低下していた.すなわち,KIF16Bは未分化マウスES細胞の増殖には必須ではないが,ひとたびこれがFGF依存性の分化サイクルに入るとKIF16Bの欠失によるなんらかのシグナル伝達の欠損によって幹細胞の分化多能性およびその胚盤葉下層への分化能に問題を生じ,そこからの基底膜の分泌が障害されるため結果的に胚の発生を維持できなくなるという経路が考えられた.
着床前後の胚の発生はES細胞を試験管内で集合塊にして胚様体(embryoid body)を形成させこれをin vitroで培養することでモデル化することができる.培養5日で球状の胚様体の外縁に胚盤葉下層が分化し,これが胚盤葉上層の細胞塊を包み込む.FGFシグナル伝達系のはたらきにより胚盤葉下層によって形成された基底膜と接触することが,胚盤葉上層の細胞にとって胚様体の内部の空洞内で上皮様に分化・増殖するため必須だとされている6).ところが,KIF16Bを欠損させたES細胞から胚様体を形成させると成長が悪くなり,胚盤葉上層の細胞の上皮化,胚盤葉下層の細胞の分化(GATA6の発現),ならびに,基底膜の形成(ラミニン1およびフィブロネクチンの発現)が有意に低下し,細胞のアポトーシスが増加していた.さらに,Kif16b遺伝子のcDNAの導入によって基底膜を取り戻すことができたため,これらの表現型はまさしくKIF16Bの欠損によって起こったものと考えられた.
このような表現型をあたえる過去の実験例を調べてみると,FGF受容体2の完全ノックアウトマウスをはじめ7),FGF受容体2のドミナントネガティブ変異体を導入した胚様体,FGF4欠損胚,Grb2欠損胚,FGFあるいはMAPキナーゼの阻害剤で処理した初期胚など,FGFシグナル伝達系カスケードの変異によって生ずるものと酷似していることがわかった.そこで,KIF16Bを欠損したマウス線維芽細胞の細胞株を初期キメラ胚から樹立して調べてみると,KIF16B欠損細胞では確かにFGFによるMAPキナーゼ系およびホスファチジルイノシトール3-キナーゼ系のシグナル伝達が低下していた.そこで,細胞表面のFGF受容体2を培地に入れた抗体により標識して免疫沈降し,あるいは,細胞表面のビオチン化により定量してみると,KIF16B欠損細胞ではその量が有意に低下していることがわかった.さらに,免疫組織化学によりFGF受容体2がKIF16Bの欠損によって細胞核の周囲のエンドソーム様の構造に集積していることが明らかとなり,さらに,Kif16b遺伝子のcDNAの導入によって一様な分布を取り戻すことができたため,KIF16Bが実際にFGF受容体2を輸送しており,その細胞膜への提示に重要な役割をはたしている可能性が示唆された.
KIF16BとFGF受容体2との関係を探るため,密度勾配遠心を用いた細胞分画法を行うとFGF受容体2およびEGF受容体とKIF16Bとが浮遊膜画分に共分画されることがわかった.さらに,小胞の免疫沈降法によりFGF受容体2がKIF16Bと特異的に共沈することがわかった.そこで,異なる蛍光タンパク質によって修飾したFGF受容体2とKIF16Bとを細胞に共発現し共焦点レーザー顕微鏡でタイムラプス観察すると,細胞核の近傍のゴルジ体と思われる領域から細胞膜の近傍の小胞膜が集積したハブ構造にむかって,これら両方のタンパク質を含んだ小胞が輸送されていくことがわかった.このハブ構造の本体を探るため,蛍光タンパク質によって修飾したFGF受容体2を発現した細胞の底面を全反射照明蛍光(total internal reflection fluorescence:TIRF)顕微鏡を用いて観察した.すると,この小胞膜の集積したハブ構造に存在するFGF受容体2は,とくに培地の血清濃度を減少させた直後においてエキソサイトーシスをへて細胞膜と融合し拡散していくという傾向がみられた.これらのことから,新規に合成されたFGF受容体の細胞膜への提示機構について,以下のような仮説が考えられた.新しく合成されたFGF受容体2はゴルジ体から小胞に包まれて出芽し,分子モーターKIF16Bにより微小管をレールとして輸送され細胞膜の直下のハブ構造に集積する.さらに,このハブ構造から一定の条件において細胞膜への組み込みが起こり細胞膜にFGF受容体2が提示されるものと思われた(図2a).
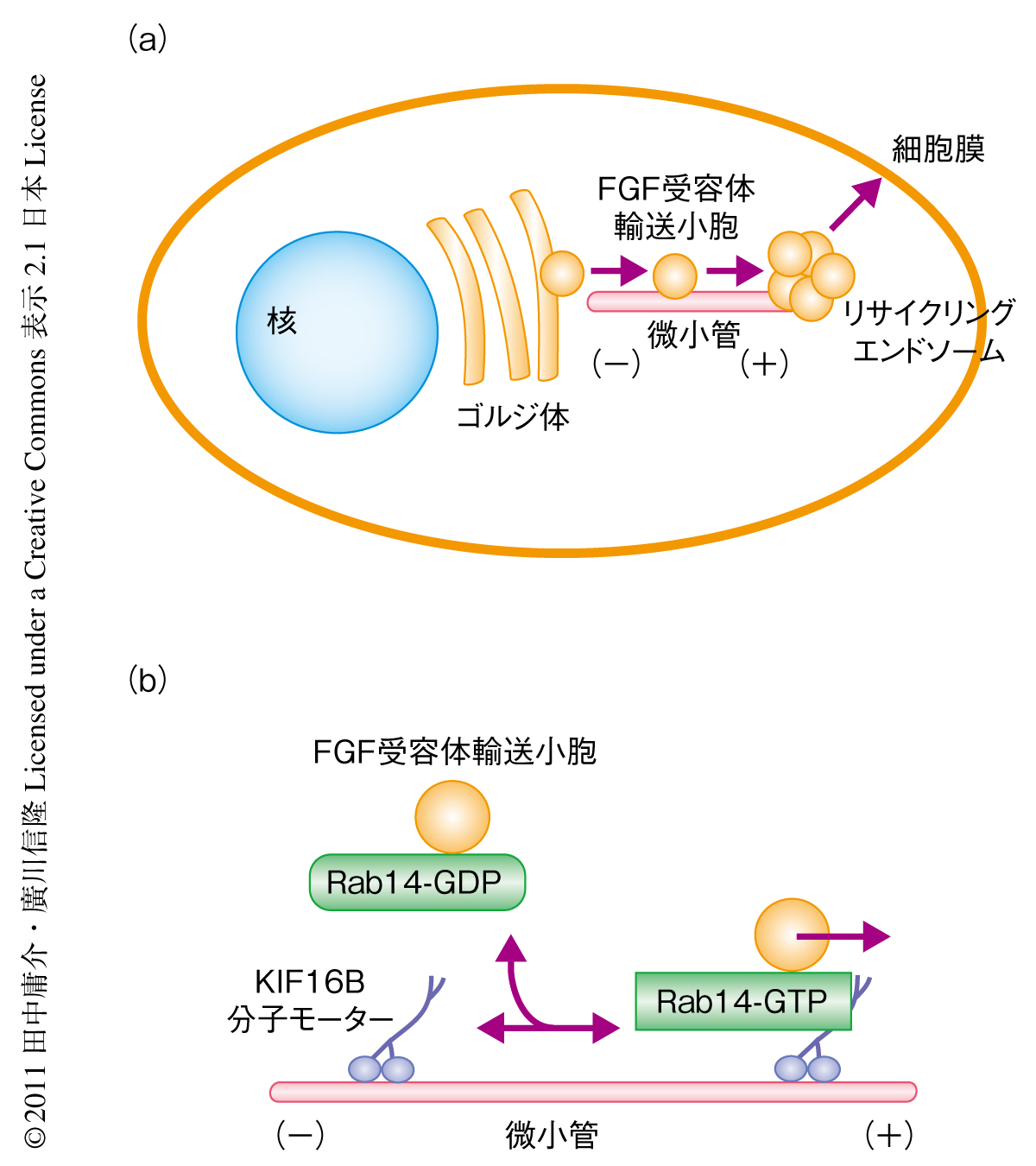
KIF16Bによる小胞輸送の分子機構をさらに詳細に追究するため,酵母ツーハイブリッド法を用いてKIF16Bの尾部に直接に結合するタンパク質をスクリーニングしたところ,低分子量Gタンパク質のひとつであるRab14の存在が示唆された.Rab14はこれまでの研究により新規に合成されたタンパク質のゴルジ体からリサイクリングエンドソームにむけた小胞輸送への関与が示唆されている8,9).また,Rabタンパク質はGTPおよびGDPの結合サイクルによってそのエフェクタータンパク質との結合能を変化させることが知られており,筆者らのグループは,以前に,Rab3とDENN/MADDの結合および解離がキネシン3ファミリーに属するほかの分子モーター,KIF1AとKIF1Bβのニューロンでのシナプス小胞前駆体の軸索輸送における制御をつかさどることを明らかにしている10).そこで,GTP結合型およびGDP結合型の分子構造を模したRab14変異体を細胞に発現して免疫沈降法を行ったところ,GTP結合型Rab14が特異的にKIF16Bに結合することが示された.また,GTPアナログの存在に特異的にKIF16B抗体によりRab14の共沈されることも示された.
つぎに,蛍光タンパク質で標識したRab14変異体を野生型の細胞に発現させると,GTP結合型Rab14変異体のみが有意にKIF16Bと共局在し細胞質の全体に分布することがわかった.GDP結合型Rab14変異体は核周囲に集積した.ところが,KIF16B欠損細胞ではいずれのRab14変異体も核周囲に集積した.これらのことから,KIF16BはGTP結合型Rab14に直接に結合しゴルジ体からリサイクリングエンドソームへの小胞輸送に関与していることが示唆された.
Rab14とFGF受容体2との関係を探るため,蛍光タンパク質で標識したRab14変異体を発現させた細胞をFGF受容体2により免疫染色してみると,GTP結合型変異体およびGDP結合型変異体ともにFGF受容体2と共局在するが,GDP結合型Rab14変異体を導入するとFGF受容体2が有意に核周囲に集積してしまうことがわかった.実際に,小胞免疫沈降法を行うとFGF受容体2はGTP結合型変異体およびGDP結合型変異体と共沈されるが,Rab14がGTP型変異体のときにのみKIF16Bとも共沈することがわかった.すなわち,Rab14はFGF受容体2を含有する小胞につねに結合しており,Rab14のヌクレオチド結合状態によってそのKIF16Bとの直接の結合による細胞の周縁にむけた輸送が制御されていると考えられた(図2b).
さらに,このRab14のヌクレオチド結合状態が実際に細胞のシグナル伝達を制御しているかどうかを確かめるため,GTP結合型変異体およびGDP結合型変異体をレンチウイルスベクターにより導入しFGFによるMAPキナーゼシグナル伝達を測定した.その結果,GDP結合型Rab14変異体はドミナントネガティブ変異体としてはたらきKIF16BによるFGF受容体2の輸送ならびにFGFシグナル伝達系を阻害することが明らかとなった.
最後に,このRab14によるシグナル伝達の制御が実際の胚発生に関与しているかどうかを確かめるため,GDP結合型Rab14変異体(ドミナントネガティブ変異体)で形質転換したES細胞株を用いて胚様体の形成アッセイを行った.その結果,ドミナントネガティブ変異体によって胚様体の成長,基底膜の形成,ならびに,胚盤葉上層の上皮化が有意に低下し,KIF16B欠損の表現型を再現することができた.すなわち,初期胚の発生および分化に必須なFGFによるシグナル伝達を直接に制御する新規の経路として,KIF16BおよびRab14によるFGF受容体2を含む小胞輸送系を同定することができた.
新しく生合成された栄養因子受容体がどのように細胞膜に提示されるか,また,その過程がどのように制御されシグナル伝達系の調節に関与しているかについては,これまでほとんど手がつけられてこなかった.筆者らは,分子モーターの細胞生物学からこの問題に新たなアプローチを試み,FGF受容体を輸送するキネシン分子モーターを同定し,Rabタンパク質との結合および解離による制御機構を発見し,さらに,その初期発生における役割を明らかにすることができた.これは,分子モーターが幹細胞の分化に必須であることを示すはじめての例であり,再生医学,発がんの機構,神経機能などの研究にも応用が可能と思われる.新規に生合成されたFGF受容体が微小管系の細胞内小胞輸送につづき,リサイクリングエンドソームと思われるハブ構造を介して細胞膜に提示されるという分子機構はきわめてユニークであり,Rabタンパク質のGTPase活性化タンパク質(GTPase-activating potein:GAP)やグアニンヌクレオチド交換因子(guanine nucleotide exchange factor:GEF)の活性化による精密な制御が予想される.このような分子モーターによるシグナル伝達のダイナミックな制御はほかの膜受容体の場合にも共通する機構であると思われ,今後,さらに詳細に検討をくわえていきたい.
略歴:1996年 東京大学大学院医学系研究科 退学,同年より同 助手.
研究テーマ:キネシンスーパーファミリーの分子遺伝学および細胞生物学.
関心事:物質輸送がもたらす細胞挙動の変化.
廣川 信隆(Nobutaka Hirokawa)
略歴:1971年 東京大学医学部 卒業,1978年 医学博士号 取得,1982年 米国Washington大学医学部 助教授,1983年 同 准教授を経て,1983年より東京大学医学部(現 大学院医学系研究科)教授.
研究テーマ:キネシンスーパーファミリーの分子細胞生物学,分子遺伝学,構造生物学的な研究.
関心事:細胞内物質輸送の分子機構の解明と,キネシンスーパーファミリーの研究をとおして生命科学の新しい扉を開くこと.
© 2011 田中庸介・廣川信隆 Licensed under CC 表示 2.1 日本
(東京大学大学院医学系研究科 分子構造・動態学講座,細胞生物学・解剖学大講座)
email:田中庸介,廣川信隆
DOI: 10.7875/first.author.2011.025
KIF16B/Rab14 molecular motor complex is critical for early embryonic development by transporting FGF receptor.
Hitoshi Ueno, Xiao Huang, Yosuke Tanaka, Nobutaka Hirokawa
Developmental Cell, 20, 60-71 (2011)
要 約
FGFは細胞から分泌されるタンパク質で周囲の細胞の表面にあるFGF受容体に結合して分化や増殖のシグナルを伝達する.細胞質で新しく合成されたFGF受容体がどのようにして細胞の表面へと輸送されるのかはほとんど不明であったが,今回,筆者らのグループは,キネシンスーパーファミリーに属する新規の分子モーターKIF16Bの分子遺伝学的な解析をつうじてFGF受容体の輸送の分子機構とその生理学的な意義を解明した.FGF受容体は細胞の中心にあるゴルジ体から小胞によって細胞膜の直下の中継地点であるリサイクリングエンドソームにむかい輸送される.この小胞に含まれるGタンパク質Rab14が活性型になったとき,小胞はRab14を介して分子モーターKIF16Bと直接に結合しリサイクリングエンドソームにむかって微小管のレールを輸送されていくことが明らかとなった.一方,FGFによるシグナル伝達は胎児の発生初期,われわれの体のすべての細胞のおおもととなる内細胞塊の分化に必要である.ところが,KIF16BあるいはRab14の活性が失われたマウス初期胚では細胞表面のFGF受容体の欠乏によるシグナル伝達の低下の結果,内細胞塊がほとんど分化できず胎仔がまったく育たないという重篤な事象が生じた.KIF16BおよびRab14によるFGF受容体の輸送制御はこのように生命の根幹にかかわるものであり,さまざまな発生過程のパターン化をはじめ,幹細胞,成人病,神経機能などとの関連が示唆された.
はじめに
キネシン分子モーターはすべての細胞において微小管をレールとしてオルガネラや分子複合体を輸送するATP分解酵素である.これまで哺乳類のゲノムには45種類の遺伝子からなるキネシンスーパーファミリーが同定されており1,2),生命の要となる種々の細胞機能に深くかかわっている3).今回,そのうちキネシン3ファミリーに属する新規の分子モーターKIF16Bの欠損胚をマウスにおいて作製したところ,これまでのなかでもっとも重篤な表現型を生じ着床期に死亡した.この表現型はさまざまな細胞現象のもととなるFGF(fibroblast growth factor,線維芽細胞栄養因子)シグナル伝達系4,5) との関係を示唆するものであり,実際に,FGF受容体2の細胞内輸送が障害されていた.そこで,このFGF受容体の細胞内輸送の分子機構を追求したところ,低分子量Gタンパク質Rab14がKIF16Bに直接に結合するアダプタータンパク質としてこの輸送をダイナミックに制御していることを発見した.これは,FGF受容体の翻訳後の細胞内輸送における分子機構のはじめての解明であり,さらに,分子モーターによるシグナル伝達の新しい制御機構を提唱するものであった.
1.KIF16B欠損胚は着床初期の発生に問題を生ずる
筆者らは,今回,新規の分子モーターKIF16Bのin vivoでのはたらきを調べるため,遺伝子ターゲティング法を用いてKIF16Bの欠損したES細胞を作出し,これを四倍体胚レスキュー(tetraploid rescue)法によって100%がES細胞に由来する初期胚へと直接に発生させその表現型を調べた.野生型の胚は着床の前後において内細胞塊(inner cell mass)とよばれる胚盤胞(blastocyst)の中心的な細胞が胚盤葉上層(epiblast)と胚盤葉下層(primitive endoderm)とに分化し胚盤葉下層の細胞がFGF依存性に基底膜を分泌する(図1).この基底膜と直接に接触することによって胚盤葉上層が成体のすべての細胞を形成する外胚葉・中胚葉・内胚葉の源となるべく上皮化し円筒胚(egg cylinder)となることが知られている.
ところが,KIF16B欠損胚の5.5日胚においてはこの円筒胚が形成されず,形態的に胚盤葉上層と胚盤葉下層が不分明であり発生を継続することができなかった.着床子宮のパラフィン切片の免疫組織化学を行ったところ,基底膜のマーカーであるラミニンおよび胚盤葉下層の分化マーカーPemないしGATA6の発現が有意に低下していた.また,胚盤胞の免疫ブロッティングを行ったところ,このGATA6のほか,分化多能性のマーカーであるOct4とSox2の発現も有意に低下していた.すなわち,KIF16Bは未分化マウスES細胞の増殖には必須ではないが,ひとたびこれがFGF依存性の分化サイクルに入るとKIF16Bの欠失によるなんらかのシグナル伝達の欠損によって幹細胞の分化多能性およびその胚盤葉下層への分化能に問題を生じ,そこからの基底膜の分泌が障害されるため結果的に胚の発生を維持できなくなるという経路が考えられた.
2.KIF16B欠損によって胚盤葉下層の細胞は基底膜を分泌できず胚盤葉上層の細胞の分化および増殖に影響を及ぼす
着床前後の胚の発生はES細胞を試験管内で集合塊にして胚様体(embryoid body)を形成させこれをin vitroで培養することでモデル化することができる.培養5日で球状の胚様体の外縁に胚盤葉下層が分化し,これが胚盤葉上層の細胞塊を包み込む.FGFシグナル伝達系のはたらきにより胚盤葉下層によって形成された基底膜と接触することが,胚盤葉上層の細胞にとって胚様体の内部の空洞内で上皮様に分化・増殖するため必須だとされている6).ところが,KIF16Bを欠損させたES細胞から胚様体を形成させると成長が悪くなり,胚盤葉上層の細胞の上皮化,胚盤葉下層の細胞の分化(GATA6の発現),ならびに,基底膜の形成(ラミニン1およびフィブロネクチンの発現)が有意に低下し,細胞のアポトーシスが増加していた.さらに,Kif16b遺伝子のcDNAの導入によって基底膜を取り戻すことができたため,これらの表現型はまさしくKIF16Bの欠損によって起こったものと考えられた.
3.KIF16B欠損細胞はFGF受容体2の細胞膜への提示に問題を生じる
このような表現型をあたえる過去の実験例を調べてみると,FGF受容体2の完全ノックアウトマウスをはじめ7),FGF受容体2のドミナントネガティブ変異体を導入した胚様体,FGF4欠損胚,Grb2欠損胚,FGFあるいはMAPキナーゼの阻害剤で処理した初期胚など,FGFシグナル伝達系カスケードの変異によって生ずるものと酷似していることがわかった.そこで,KIF16Bを欠損したマウス線維芽細胞の細胞株を初期キメラ胚から樹立して調べてみると,KIF16B欠損細胞では確かにFGFによるMAPキナーゼ系およびホスファチジルイノシトール3-キナーゼ系のシグナル伝達が低下していた.そこで,細胞表面のFGF受容体2を培地に入れた抗体により標識して免疫沈降し,あるいは,細胞表面のビオチン化により定量してみると,KIF16B欠損細胞ではその量が有意に低下していることがわかった.さらに,免疫組織化学によりFGF受容体2がKIF16Bの欠損によって細胞核の周囲のエンドソーム様の構造に集積していることが明らかとなり,さらに,Kif16b遺伝子のcDNAの導入によって一様な分布を取り戻すことができたため,KIF16Bが実際にFGF受容体2を輸送しており,その細胞膜への提示に重要な役割をはたしている可能性が示唆された.
4.KIF16BはFGF受容体2を含んだ小胞と結合しこれを輸送している
KIF16BとFGF受容体2との関係を探るため,密度勾配遠心を用いた細胞分画法を行うとFGF受容体2およびEGF受容体とKIF16Bとが浮遊膜画分に共分画されることがわかった.さらに,小胞の免疫沈降法によりFGF受容体2がKIF16Bと特異的に共沈することがわかった.そこで,異なる蛍光タンパク質によって修飾したFGF受容体2とKIF16Bとを細胞に共発現し共焦点レーザー顕微鏡でタイムラプス観察すると,細胞核の近傍のゴルジ体と思われる領域から細胞膜の近傍の小胞膜が集積したハブ構造にむかって,これら両方のタンパク質を含んだ小胞が輸送されていくことがわかった.このハブ構造の本体を探るため,蛍光タンパク質によって修飾したFGF受容体2を発現した細胞の底面を全反射照明蛍光(total internal reflection fluorescence:TIRF)顕微鏡を用いて観察した.すると,この小胞膜の集積したハブ構造に存在するFGF受容体2は,とくに培地の血清濃度を減少させた直後においてエキソサイトーシスをへて細胞膜と融合し拡散していくという傾向がみられた.これらのことから,新規に合成されたFGF受容体の細胞膜への提示機構について,以下のような仮説が考えられた.新しく合成されたFGF受容体2はゴルジ体から小胞に包まれて出芽し,分子モーターKIF16Bにより微小管をレールとして輸送され細胞膜の直下のハブ構造に集積する.さらに,このハブ構造から一定の条件において細胞膜への組み込みが起こり細胞膜にFGF受容体2が提示されるものと思われた(図2a).
5.KIF16BはGTP結合型Rab14に直接に結合しそれを輸送する
KIF16Bによる小胞輸送の分子機構をさらに詳細に追究するため,酵母ツーハイブリッド法を用いてKIF16Bの尾部に直接に結合するタンパク質をスクリーニングしたところ,低分子量Gタンパク質のひとつであるRab14の存在が示唆された.Rab14はこれまでの研究により新規に合成されたタンパク質のゴルジ体からリサイクリングエンドソームにむけた小胞輸送への関与が示唆されている8,9).また,Rabタンパク質はGTPおよびGDPの結合サイクルによってそのエフェクタータンパク質との結合能を変化させることが知られており,筆者らのグループは,以前に,Rab3とDENN/MADDの結合および解離がキネシン3ファミリーに属するほかの分子モーター,KIF1AとKIF1Bβのニューロンでのシナプス小胞前駆体の軸索輸送における制御をつかさどることを明らかにしている10).そこで,GTP結合型およびGDP結合型の分子構造を模したRab14変異体を細胞に発現して免疫沈降法を行ったところ,GTP結合型Rab14が特異的にKIF16Bに結合することが示された.また,GTPアナログの存在に特異的にKIF16B抗体によりRab14の共沈されることも示された.
つぎに,蛍光タンパク質で標識したRab14変異体を野生型の細胞に発現させると,GTP結合型Rab14変異体のみが有意にKIF16Bと共局在し細胞質の全体に分布することがわかった.GDP結合型Rab14変異体は核周囲に集積した.ところが,KIF16B欠損細胞ではいずれのRab14変異体も核周囲に集積した.これらのことから,KIF16BはGTP結合型Rab14に直接に結合しゴルジ体からリサイクリングエンドソームへの小胞輸送に関与していることが示唆された.
6.Rab14はFGF受容体2を含んだ小胞に分布しKIF16Bとの結合を調節する
Rab14とFGF受容体2との関係を探るため,蛍光タンパク質で標識したRab14変異体を発現させた細胞をFGF受容体2により免疫染色してみると,GTP結合型変異体およびGDP結合型変異体ともにFGF受容体2と共局在するが,GDP結合型Rab14変異体を導入するとFGF受容体2が有意に核周囲に集積してしまうことがわかった.実際に,小胞免疫沈降法を行うとFGF受容体2はGTP結合型変異体およびGDP結合型変異体と共沈されるが,Rab14がGTP型変異体のときにのみKIF16Bとも共沈することがわかった.すなわち,Rab14はFGF受容体2を含有する小胞につねに結合しており,Rab14のヌクレオチド結合状態によってそのKIF16Bとの直接の結合による細胞の周縁にむけた輸送が制御されていると考えられた(図2b).
さらに,このRab14のヌクレオチド結合状態が実際に細胞のシグナル伝達を制御しているかどうかを確かめるため,GTP結合型変異体およびGDP結合型変異体をレンチウイルスベクターにより導入しFGFによるMAPキナーゼシグナル伝達を測定した.その結果,GDP結合型Rab14変異体はドミナントネガティブ変異体としてはたらきKIF16BによるFGF受容体2の輸送ならびにFGFシグナル伝達系を阻害することが明らかとなった.
7.Rab14はFGFによるシグナル伝達を調節し初期胚の発生に必須である
最後に,このRab14によるシグナル伝達の制御が実際の胚発生に関与しているかどうかを確かめるため,GDP結合型Rab14変異体(ドミナントネガティブ変異体)で形質転換したES細胞株を用いて胚様体の形成アッセイを行った.その結果,ドミナントネガティブ変異体によって胚様体の成長,基底膜の形成,ならびに,胚盤葉上層の上皮化が有意に低下し,KIF16B欠損の表現型を再現することができた.すなわち,初期胚の発生および分化に必須なFGFによるシグナル伝達を直接に制御する新規の経路として,KIF16BおよびRab14によるFGF受容体2を含む小胞輸送系を同定することができた.
おわりに
新しく生合成された栄養因子受容体がどのように細胞膜に提示されるか,また,その過程がどのように制御されシグナル伝達系の調節に関与しているかについては,これまでほとんど手がつけられてこなかった.筆者らは,分子モーターの細胞生物学からこの問題に新たなアプローチを試み,FGF受容体を輸送するキネシン分子モーターを同定し,Rabタンパク質との結合および解離による制御機構を発見し,さらに,その初期発生における役割を明らかにすることができた.これは,分子モーターが幹細胞の分化に必須であることを示すはじめての例であり,再生医学,発がんの機構,神経機能などの研究にも応用が可能と思われる.新規に生合成されたFGF受容体が微小管系の細胞内小胞輸送につづき,リサイクリングエンドソームと思われるハブ構造を介して細胞膜に提示されるという分子機構はきわめてユニークであり,Rabタンパク質のGTPase活性化タンパク質(GTPase-activating potein:GAP)やグアニンヌクレオチド交換因子(guanine nucleotide exchange factor:GEF)の活性化による精密な制御が予想される.このような分子モーターによるシグナル伝達のダイナミックな制御はほかの膜受容体の場合にも共通する機構であると思われ,今後,さらに詳細に検討をくわえていきたい.
文 献
- Aizawa, H., Sekine, Y., Takemura, R. et al.: Kinesin family in murine central nervous system. J. Cell Biol., 119, 1287-1296 (1992)[PubMed]
- Miki, H., Okada, Y. & Hirokawa, N.: Analysis of the kinesin superfamily: insights into structure and function. Trends Cell Biol., 15, 467-476 (2005)[PubMed]
- Hirokawa, N., Noda, Y., Tanaka, Y. et al.: Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell. Biol., 10, 682-696 (2009)[PubMed]
- Tanaka, Y., Okada, Y. & Hirokawa, N.: FGF-induced vesicular release of Sonic hedgehog and retinoic acid in leftward nodal flow is critical for left-right determination. Nature, 435, 172-177 (2005)[PubMed]
- Turner, N. & Grose, R.: Fibroblast growth factor signalling: from development to cancer. Nat. Rev. Cancer, 10, 116-129 (2010)[PubMed]
- Li, L., Arman, E., Ekblom, P. et al.: Distinct GATA6- and laminin-dependent mechanisms regulate endodermal and ectodermal embryonic stem cell fates. Development, 131, 5277-5286 (2004)[PubMed]
- Arman, E., Haffner-Krausz, R., Chen, Y. et al.: Targeted disruption of fibroblast growth factor (FGF) receptor 2 suggests a role for FGF signaling in pregastrulation mammalian development. Proc. Natl. Acad. Sci. USA, 95, 5082-5087 (1998)[PubMed]
- Junutula, J. R., De Maziere, A. M., Peden, A. A. et al.: Rab14 is involved in membrane trafficking between the Golgi complex and endosomes. Mol. Biol. Cell, 15, 2218-2229 (2004)[PubMed]
- Kitt, K. N., Hernandez-Deviez, D., Ballantyne, S. D. et al.: Rab14 regulates apical targeting in polarized epithelial cells. Traffic, 9, 1218-1231 (2008)[PubMed]
- Niwa, S., Tanaka, Y. & Hirokawa, N.: KIF1Bβ- and KIF1A-mediated axonal transport of presynaptic regulator Rab3 occurs in a GTP-dependent manner through DENN/MADD. Nat. Cell Biol., 10, 1269-1279 (2008)[PubMed]
著者プロフィール
略歴:1996年 東京大学大学院医学系研究科 退学,同年より同 助手.
研究テーマ:キネシンスーパーファミリーの分子遺伝学および細胞生物学.
関心事:物質輸送がもたらす細胞挙動の変化.
廣川 信隆(Nobutaka Hirokawa)
略歴:1971年 東京大学医学部 卒業,1978年 医学博士号 取得,1982年 米国Washington大学医学部 助教授,1983年 同 准教授を経て,1983年より東京大学医学部(現 大学院医学系研究科)教授.
研究テーマ:キネシンスーパーファミリーの分子細胞生物学,分子遺伝学,構造生物学的な研究.
関心事:細胞内物質輸送の分子機構の解明と,キネシンスーパーファミリーの研究をとおして生命科学の新しい扉を開くこと.
© 2011 田中庸介・廣川信隆 Licensed under CC 表示 2.1 日本
