転写因子により植物の1次細胞壁を肥厚させる
坂本真吾・光田展隆
(産業技術総合研究所生物プロセス研究部門 植物機能制御研究グループ)
email:光田展隆
DOI: 10.7875/first.author.2018.110
Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis.
Shingo Sakamoto, Marc Somssich, Miyuki T. Nakata, Faride Unda, Kimie Atsuzawa, Yasuko Kaneko, Ting Wang, Anne-Maarit Bågman, Allison Gaudinier, Kouki Yoshida, Siobhan M. Brady, Shawn D. Mansfield, Staffan Persson, Nobutaka Mitsuda
Nature Plants, 4, 777-783 (2018)
植物の1次細胞壁は植物にとり必須の構造であるが,これまで,その形成を制御する転写因子についてはあまりよくわかっていない.この研究において,筆者らは,シロイヌナズナを用いて,茎の繊維細胞に2次細胞壁の形成されないnst1 nst3二重変異体にさまざまな転写因子を繊維細胞に特異的に発現させることにより,1次細胞壁を肥厚させる転写因子としてクラスIIIdおよびクラスIIIeのERF転写因子ファミリーを同定した.繊維細胞にクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体においては,2次細胞壁をもたない茎の繊維細胞に1次細胞壁のような特徴をもつ細胞壁が形成された.すなわち,2次細胞壁を1次細胞壁に置き換えたような植物になった.クラスIIIdおよびクラスIIIeのERF転写因子ファミリーはさまざまな部位にあまり重なることなく発現しており,全体として1次細胞壁の形成を制御すると考えられた.1次細胞壁は,セルロースが酵素により糖化されやすいという特長にくわえ,果実の食感や保存性を左右するほか,細胞へのマイクロインジェクションにおいて障害になるなど,植物バイオテクノロジーにとり重要な要素であり,今回の発見が今後の研究開発に貢献することが期待される.
植物の細胞壁は1次細胞壁と2次細胞壁に大別される.1次細胞壁は比較的薄くすべての細胞がもち,植物の細胞を特徴づける構造のひとつである.2次細胞壁は茎など強度の必要な組織の繊維細胞,水分を全身に供給するため水圧に耐える必要のある道管,自然に開裂するため張力を蓄える必要のある葯や果実莢の内皮細胞に形成される,厚い細胞壁である.どちらも細胞膜の外側に形成されるが,2次細胞壁は1次細胞壁と細胞膜のあいだに形成される.すなわち,かならず1次細胞壁,2次細胞壁の順に形成され,一般には,細胞が伸長しているときに再編成をくり返しながら形成されるのが1次細胞壁で,伸長が停止した細胞において形成されるのが2次細胞壁と定義される.構成成分の観点からは,1次細胞壁はおもにセルロース,ヘミセルロース,ペクチンから構成されるが,2次細胞壁はおもにセルロース,ヘミセルロース,リグニンから構成される.どちらにもヘミセルロースが含まれるが,ヘミセルロースはアルカリに可溶性の鎖状ヘテロ多糖類の総称であり,1次細胞壁のヘミセルロースはキシログルカンに富み,2次細胞壁のヘミセルロースはキシラン(あるいは,グルコマンナン)に富むといった特徴がある.また,1次細胞壁のみにみられるペクチンは酸に可溶性の鎖状ヘテロ多糖類であり,構造的にはヘミセルロースに近い.セルロースはどちらにも含まれるが,1次細胞壁と2次細胞壁とでは重合度や結晶化度が異なる.リグニンはおもに2次細胞壁に沈着しているが,一部の1次細胞壁にも沈着がみられる.
これまで,それぞれの構成成分の合成にかかわる酵素の同定が進んできたほか,2次細胞壁に関しては,形成を遺伝子発現のレベルで制御する転写因子が多く同定されてきた.なかでも,マスター転写因子と考えられているのがVND転写因子ファミリー1,2) およびNST転写因子ファミリー3,4) である.これらの転写因子ファミリーは植物に特異的な巨大な転写因子ファミリーであるNAC転写因子ファミリーに属するが,同じサブファミリーに属しており相同性は高い.VND転写因子ファミリーはシロイヌナズナにおいて道管の形成を制御しており,道管において2次細胞壁の形成からプログラム細胞死の制御までを担う1,2).NST転写因子ファミリーはシロイヌナズナにおいて茎の繊維細胞3),葯4),果実莢5) の内皮細胞において2次細胞壁の形成を制御しており,nst1 nst3二重変異体においては茎や胚軸の繊維細胞における2次細胞壁の形成が完全に失われる3)(図1).植物において2次細胞壁は一般に乾燥重量の過半をしめる巨大な代謝産物であることから,筆者らは,nst1 nst3二重変異体を器だけあって中身のない植物ととらえ,繊維細胞に特異的にさまざまな転写因子を発現させることにより,新しい細胞壁を形成するような転写因子を探索した6)(図1).
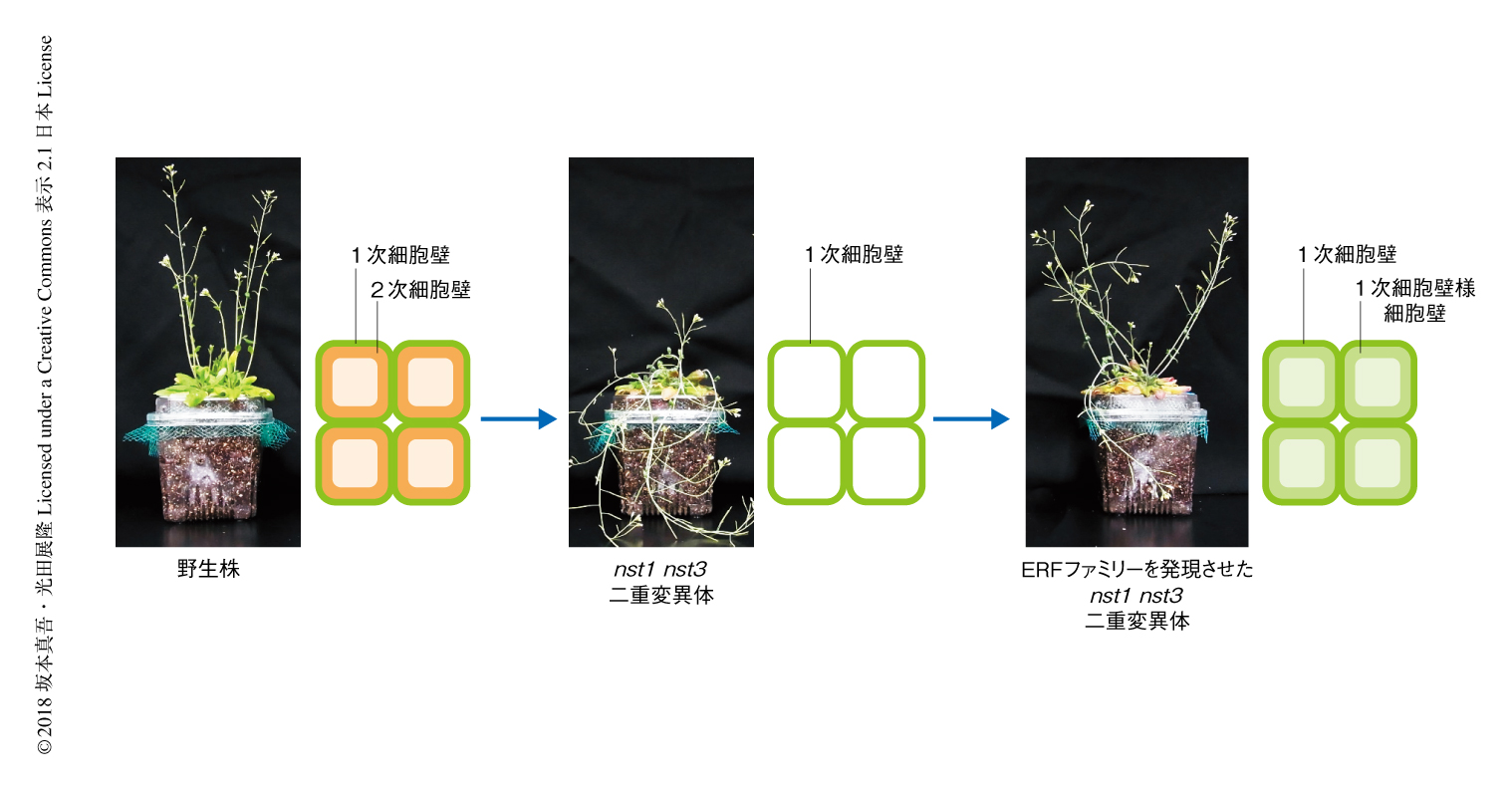
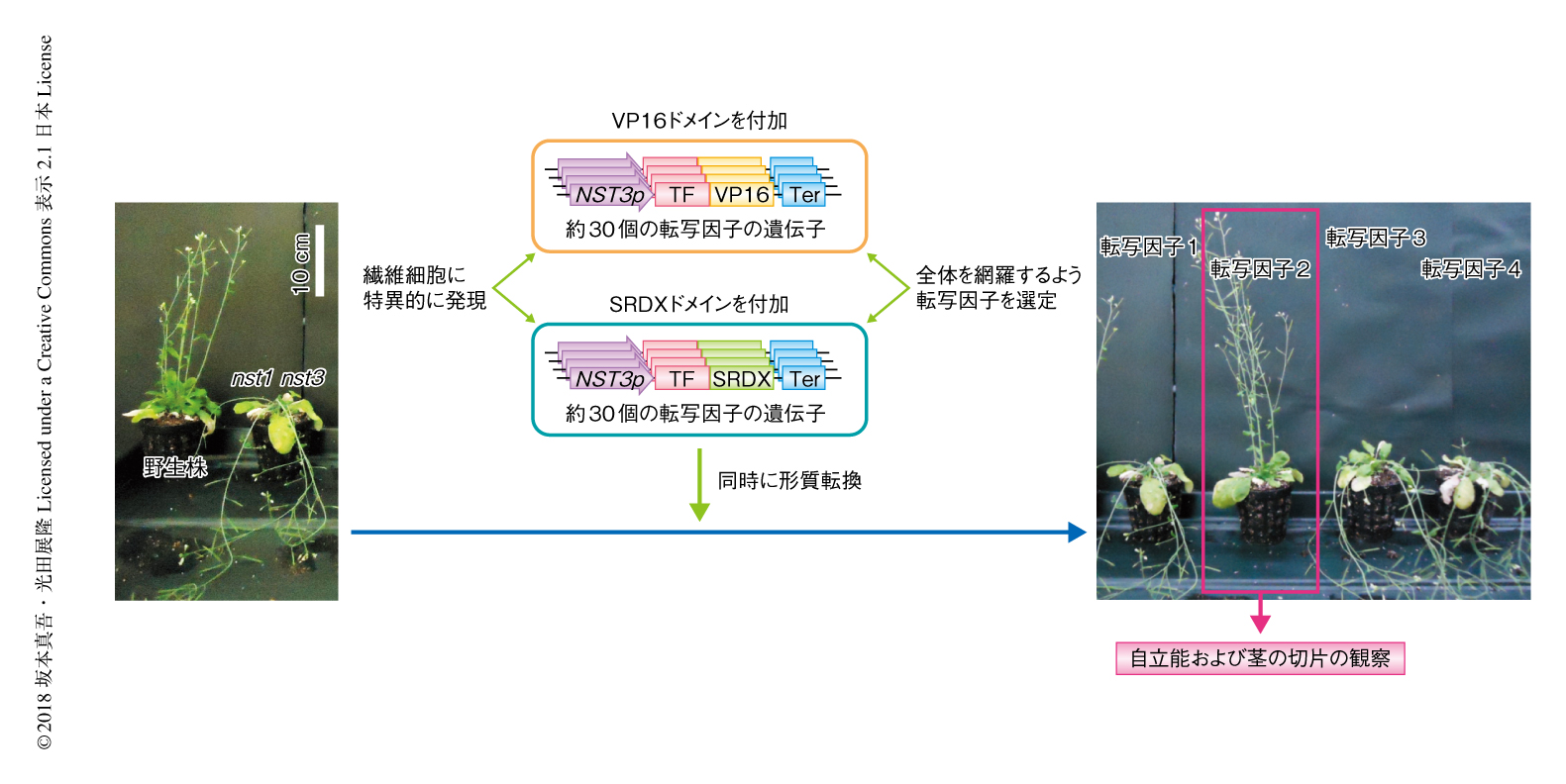
茎の繊維細胞において2次細胞壁の形成が失われたnst1 nst3変異体は,一定の高さまで茎が成長すると自重に耐えきれず倒れるという表現型を示す3)(図1).また,茎は非常に折れやすく手で曲げることにより野生株との違いがはっきりとわかる.シロイヌナズナのもつすべての転写因子約1900個からおのおののサブファミリーを代表する転写因子をひとつずつ選出し,合計で約300個の転写因子をスクリーニングの対象とした.これらに転写活性化ドメインであるVP16ドメインあるいは転写抑制化ドメインであるSRDXドメインを付加し,約30個ずつまとめて繊維細胞に特異的な遺伝子発現を誘導するNST3プロモーターによりnst1 nst3変異体に発現させ,手曲げ試験や徒手切片の観察によりスクリーニングを実施した6).その結果,さまざまな転写因子が表現型を部分的に回復させ,その多くでは2次細胞壁の形成が部分的に回復していた6).しかし,VP16ドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させた場合には,細胞壁が肥厚しているにもかかわらず,通常の2次細胞壁にみられるリグニンの沈着はまったくみられなかった.この肥厚した細胞壁においては,リグニンがないだけでなく,通常の2次細胞壁にみられるキシランも確認されなかった.透過型電子顕微鏡により超微細構造を観察したところ,通常の2次細胞壁に近い細胞壁の形成がみられたものの,細胞壁は層状の構造をとるようにみえたほか,細胞壁の輪郭がなめらかでないようにみえた.茎の強度もnst1 nst3二重変異体よりは強かったが野生株に比べるとかなり弱かった.このような結果から,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーは通常の2次細胞壁とは明らかに異なる細胞壁を形成させると考えられた.この結果は,VP16ドメインを付加しないクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させた場合も同様であった.
クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体のもつ,通常の2次細胞壁とは明らかに異なる細胞壁の正体を明らかにするため,RNAを抽出してマイクロアレイ法により解析したところ,nst1 nst3二重変異体と比べ,2次細胞壁の形成にかかわることの知られている遺伝子のほとんどについて,発現は上昇していなかった.それに対し,細胞壁に関連する遺伝子のなかでも1次細胞壁の形成や再編成に関与するとされる遺伝子の発現は上昇していた.とくに示唆的であったのは,セルロース合成酵素の遺伝子の発現パターンであった.セルロース合成酵素の遺伝子は,1次細胞壁においてセルロースの合成を制御する遺伝子と,2次細胞壁においてセルロースの合成を制御する遺伝子とに完全に分かれていることが知られているが,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体においては,1次細胞壁においてセルロースの合成を制御する遺伝子のみ発現が上昇していた.また,細胞壁を硫酸により加水分解して得られる単糖の組成比を分析したところ,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体にはガラクツロン酸,ラムノース,ガラクトースなど1次細胞壁に多い単糖が多く含まれており,ほとんど1次細胞壁しかもたないnst1 nst3二重変異体の組成比と非常に類似していた.また,茎の切片の抗体染色により,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体の肥厚した細胞壁には,通常の2次細胞壁によくみられるキシランがなく,ペクチンに富むことがわかった.これらの結果から,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体の肥厚した細胞壁は,1次細胞壁にきわめて類似した特徴をもつと考えられた(図2).
クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーが一般的に1次細胞壁の形成を制御するのかどうかを明らかにするため,プロトプラストからの1次細胞壁の再生の過程において,VP16ドメインあるいはSRDXドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーの過剰な発現のおよぼす影響について調べた.その結果,VP16ドメインを付加あるいはなにも付加しないクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを過剰に発現させた場合にはプロトプラストからの1次細胞壁の再生が早まり,逆に,SRDXドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを過剰に発現させた場合にはプロトプラストからの1次細胞壁の再生が遅れた.この結果から,クラスIIIdおよびクラスIIIeのERF転写因子ファミリーは1次細胞壁の形成においてマスター転写因子として機能することが示唆された.クラスIIIdおよびクラスIIIeのERF転写因子ファミリーには10個の転写因子が含まれるが,それぞれの遺伝子のプロモーター活性について調べたところ,さまざまな部位において活性が認められた.このようなことから,このクラスIIIdおよびクラスIIIeのERF転写因子ファミリーはさまざまな部位において1次細胞壁の形成を制御するマスター転写因子であると考えられた.
この研究の意義は,つぎの2点に集約される.すなわち,1)ほとんど未知であった植物の1次細胞壁の形成を制御する転写因子を同定したこと,2)多くの細胞において通常は肥厚しない1次細胞壁を2次細胞壁のように肥厚させたこと,である.1)についての今後の課題のひとつは,クラスIIIdおよびクラスIIIeのERF転写因子ファミリーの10個の転写因子それぞれがどのような生物学的な機能をはたすかである.これについては,ゲノム編集技術などを駆使して丹念に変異体,とくに多重変異体を作製することでしか明らかにされないであろうが,植物における1次細胞壁の重要性をかんがみると,変異体が致死的な表現型を示すことも想定される.また別の課題は,1次細胞壁の形成を制御する転写制御カスケードがどうなっているかである.2次細胞壁の形成を制御する転写制御カスケードはNST転写因子ファミリーおよびVND転写因子ファミリーを頂点とした多層構造をとることが明らかにされているが,1次細胞壁の形成の場合はどうだろうか.さらなる将来の展望のひとつは,細胞分裂あるいは細胞伸長との関係である.1次細胞壁は細胞分裂とそののちの細胞伸長にあたり新生および再編成を必要とするが,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーがどのように関係するかは興味深い.また,1次細胞壁を制御することにより産業あるいは農業への応用につながるかという点も重要である.2)についての今後の課題は,nst1 nst3二重変異体に肥厚した1次細胞壁に近い細胞壁のさらなる性質の解明および分子機構の理解である.たとえば,この1次細胞壁様の細胞壁に含まれるセルロースは,重合度および結晶化度が通常の1次細胞壁にみられるセルロースより高くなる傾向があった.通常は肥厚しない1次細胞壁がここまで肥厚することにより,セルロースの性質が2次細胞壁のセルロースに近づいている可能性がある.さらなる将来の展望としては,この肥厚した1次細胞壁様の細胞壁にリグニンやキシランを追加することにより2次細胞壁の性質に近い細胞壁が形成されるかどうかという点に興味がもたれる.リグニンの生合成を制御する転写因子はすでに明らかにされているので7),技術的には可能ではないかと思われる.2次細胞壁におけるリグニンやキシランの意義については,さまざまなことが言及されているが,証拠をもとに明確に議論された例はほとんどないかと思われる.クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体が,こうした疑問への回答を与えるツールとなることを期待している.
ほとんど蛇足かもしれないが,nst1 nst3二重変異体に形成された1次細胞壁に近い肥厚した細胞壁をなんとよぶべきだろうか(図2).本来の定義としては,細胞が伸長しているときに形成された細胞壁が1次細胞壁で,成長の停止した一部の細胞に2次的に形成された細胞壁が2次細胞壁である.その定義にしたがえば,nst1 nst3二重変異体に形成された肥厚した細胞壁は依然として2次細胞壁とよぶべきものであるが,その性質は明らかに1次細胞壁であり,2次細胞壁とよぶのははばかられる.語句の使い方は本質的な問題ではないが,ある意味,この研究の成果はこれまでの概念をくつがえすようなものであるといえ,古典的な分類では定義がむずかしいほかの事象ともあわせて,細胞壁の分類および定義を再考する必要があるかもしれない.
略歴:2013年 広島大学大学院生物圏科学研究科にて博士号取得,同年より産業技術総合研究所生物プロセス研究部門 研究員.
研究テーマ:植物の細胞壁に関する基礎的および応用的な研究.
光田 展隆(Nobutaka Mitsuda)
産業技術総合研究所生物プロセス研究部門 グループ長.
研究室URL:https://unit.aist.go.jp/bpri/bpri-pgrr/
© 2018 坂本真吾・光田展隆 Licensed under CC 表示 2.1 日本
(産業技術総合研究所生物プロセス研究部門 植物機能制御研究グループ)
email:光田展隆
DOI: 10.7875/first.author.2018.110
Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis.
Shingo Sakamoto, Marc Somssich, Miyuki T. Nakata, Faride Unda, Kimie Atsuzawa, Yasuko Kaneko, Ting Wang, Anne-Maarit Bågman, Allison Gaudinier, Kouki Yoshida, Siobhan M. Brady, Shawn D. Mansfield, Staffan Persson, Nobutaka Mitsuda
Nature Plants, 4, 777-783 (2018)
要 約
植物の1次細胞壁は植物にとり必須の構造であるが,これまで,その形成を制御する転写因子についてはあまりよくわかっていない.この研究において,筆者らは,シロイヌナズナを用いて,茎の繊維細胞に2次細胞壁の形成されないnst1 nst3二重変異体にさまざまな転写因子を繊維細胞に特異的に発現させることにより,1次細胞壁を肥厚させる転写因子としてクラスIIIdおよびクラスIIIeのERF転写因子ファミリーを同定した.繊維細胞にクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体においては,2次細胞壁をもたない茎の繊維細胞に1次細胞壁のような特徴をもつ細胞壁が形成された.すなわち,2次細胞壁を1次細胞壁に置き換えたような植物になった.クラスIIIdおよびクラスIIIeのERF転写因子ファミリーはさまざまな部位にあまり重なることなく発現しており,全体として1次細胞壁の形成を制御すると考えられた.1次細胞壁は,セルロースが酵素により糖化されやすいという特長にくわえ,果実の食感や保存性を左右するほか,細胞へのマイクロインジェクションにおいて障害になるなど,植物バイオテクノロジーにとり重要な要素であり,今回の発見が今後の研究開発に貢献することが期待される.
はじめに
植物の細胞壁は1次細胞壁と2次細胞壁に大別される.1次細胞壁は比較的薄くすべての細胞がもち,植物の細胞を特徴づける構造のひとつである.2次細胞壁は茎など強度の必要な組織の繊維細胞,水分を全身に供給するため水圧に耐える必要のある道管,自然に開裂するため張力を蓄える必要のある葯や果実莢の内皮細胞に形成される,厚い細胞壁である.どちらも細胞膜の外側に形成されるが,2次細胞壁は1次細胞壁と細胞膜のあいだに形成される.すなわち,かならず1次細胞壁,2次細胞壁の順に形成され,一般には,細胞が伸長しているときに再編成をくり返しながら形成されるのが1次細胞壁で,伸長が停止した細胞において形成されるのが2次細胞壁と定義される.構成成分の観点からは,1次細胞壁はおもにセルロース,ヘミセルロース,ペクチンから構成されるが,2次細胞壁はおもにセルロース,ヘミセルロース,リグニンから構成される.どちらにもヘミセルロースが含まれるが,ヘミセルロースはアルカリに可溶性の鎖状ヘテロ多糖類の総称であり,1次細胞壁のヘミセルロースはキシログルカンに富み,2次細胞壁のヘミセルロースはキシラン(あるいは,グルコマンナン)に富むといった特徴がある.また,1次細胞壁のみにみられるペクチンは酸に可溶性の鎖状ヘテロ多糖類であり,構造的にはヘミセルロースに近い.セルロースはどちらにも含まれるが,1次細胞壁と2次細胞壁とでは重合度や結晶化度が異なる.リグニンはおもに2次細胞壁に沈着しているが,一部の1次細胞壁にも沈着がみられる.
これまで,それぞれの構成成分の合成にかかわる酵素の同定が進んできたほか,2次細胞壁に関しては,形成を遺伝子発現のレベルで制御する転写因子が多く同定されてきた.なかでも,マスター転写因子と考えられているのがVND転写因子ファミリー1,2) およびNST転写因子ファミリー3,4) である.これらの転写因子ファミリーは植物に特異的な巨大な転写因子ファミリーであるNAC転写因子ファミリーに属するが,同じサブファミリーに属しており相同性は高い.VND転写因子ファミリーはシロイヌナズナにおいて道管の形成を制御しており,道管において2次細胞壁の形成からプログラム細胞死の制御までを担う1,2).NST転写因子ファミリーはシロイヌナズナにおいて茎の繊維細胞3),葯4),果実莢5) の内皮細胞において2次細胞壁の形成を制御しており,nst1 nst3二重変異体においては茎や胚軸の繊維細胞における2次細胞壁の形成が完全に失われる3)(図1).植物において2次細胞壁は一般に乾燥重量の過半をしめる巨大な代謝産物であることから,筆者らは,nst1 nst3二重変異体を器だけあって中身のない植物ととらえ,繊維細胞に特異的にさまざまな転写因子を発現させることにより,新しい細胞壁を形成するような転写因子を探索した6)(図1).
1.クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーは通常の2次細胞壁とは異なる細胞壁を形成させる
茎の繊維細胞において2次細胞壁の形成が失われたnst1 nst3変異体は,一定の高さまで茎が成長すると自重に耐えきれず倒れるという表現型を示す3)(図1).また,茎は非常に折れやすく手で曲げることにより野生株との違いがはっきりとわかる.シロイヌナズナのもつすべての転写因子約1900個からおのおののサブファミリーを代表する転写因子をひとつずつ選出し,合計で約300個の転写因子をスクリーニングの対象とした.これらに転写活性化ドメインであるVP16ドメインあるいは転写抑制化ドメインであるSRDXドメインを付加し,約30個ずつまとめて繊維細胞に特異的な遺伝子発現を誘導するNST3プロモーターによりnst1 nst3変異体に発現させ,手曲げ試験や徒手切片の観察によりスクリーニングを実施した6).その結果,さまざまな転写因子が表現型を部分的に回復させ,その多くでは2次細胞壁の形成が部分的に回復していた6).しかし,VP16ドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させた場合には,細胞壁が肥厚しているにもかかわらず,通常の2次細胞壁にみられるリグニンの沈着はまったくみられなかった.この肥厚した細胞壁においては,リグニンがないだけでなく,通常の2次細胞壁にみられるキシランも確認されなかった.透過型電子顕微鏡により超微細構造を観察したところ,通常の2次細胞壁に近い細胞壁の形成がみられたものの,細胞壁は層状の構造をとるようにみえたほか,細胞壁の輪郭がなめらかでないようにみえた.茎の強度もnst1 nst3二重変異体よりは強かったが野生株に比べるとかなり弱かった.このような結果から,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーは通常の2次細胞壁とは明らかに異なる細胞壁を形成させると考えられた.この結果は,VP16ドメインを付加しないクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させた場合も同様であった.
2.クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーにより形成される細胞壁は1次細胞壁のような特徴をもつ
クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体のもつ,通常の2次細胞壁とは明らかに異なる細胞壁の正体を明らかにするため,RNAを抽出してマイクロアレイ法により解析したところ,nst1 nst3二重変異体と比べ,2次細胞壁の形成にかかわることの知られている遺伝子のほとんどについて,発現は上昇していなかった.それに対し,細胞壁に関連する遺伝子のなかでも1次細胞壁の形成や再編成に関与するとされる遺伝子の発現は上昇していた.とくに示唆的であったのは,セルロース合成酵素の遺伝子の発現パターンであった.セルロース合成酵素の遺伝子は,1次細胞壁においてセルロースの合成を制御する遺伝子と,2次細胞壁においてセルロースの合成を制御する遺伝子とに完全に分かれていることが知られているが,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体においては,1次細胞壁においてセルロースの合成を制御する遺伝子のみ発現が上昇していた.また,細胞壁を硫酸により加水分解して得られる単糖の組成比を分析したところ,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体にはガラクツロン酸,ラムノース,ガラクトースなど1次細胞壁に多い単糖が多く含まれており,ほとんど1次細胞壁しかもたないnst1 nst3二重変異体の組成比と非常に類似していた.また,茎の切片の抗体染色により,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体の肥厚した細胞壁には,通常の2次細胞壁によくみられるキシランがなく,ペクチンに富むことがわかった.これらの結果から,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体の肥厚した細胞壁は,1次細胞壁にきわめて類似した特徴をもつと考えられた(図2).
3.クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーはプロトプラストからの細胞壁の再生を制御する
クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーが一般的に1次細胞壁の形成を制御するのかどうかを明らかにするため,プロトプラストからの1次細胞壁の再生の過程において,VP16ドメインあるいはSRDXドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーの過剰な発現のおよぼす影響について調べた.その結果,VP16ドメインを付加あるいはなにも付加しないクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを過剰に発現させた場合にはプロトプラストからの1次細胞壁の再生が早まり,逆に,SRDXドメインを付加したクラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを過剰に発現させた場合にはプロトプラストからの1次細胞壁の再生が遅れた.この結果から,クラスIIIdおよびクラスIIIeのERF転写因子ファミリーは1次細胞壁の形成においてマスター転写因子として機能することが示唆された.クラスIIIdおよびクラスIIIeのERF転写因子ファミリーには10個の転写因子が含まれるが,それぞれの遺伝子のプロモーター活性について調べたところ,さまざまな部位において活性が認められた.このようなことから,このクラスIIIdおよびクラスIIIeのERF転写因子ファミリーはさまざまな部位において1次細胞壁の形成を制御するマスター転写因子であると考えられた.
おわりに
この研究の意義は,つぎの2点に集約される.すなわち,1)ほとんど未知であった植物の1次細胞壁の形成を制御する転写因子を同定したこと,2)多くの細胞において通常は肥厚しない1次細胞壁を2次細胞壁のように肥厚させたこと,である.1)についての今後の課題のひとつは,クラスIIIdおよびクラスIIIeのERF転写因子ファミリーの10個の転写因子それぞれがどのような生物学的な機能をはたすかである.これについては,ゲノム編集技術などを駆使して丹念に変異体,とくに多重変異体を作製することでしか明らかにされないであろうが,植物における1次細胞壁の重要性をかんがみると,変異体が致死的な表現型を示すことも想定される.また別の課題は,1次細胞壁の形成を制御する転写制御カスケードがどうなっているかである.2次細胞壁の形成を制御する転写制御カスケードはNST転写因子ファミリーおよびVND転写因子ファミリーを頂点とした多層構造をとることが明らかにされているが,1次細胞壁の形成の場合はどうだろうか.さらなる将来の展望のひとつは,細胞分裂あるいは細胞伸長との関係である.1次細胞壁は細胞分裂とそののちの細胞伸長にあたり新生および再編成を必要とするが,クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーがどのように関係するかは興味深い.また,1次細胞壁を制御することにより産業あるいは農業への応用につながるかという点も重要である.2)についての今後の課題は,nst1 nst3二重変異体に肥厚した1次細胞壁に近い細胞壁のさらなる性質の解明および分子機構の理解である.たとえば,この1次細胞壁様の細胞壁に含まれるセルロースは,重合度および結晶化度が通常の1次細胞壁にみられるセルロースより高くなる傾向があった.通常は肥厚しない1次細胞壁がここまで肥厚することにより,セルロースの性質が2次細胞壁のセルロースに近づいている可能性がある.さらなる将来の展望としては,この肥厚した1次細胞壁様の細胞壁にリグニンやキシランを追加することにより2次細胞壁の性質に近い細胞壁が形成されるかどうかという点に興味がもたれる.リグニンの生合成を制御する転写因子はすでに明らかにされているので7),技術的には可能ではないかと思われる.2次細胞壁におけるリグニンやキシランの意義については,さまざまなことが言及されているが,証拠をもとに明確に議論された例はほとんどないかと思われる.クラスIIIdあるいはクラスIIIeのERF転写因子ファミリーを発現させたnst1 nst3二重変異体が,こうした疑問への回答を与えるツールとなることを期待している.
ほとんど蛇足かもしれないが,nst1 nst3二重変異体に形成された1次細胞壁に近い肥厚した細胞壁をなんとよぶべきだろうか(図2).本来の定義としては,細胞が伸長しているときに形成された細胞壁が1次細胞壁で,成長の停止した一部の細胞に2次的に形成された細胞壁が2次細胞壁である.その定義にしたがえば,nst1 nst3二重変異体に形成された肥厚した細胞壁は依然として2次細胞壁とよぶべきものであるが,その性質は明らかに1次細胞壁であり,2次細胞壁とよぶのははばかられる.語句の使い方は本質的な問題ではないが,ある意味,この研究の成果はこれまでの概念をくつがえすようなものであるといえ,古典的な分類では定義がむずかしいほかの事象ともあわせて,細胞壁の分類および定義を再考する必要があるかもしれない.
文 献
- Kubo, M., Udagawa, M., Nishikubo, N. et al.: Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev., 19, 1855-1860 (2005)[PubMed]
- Tan, T. T., Endo, H., Sano, R. et al.: Transcription factors VND1-VND3 contribute to cotyledon xylem vessel formation. Plant Physiol., 176, 773-789 (2018)[PubMed]
- Mitsuda, N., Iwase, A., Yamamoto, H. et al.: NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell, 19, 270-280 (2007)[PubMed]
- Mitsuda, N., Seki, M., Shinozaki, K. et al.: The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell, 17, 2993-3006 (2005)[PubMed]
- Mitsuda, N. & Ohme-Takagi, M.: NAC transcription factors NST1 and NST3 regulate pod shattering in a partially redundant manner by promoting secondary wall formation after the establishment of tissue identity. Plant J., 56, 768-778 (2008)[PubMed]
- Sakamoto, S. & Mitsuda, N.: Reconstitution of a secondary cell wall in a secondary cell wall-deficient Arabidopsis mutant. Plant Cell Physiol., 56, 299-310 (2015)[PubMed]
- Zhou, J., Lee, C., Zhong, R. et al.: MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabidopsis. Plant Cell, 21, 248-266 (2009)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2013年 広島大学大学院生物圏科学研究科にて博士号取得,同年より産業技術総合研究所生物プロセス研究部門 研究員.
研究テーマ:植物の細胞壁に関する基礎的および応用的な研究.
光田 展隆(Nobutaka Mitsuda)
産業技術総合研究所生物プロセス研究部門 グループ長.
研究室URL:https://unit.aist.go.jp/bpri/bpri-pgrr/
© 2018 坂本真吾・光田展隆 Licensed under CC 表示 2.1 日本
