ラットの海馬における“自己の場所”と“他者の場所”の表象
檀上輝子・藤澤茂義
(理化学研究所脳科学総合研究センター システム神経生理学研究チーム)
email:藤澤茂義
DOI: 10.7875/first.author.2018.015
Spatial representations of self and other in the hippocampus.
Teruko Danjo, Taro Toyoizumi, Shigeyoshi Fujisawa
Science, 359, 213-218 (2018)
動物が自己の存在する場所を認知するには海馬の機能が必須である.しかしながら,これまで,他者がどこに存在するか認知する機構は不明であった.この研究において,筆者らは,他者の観察を必要とする行動実験により,他者の空間位置の情報が海馬においてどのように表象されるかを解析し,これまで,自己の空間位置を表象するとされてきた場所細胞が,他者の空間位置をも同時に表象する“ジョイント場所細胞”であることを明らかにした.さらに,自己が存在する場合でも他者が存在する場合でも反応する“共通場所細胞”の存在も見い出された.この結果により,海馬において表象される空間地図には,自己のみでなく,他者という異なる次元の空間事象も表象されることが示された.
ヒトをはじめとする高等な哺乳動物は客観的(あるいは,他者中心的)な空間の認知が可能である.このような空間の認知を可能にする神経基盤として,海馬や嗅内皮質を含む神経ネットワークに認識地図が存在することが提唱された1,2).認識地図の代表的な構成要素として場所細胞があげられる.場所細胞は海馬のCA1野およびCA3野に多く存在する錐体細胞であり,空間の特定の場所に場所受容野をもち,自己がその場所受容野にいるとき特異的に発火する.場所細胞の発火のパターンは環境の変化により柔軟に変化するため,たんに自己中心的な自己の空間位置を表象するのではなく,他者中心的な空間位置を表象すると考えられる3).そのほか,嗅内皮質に存在する頭位方向細胞やグリッド細胞なども認識地図の構成要素であり,これまで,自己が存在する場所の位置情報がこれらの神経細胞によりどのようにコードされるか,多くの研究がなされてきた.他方,客観的な空間の認知には自己が存在する以外の場所,たとえば,なにかの目印になる場所や動きまわる物体の空間位置をもコードすることが必要である.しかし,これまで,どのような機構により自己以外の位置情報が脳において表象されるのかは不明であった.この研究は,自己以外の例として,“他者”の位置情報が海馬においてどのように表象されるかを明らかにすることを目的とした.
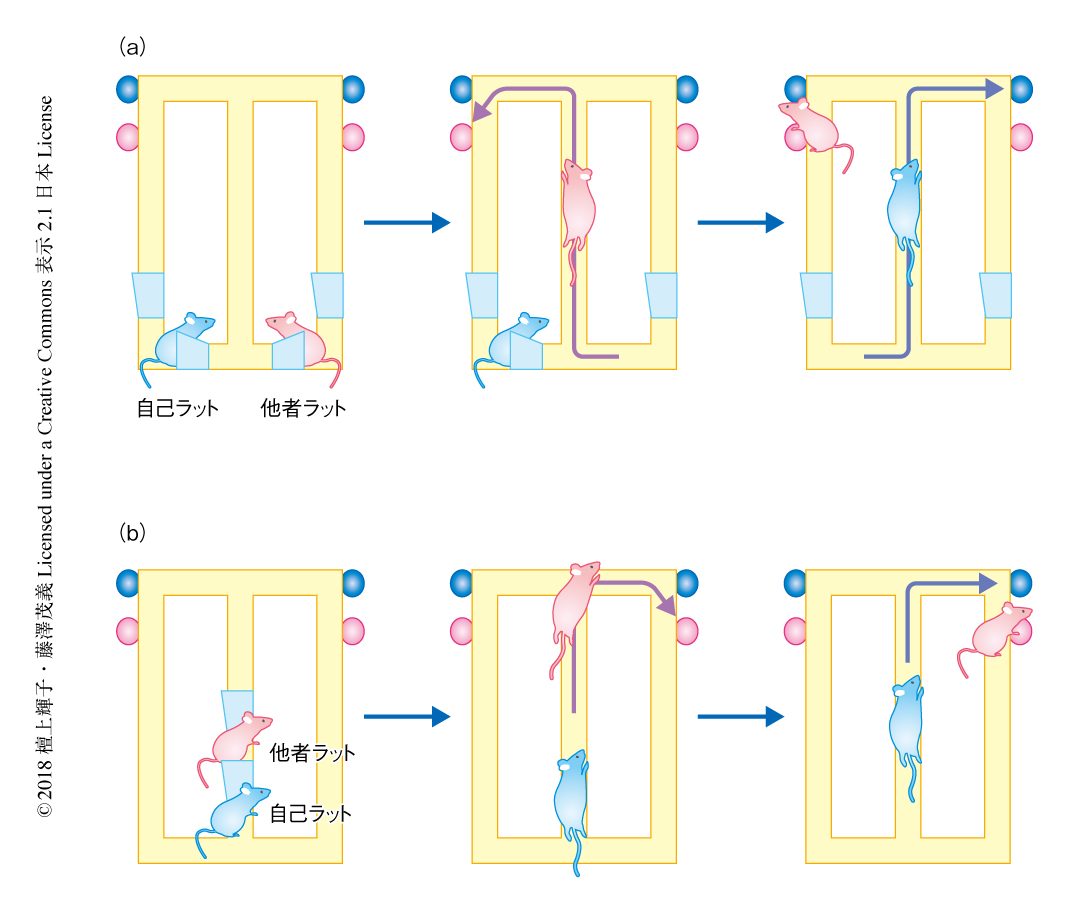
この目的のため,他者の観察を必要とする行動課題を開発し,他者を観察しているラットの海馬から神経活動を記録し,他者の位置情報がどのように表象されるかを解析した.他者の観察を必要とする行動課題は,T字型迷路および自己と他者の2匹のラットを用い,1匹目の他者ラットがさきにスタートし,2匹目の自己ラットがそれを追いかけるかたちで開始する.行動課題には2種類あり,反対方向課題(図1a)では2匹目の自己ラットが他者ラットと反対の方向を選択すると報酬の水が得られ,同一方向課題(図1b)では自己ラットが他者ラットと同じ方向を選択すると報酬が得られる.したがって,この行動課題においては,自己ラットがどこに他者ラットがいるのかを認識することが必須である.
自己ラットの海馬の背側のCA1野に64チャネルのシリコンプローブを挿入し神経活動を記録した.神経活動の記録データからスパイクをソートすることにより単一ユニットを分離し,スパイクの波形の形状から錐体細胞と介在ニューロンとに分類し,錐体細胞の単一のユニットにおける発火のタイミングと行動記録(自己ラットおよび他者ラットの位置情報)とをつきあわせることにより,他者ラットの位置情報が自己ラットの海馬においてどのように表象されるか解析した.
反対方向課題の記録データから自己の場所に特異的に発火する,いわゆる,場所細胞を同定した.行動課題の開始地点から報酬地点までの空間に場所受容野をもつ1298個の場所細胞が同定され,この場所細胞に限定して解析を進めた.場所細胞が発火したときに自己ラットがT字型迷路のどこに存在したのか,場所-発火頻度のマップ(自己マップ)を作成した.さらに,これら自己ラットの場所細胞が発火したときに他者ラットがT字型迷路のどこに存在したかを明らかにし,自己マップと同様に,場所-発火頻度マップ(他者マップ)を作成した.多くの場所細胞において,他者マップにもはっきりとした場所受容野が認められた.
この他者マップにおける場所受容野は,自己と他者の空間位置の関係の拘束性に依存するものではなく,実際に他者の場所に特異的に発火したものなのかどうかを解析した.そのための帰無仮説として,場所細胞の発火が完全に自己の場所に依存すると仮定し,この仮定にしたがう対照のデータを実際のデータをリサンプリングすることにより作成し,実際のデータを対照のデータと比較することにより帰無仮説が棄却されるかどうか検定した4).具体的には,他者の場所に対する発火の頻度を対照のデータにおいても計算し,上位5%の値を閾値としてこれを実際のデータがこえた場合,その場所を他者に有意に発火する領域とした.85%の場所細胞においてこのような有意な領域が認められたため帰無仮説は棄却され,場所細胞の多くが他者の場所に特異的に発火することが明らかにされた.他者の場所に対する有意な領域は一様に分布し,自己あるいは他者の特別な行動や場所に依存するものではないことがわかった.同一方向課題においても同様の結果が得られ,また,多くの錐体細胞は,課題のスタートから経過した時間に依存して発火する,いわゆる,時間細胞5) ではないことも示された.以上の結果から,海馬に存在する錐体細胞の多くは自己の場所と他者の場所を同時に表象すると結論し,この性質をもつ場所細胞を“ジョイント場所細胞”と命名した(図2a).
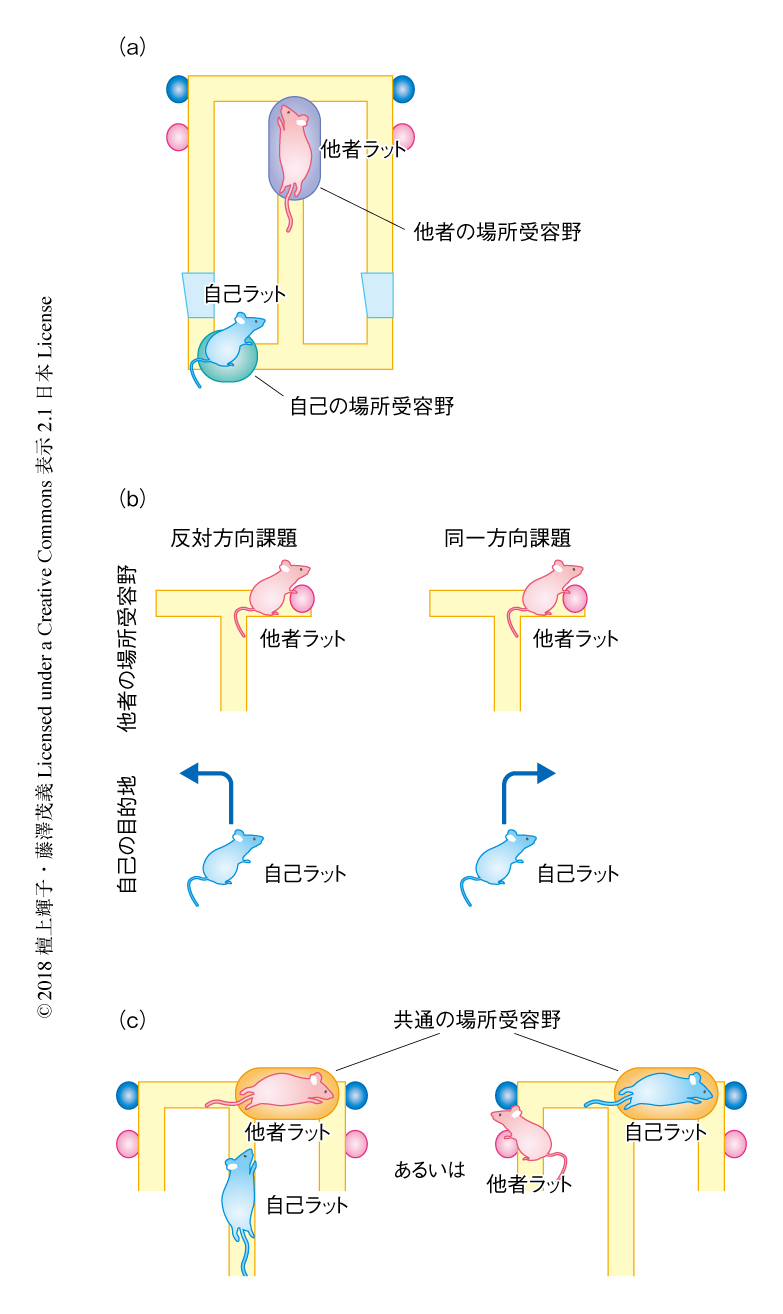
反対方向課題と同一方向課題とでは,他者ラットの場所と自己ラットの目的地との関係性が異なる.他者ラットが右側にいるとき,自己ラットの目的地は反対方向課題では左側だが同一方向課題では右側になる(図2b).この行動課題の特性をいかして,他者の空間位置を特異的にコードする“他者の場所細胞”が13%(58個),自己の目的地を特異的にコードする“ゴール細胞”6) が58%(252個)同定された.また,他者の場所細胞の場所受容野はT字型迷路において左右のサイドアームの全体に分布した.これらの結果から,この他者の観察を必要とする行動課題においては,自己の目的地と他者の場所が独立して表象されることが示された.このような他者の場所に特異的に発火する場所細胞は,この研究においてはじめて同定されたものである.
共通場所細胞は,場所受容野に自己が存在するときも他者が存在するときも発火する場所細胞であり(図2c),自己マップと他者マップが共通の場所受容野をもつため,両者の要素積においても同じ場所にピークが認められる.このような発火のパターンをもつ共通場所細胞が合計で46個同定され,その場所受容野はT字型迷路の全域に分布することが確認された.この共通場所細胞は自己の場所と他者の場所に同じように発火することから,サルの前頭葉においてはじめに報告された,自己の動作と他者の動作に同じように発火するミラー細胞7) と同様の特徴をもつものであった.
ある時間tにおいて,自己の空間位置は,その時間tにおける個々の場所細胞の発火の回数からベイズ統計により推定することが可能であると報告されている8).この研究の記録データからも,自己の場所を15 cm程度の誤差で再構築することが可能であった.さらに,同様の手法を用いて,自己の場所を再構築したのと同じ場所細胞アセンブリの発火の回数を用いたところ,他者の場所も20 cm程度の誤差で再構築された.
さらに,反対方向課題と同一方向課題の両方のデータをあわせて他者の場所を再構築した場合と,再構築した自己の場所と自己ラットと他者ラットとの位置関係の時間分布から他者の場所を推定した場合とを比較した.その結果,前者のほうが誤差は小さく,場所細胞アセンブリの発火のパターンのほうが,自己ラットと他者ラットとの位置関係よりも有意に詳細な他者の場所情報を含むことが明らかにされた.
この研究においては,他者の観察を必要とする行動課題を遂行しているラットの海馬のCA1野において,他者の位置情報がどのように表象されるかを解析することにより,場所細胞の多くが自己の場所と他者の場所を同時に表象するジョイント場所細胞であることが明らかにされた.自己のゴールとは独立して他者の場所を表象する他者の場所細胞,および,自己あるいは他者にかかわらず反応する場所受容野をもつ共通場所細胞が同定され,さらに,他者の空間位置を場所細胞の発火のパターンから再構築できることが示された.この研究により,従来は自己の場所をコードすると考えられていた場所細胞が,他者の場所という異なる次元の空間事象をも表象することがはじめて示された.
動物の社会的な行動においては,その前提として,他者の場所や行動を認識する必要があると考えられる.今回,海馬が他者の位置情報を表象することが明らかにされたことから,海馬や脳のほかの領域が動物の社会性9,10) に関してどのように機能するのか,今後,さらに解明が進むことが期待される.
略歴:2011年 京都大学大学院医学研究科 修了,同年 大阪バイオサイエンス研究所 研究員を経て,2013年より理化学研究所脳科学総合研究センター 研究員.
藤澤 茂義(Shigeyoshi Fujisawa)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://fujisawalab.brain.riken.jp/aboutjp.html
© 2018 檀上輝子・藤澤茂義 Licensed under CC 表示 2.1 日本
(理化学研究所脳科学総合研究センター システム神経生理学研究チーム)
email:藤澤茂義
DOI: 10.7875/first.author.2018.015
Spatial representations of self and other in the hippocampus.
Teruko Danjo, Taro Toyoizumi, Shigeyoshi Fujisawa
Science, 359, 213-218 (2018)
要 約
動物が自己の存在する場所を認知するには海馬の機能が必須である.しかしながら,これまで,他者がどこに存在するか認知する機構は不明であった.この研究において,筆者らは,他者の観察を必要とする行動実験により,他者の空間位置の情報が海馬においてどのように表象されるかを解析し,これまで,自己の空間位置を表象するとされてきた場所細胞が,他者の空間位置をも同時に表象する“ジョイント場所細胞”であることを明らかにした.さらに,自己が存在する場合でも他者が存在する場合でも反応する“共通場所細胞”の存在も見い出された.この結果により,海馬において表象される空間地図には,自己のみでなく,他者という異なる次元の空間事象も表象されることが示された.
はじめに
ヒトをはじめとする高等な哺乳動物は客観的(あるいは,他者中心的)な空間の認知が可能である.このような空間の認知を可能にする神経基盤として,海馬や嗅内皮質を含む神経ネットワークに認識地図が存在することが提唱された1,2).認識地図の代表的な構成要素として場所細胞があげられる.場所細胞は海馬のCA1野およびCA3野に多く存在する錐体細胞であり,空間の特定の場所に場所受容野をもち,自己がその場所受容野にいるとき特異的に発火する.場所細胞の発火のパターンは環境の変化により柔軟に変化するため,たんに自己中心的な自己の空間位置を表象するのではなく,他者中心的な空間位置を表象すると考えられる3).そのほか,嗅内皮質に存在する頭位方向細胞やグリッド細胞なども認識地図の構成要素であり,これまで,自己が存在する場所の位置情報がこれらの神経細胞によりどのようにコードされるか,多くの研究がなされてきた.他方,客観的な空間の認知には自己が存在する以外の場所,たとえば,なにかの目印になる場所や動きまわる物体の空間位置をもコードすることが必要である.しかし,これまで,どのような機構により自己以外の位置情報が脳において表象されるのかは不明であった.この研究は,自己以外の例として,“他者”の位置情報が海馬においてどのように表象されるかを明らかにすることを目的とした.
この目的のため,他者の観察を必要とする行動課題を開発し,他者を観察しているラットの海馬から神経活動を記録し,他者の位置情報がどのように表象されるかを解析した.他者の観察を必要とする行動課題は,T字型迷路および自己と他者の2匹のラットを用い,1匹目の他者ラットがさきにスタートし,2匹目の自己ラットがそれを追いかけるかたちで開始する.行動課題には2種類あり,反対方向課題(図1a)では2匹目の自己ラットが他者ラットと反対の方向を選択すると報酬の水が得られ,同一方向課題(図1b)では自己ラットが他者ラットと同じ方向を選択すると報酬が得られる.したがって,この行動課題においては,自己ラットがどこに他者ラットがいるのかを認識することが必須である.
自己ラットの海馬の背側のCA1野に64チャネルのシリコンプローブを挿入し神経活動を記録した.神経活動の記録データからスパイクをソートすることにより単一ユニットを分離し,スパイクの波形の形状から錐体細胞と介在ニューロンとに分類し,錐体細胞の単一のユニットにおける発火のタイミングと行動記録(自己ラットおよび他者ラットの位置情報)とをつきあわせることにより,他者ラットの位置情報が自己ラットの海馬においてどのように表象されるか解析した.
1.場所細胞の多くは自己の場所と他者の場所を同時に表象するジョイント場所細胞である
反対方向課題の記録データから自己の場所に特異的に発火する,いわゆる,場所細胞を同定した.行動課題の開始地点から報酬地点までの空間に場所受容野をもつ1298個の場所細胞が同定され,この場所細胞に限定して解析を進めた.場所細胞が発火したときに自己ラットがT字型迷路のどこに存在したのか,場所-発火頻度のマップ(自己マップ)を作成した.さらに,これら自己ラットの場所細胞が発火したときに他者ラットがT字型迷路のどこに存在したかを明らかにし,自己マップと同様に,場所-発火頻度マップ(他者マップ)を作成した.多くの場所細胞において,他者マップにもはっきりとした場所受容野が認められた.
この他者マップにおける場所受容野は,自己と他者の空間位置の関係の拘束性に依存するものではなく,実際に他者の場所に特異的に発火したものなのかどうかを解析した.そのための帰無仮説として,場所細胞の発火が完全に自己の場所に依存すると仮定し,この仮定にしたがう対照のデータを実際のデータをリサンプリングすることにより作成し,実際のデータを対照のデータと比較することにより帰無仮説が棄却されるかどうか検定した4).具体的には,他者の場所に対する発火の頻度を対照のデータにおいても計算し,上位5%の値を閾値としてこれを実際のデータがこえた場合,その場所を他者に有意に発火する領域とした.85%の場所細胞においてこのような有意な領域が認められたため帰無仮説は棄却され,場所細胞の多くが他者の場所に特異的に発火することが明らかにされた.他者の場所に対する有意な領域は一様に分布し,自己あるいは他者の特別な行動や場所に依存するものではないことがわかった.同一方向課題においても同様の結果が得られ,また,多くの錐体細胞は,課題のスタートから経過した時間に依存して発火する,いわゆる,時間細胞5) ではないことも示された.以上の結果から,海馬に存在する錐体細胞の多くは自己の場所と他者の場所を同時に表象すると結論し,この性質をもつ場所細胞を“ジョイント場所細胞”と命名した(図2a).
2.他者の場所をコードする他者の場所細胞と自己の目的地をコードするゴール細胞
反対方向課題と同一方向課題とでは,他者ラットの場所と自己ラットの目的地との関係性が異なる.他者ラットが右側にいるとき,自己ラットの目的地は反対方向課題では左側だが同一方向課題では右側になる(図2b).この行動課題の特性をいかして,他者の空間位置を特異的にコードする“他者の場所細胞”が13%(58個),自己の目的地を特異的にコードする“ゴール細胞”6) が58%(252個)同定された.また,他者の場所細胞の場所受容野はT字型迷路において左右のサイドアームの全体に分布した.これらの結果から,この他者の観察を必要とする行動課題においては,自己の目的地と他者の場所が独立して表象されることが示された.このような他者の場所に特異的に発火する場所細胞は,この研究においてはじめて同定されたものである.
3.自己と他者とで共通の場所受容野をもつ共通場所細胞
共通場所細胞は,場所受容野に自己が存在するときも他者が存在するときも発火する場所細胞であり(図2c),自己マップと他者マップが共通の場所受容野をもつため,両者の要素積においても同じ場所にピークが認められる.このような発火のパターンをもつ共通場所細胞が合計で46個同定され,その場所受容野はT字型迷路の全域に分布することが確認された.この共通場所細胞は自己の場所と他者の場所に同じように発火することから,サルの前頭葉においてはじめに報告された,自己の動作と他者の動作に同じように発火するミラー細胞7) と同様の特徴をもつものであった.
4.他者の場所も場所細胞の発火のパターンから再構築することが可能である
ある時間tにおいて,自己の空間位置は,その時間tにおける個々の場所細胞の発火の回数からベイズ統計により推定することが可能であると報告されている8).この研究の記録データからも,自己の場所を15 cm程度の誤差で再構築することが可能であった.さらに,同様の手法を用いて,自己の場所を再構築したのと同じ場所細胞アセンブリの発火の回数を用いたところ,他者の場所も20 cm程度の誤差で再構築された.
さらに,反対方向課題と同一方向課題の両方のデータをあわせて他者の場所を再構築した場合と,再構築した自己の場所と自己ラットと他者ラットとの位置関係の時間分布から他者の場所を推定した場合とを比較した.その結果,前者のほうが誤差は小さく,場所細胞アセンブリの発火のパターンのほうが,自己ラットと他者ラットとの位置関係よりも有意に詳細な他者の場所情報を含むことが明らかにされた.
おわりに
この研究においては,他者の観察を必要とする行動課題を遂行しているラットの海馬のCA1野において,他者の位置情報がどのように表象されるかを解析することにより,場所細胞の多くが自己の場所と他者の場所を同時に表象するジョイント場所細胞であることが明らかにされた.自己のゴールとは独立して他者の場所を表象する他者の場所細胞,および,自己あるいは他者にかかわらず反応する場所受容野をもつ共通場所細胞が同定され,さらに,他者の空間位置を場所細胞の発火のパターンから再構築できることが示された.この研究により,従来は自己の場所をコードすると考えられていた場所細胞が,他者の場所という異なる次元の空間事象をも表象することがはじめて示された.
動物の社会的な行動においては,その前提として,他者の場所や行動を認識する必要があると考えられる.今回,海馬が他者の位置情報を表象することが明らかにされたことから,海馬や脳のほかの領域が動物の社会性9,10) に関してどのように機能するのか,今後,さらに解明が進むことが期待される.
文 献
- O'Keefe, J. & Nadel, L.: The Hippocampus as a Cognitive Map. Oxford University Press, Oxford (1978)
- Buzsaki, G. & Moser, E. I.: Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci., 16, 130-138 (2013)[PubMed]
- Leutgeb, S., Leutgeb, J. K., Barnes, C. A. et al.: Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science, 309, 619-623 (2005)[PubMed]
- Fujisawa, S., Amarasingham, A., Harrison, M. T. et al.: Behavior-dependent short-term assembly dynamics in the medial prefrontal cortex. Nat. Neurosci., 11, 823-833 (2008)[PubMed]
- MacDonald, C. J., Lepage, K. Q., Eden, U. T. et al.: Hippocampal "time cells" bridge the gap in memory for discontiguous events. Neuron, 71, 737-749 (2011)[PubMed]
- Frank, L. M., Brown, E. N. & Wilson, M.: Trajectory encoding in the hippocampus and entorhinal cortex. Neuron, 27, 169-178 (2000)[PubMed]
- Rizzolatti, G. & Craighero, L.: The mirror-neuron system. Annu. Rev. Neurosci., 27, 169-192 (2004)[PubMed]
- Wilson, M. A. & McNaughton, B. L.: Dynamics of the hippocampal ensemble code for space. Science, 261, 1055-1058 (1993)[PubMed]
- Quiroga, R. Q., Reddy, L., Kreiman, G. et al.: Invariant visual representation by single neurons in the human brain. Nature, 435, 1102-1107 (2005)[PubMed]
- Omer, D. B., Maimon, S. R., Las, L. et al.: Social place-cells in the bat hippocampus. Science, 359, 218-224 (2018)[PubMed]
著者プロフィール
略歴:2011年 京都大学大学院医学研究科 修了,同年 大阪バイオサイエンス研究所 研究員を経て,2013年より理化学研究所脳科学総合研究センター 研究員.
藤澤 茂義(Shigeyoshi Fujisawa)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://fujisawalab.brain.riken.jp/aboutjp.html
© 2018 檀上輝子・藤澤茂義 Licensed under CC 表示 2.1 日本
