微小管ネットワークおよび細胞の極性に依存的な上皮組織の折りたたみ構造の形成機構
武田美智子・Mustafa M. Sami・Yu-Chiun Wang
(理化学研究所多細胞システム形成研究センター 上皮形態形成研究チーム)
email:武田美智子
DOI: 10.7875/first.author.2018.002
A homeostatic apical microtubule network shortens cells for epithelial folding via a basal polarity shift.
Michiko Takeda, Mustafa M. Sami, Yu-Chiun Wang
Nature Cell Biology, 20, 36-45 (2018)
上皮組織に高次構造が形成されるしくみを理解するためには,個々の細胞の形態形成の機構を明らかにすることが必須である.これまでに,ミオシンの収縮による細胞の頂端面の収縮が広く知られているが,この研究において,筆者らは,微小管がネットワークを形成し能動的に細胞の形態変化を制御するしくみを見い出した.Patroninにより頂端膜に裏打ちされた微小管ネットワークは,モータータンパク質Dyneinを駆動力とした頂端面を押し上げる力を生じ,ドーム様構造の形成に貢献する.また,微小管ネットワークはPatroninとKataninとの拮抗による急速なリモデリングのもとにあり,組織に形態変化が起こるまでは細胞の大きさおよび形態の恒常性を維持する.のちに折りたたみ構造の形成されるときには頂端面が胚の表面から沈み込むが,この変形には細胞の極性の変化に応答したPatroninの再分布が重要な役割をはたすことが示唆された.この研究により,微小管ネットワークが細胞の極性の変化に依存して細胞の形態変化を制御する機構が明らかにされた.
上皮組織の折りたたみ構造の形成は,2次元のシートが折りたたまれ3次元の構造へと変化する,発生の過程においてよくみられる基本的な形態形成の過程のひとつである.この形態の変化を起点として,内部組織の形成や機能的な器官の形成へと発生の過程はつづいていく.さまざまな3次元の形態形成の機構が研究されているが,単層の上皮組織が折りたたまれる過程は,陥入がはじまる領域において生じた局所的な力,あるいは,組織の近辺の不均一な力学的な環境から生じた広域的なストレスにより押し曲げられることによりはじまる.いずれの場合も,ミオシンに依存性の収縮の変化が深くかかわることが明らかにされてきた1).しかし,このようなミオシンの顕著な変化の起こらない上皮組織において,どのような機構により組織の変形が起こるのかは知られていない.
ショウジョウバエの原腸胚に形成されるdorsal foldは,細胞膜の形成をへて上皮細胞層が形成されたのち,背側中心線を横切って並ぶ2つの線に並んだ領域において細胞の背丈が減少することから形成がはじまる,上皮組織の折りたたみ構造である2)(図1).dorsal foldが形成される前後の背側の領域においては,ほかの領域とは異なり,ミオシンの活性は低く均一に保たれている2,3).そのため,dorsal foldの形成の過程はこれまでに知られていない折りたたみ構造の形成機構を解明するうえで重要なモデルである.以前に,筆者らは,細胞の背丈が減少するまえに,側底膜を特定するMARKファミリーキナーゼPar-1が減少しその局在の境界が基底側へと移動し,その結果,接着結合が基底側へと移動することを報告した2).陥入先行細胞においては,この極性の基底の方向への変化につづき頂端面は収縮し胚の表面から沈み込むが,これらのイベントを結ぶ機構は明らかにされていない.この研究においては,上皮組織の折りたたみ構造の形成において,極性の変化が細胞の形態変化をみちびく機構の解明をめざした.
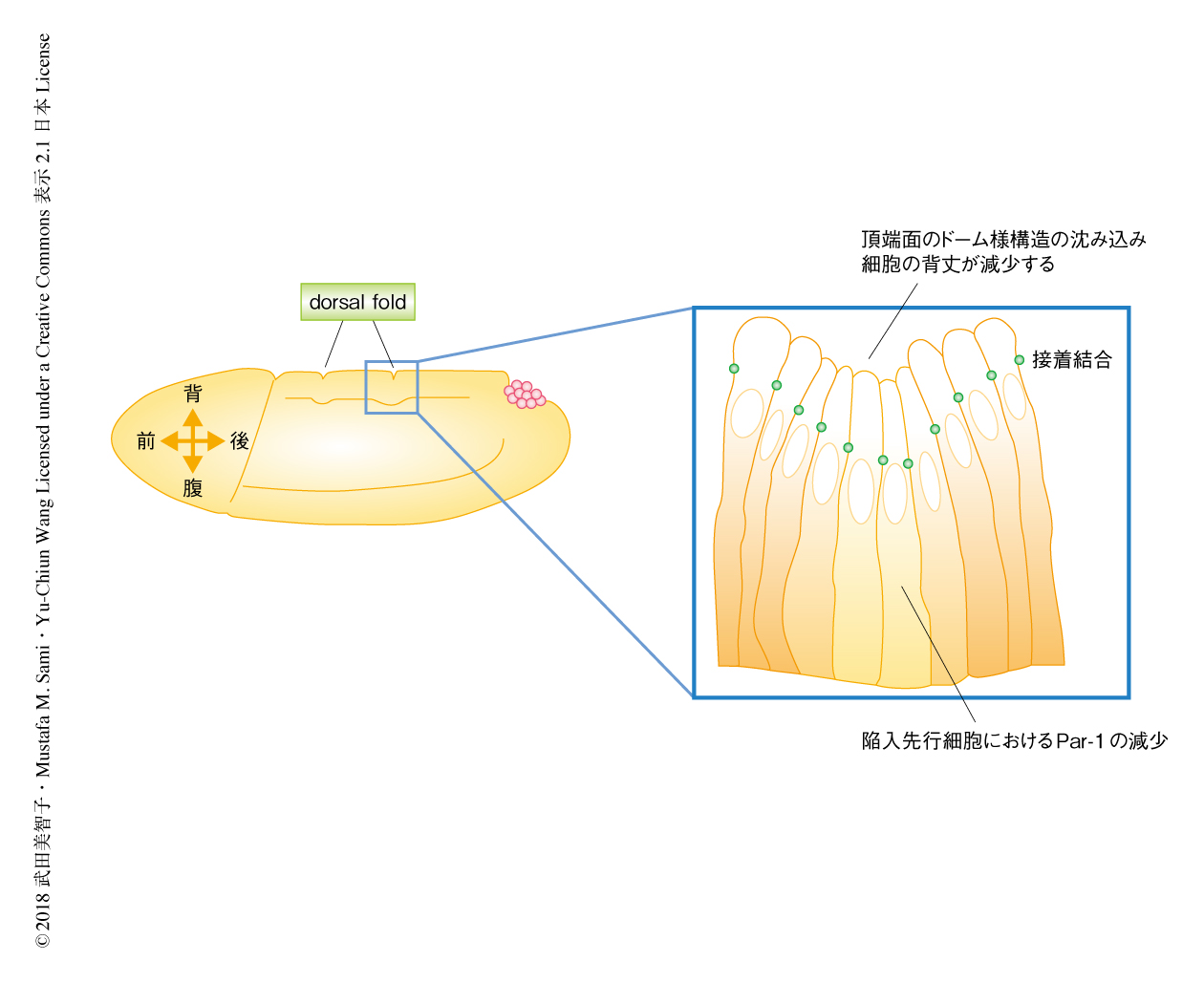
近年,ショウジョウバエの卵母細胞において,Par-1は微小管マイナス端結合タンパク質Patronin/CAMSAPの局在を制御することが報告された4).そこで,Par-1の減少につづき,Patroninに依存的な微小管の構造の制御がdorsal foldの形成の開始をみちびく可能性について検証した.
細胞の形態変化におけるPatroninの機能について調べるため,RNAi法によりPatroninをノックダウンし,微小管を蛍光により標識して細胞の頂端の側のライブイメージングを行った.このイメージングにより,単量体のチューブリンに由来すると考えられる中程度のシグナルから頂端面の形態を推定し,また,繊維状の強いシグナルより微小管フィラメントを観察することが可能である.野生型の胚においては,細胞膜が形成されるときには荒かった頂端面が,陥入が開始する10分ほどまえからなめらかに整い,細胞ごとに半球形のドーム様構造が形成された.また,このドーム様構造の底部にはリング状の微小管の構造がみられた.一方,Patroninをノックダウンした胚においては,繊維状に突出した頂端面の構造が現われ,ドーム様構造およびリング状構造ともに形成されなかった.さらに,頂端面の形態を,円形にどれほど近いかを示すcircularityおよびなめらかさを示すsolidityにより定量的に解析した結果,野生型の胚においては陥入の直前までにcircularityおよびsolidityは上昇したが,Patroninをノックダウンした胚では低いままであった.したがって,Patroninは頂端面におけるなめらかなドーム様構造の形成に必須であることが明らかにされた.
それでは,Patroninはどのように頂端面の形態形成に貢献するのだろうか? 頂端面における微小管の形状および構成について調べるため,不安定型の微小管および安定型の微小管を免疫染色し,超解像度でのイメージングを行った.野生型の胚のドーム様構造の内面においてはおもに不安定型の微小管から構成された非中心体微小管がフィラメント状のネットワークを形成していた.一方,中心体や核膜をかこんで基底側へと伸びる中心体微小管は不安定型と安定型とが混合して存在し,頂端面の微小管ネットワークとは性質が異なっていた.Patroninをノックダウンした胚の頂端面においては不安定型の微小管から構成されたフィラメントが増加し束状の構造になっていたことから,Patroninは頂端面において微小管ネットワークの構造の形成を制御し,ドーム様構造を形成することが示唆された.また,頂端面には微小管マイナス端結合タンパク質であるPatroninにくわえ,微小管のプラス端に集積するEB1の局在も確認され,無秩序な微小管ネットワークが形成されていることが示唆された.
微小管ネットワークに裏打ちされた頂端面にドーム様構造を形成する機構は何だろうか? 微小管フィラメントは湾曲に対する剛性をもつことが知られている.そこで,Patroninにより微小管のマイナス端を固定された微小管ネットワークを,微小管の逆行性モータータンパク質であるDyneinが架橋しつつ移動することにより,頂端面を外向きにささえる力が生じる,との仮説をたてた.Dyneinのノックダウンによる機能欠損解析において,Patroninによる微小管ネットワークの形成に影響はなかったが,頂端面の形態は繊維状に突出しドーム様構造は形成されなかった.頂端面の形態を定量的に示すcircularityおよびsolidityは低いままで,Patroninのノックダウンと同様であった.よって,微小管ネットワークにおいて,Dyneinに依存的に生じる外方向へと押し上げる力を駆動力として,なめらかなドーム様構造をささえる力学的なモデルが示唆された.
dorsal foldの形成が開始するときにPar-1の減少につづき頂端面が沈み込む機構について明らかにするため,細胞におけるPatroninの局在を経時的に観察した.細胞膜の形成の初期にはPatroninは中心体から頂端の側の細胞質に散在していたが,頂端面のドーム様構造の形成の直前に頂端面に局在を変え点状のパターンになった.頂端面に局在するaPKC,Par-6,Bazookaの阻害実験から,極性の特定化に依存してPatroninは頂端面に局在することが示された.以上より,極性化,Patroninの頂端面への移行,頂端面のドーム様構造の形成の機能的なつながりが示唆された.
dorsal foldの形成が開始したときの陥入先行細胞において,Par-1の減少につづく接着結合の基底の方向への移動にともない,Patroninの局在の境界も基底の方向へと移動し,拡張した頂端の側の領域に再分布した.また,Par-1の阻害実験において,Patroninの局在は基底の方向へ拡張したことから,Par-1はPatroninの局在を頂端の側に制限することが明らかにされた.さらに,野生型におけるPatroninの局在の定量的な解析から,陥入先行細胞のPatroninは,近隣の細胞と比較して,より基底の側へと局在が拡張していることが示された.これらの結果から,頂端面に局在化したPatroninはPar-1の減少に応答し基底の側へと再分布することが明らかにされた.
そこで,陥入細胞においてPatroninの再分布によりドーム様構造を押し上げる力が減少することが原因となり細胞の頂端面が沈み込む可能性について検証した.逆に,Patroninを均一に過剰発現すると,通常の陥入細胞の領域において接着結合の位置は基底の方向へと移動するにもかかわらず,折りたたみ構造は形成されなかった.頂端面の微小管ネットワークにおいては安定型および不安定型の微小管が増加し,太いアーチ状の構造が形成された.また,頂端面は波打つように動き,頂端面の沈下が阻害されたことから,周辺の組織からうける力学的なストレスに抵抗していると予想された.このように,Patroninが過剰になると微小管の安定性が増大し微小管ネットワークはまとまった束状になり,力学的な抵抗力が増強されることから,Patroninの再分布による陥入細胞と近隣の細胞とのあいだのPatroninの分布の不均衡が折りたたみ構造の形成に必要であると考えられた.
頂端面における微小管ネットワークはおもに不安定型の微小管から構成され,極性の変化に応答し急速に再分布することから,つねに急速なリモデリングのもとにある可能性が考えられた.微小管切断酵素KataninとPatroninとの結合が報告されていたことから4,5),微小管ネットワークにおけるKataninの機能および局在について検証した.Kataninは頂端面に点状の局在を示し,一部はPatroninと共局在し,Patroninをノックダウンすると頂端面におけるKataninの局在量は減少した.微小管ネットワークのリモデリングにおける役割を微小管フィラメントのカイモグラフ解析により検証すると,PatroninあるいはKataninを阻害すると微小管フィラメントの長時間の安定化がみられた.したがって,Patroninにより頂端面の微小管ネットワークに局在化されたKataninは,Patroninと複合体を形成し,Patroninと拮抗して微小管ネットワークのリモデリングを促進することが示唆された.
Kataninの細胞形態に関する機能について解析した.Kataninを減少させた胚においては,dorsal foldの形成のときに細胞の背丈の減少および頂端面の沈み込みが進行せず,頂端面は胚の表面に残存した.頂端面の形態変化を測定した結果,Kataninを減少させた胚の陥入先行細胞における頂端の側の領域は伸長しており,そのため,細胞の背丈の減少が阻害されると考えられた.したがって,Kataninを介した急速なリモデリングを維持した微小管ネットワークは頂端面の沈み込みの促進に必要であることが示された.これにくわえ,本来,上皮組織の細胞の形態は頂端面および側面ともに均一であるが,Patroninをノックダウンした胚あるいはKataninを減少させた胚においては不均一となった.よって,PatroninおよびKataninにより制御される正常な微小管ネットワークのリモデリングが細胞の大きさの維持に必要であることが示された.以上より,頂端面における微小管ネットワークのリモデリングは2段階の機能を担い,陥入よりまえには細胞形態の恒常性の維持,それにつづく陥入のときには極性の変化に応答して頂端面の形態の変化をみちびくと考えられた.
この研究において,ミオシンのレベルが低く均一に保たれた上皮組織における折りたたみ構造の形成機構のひとつとして,細胞の頂端面に裏打ちされた微小管ネットワークが細胞の形態形成を制御する分子的および力学的な機構が明らかにされた(図2).この微小管ネットワークはおもに不安定型の微小管から構成され,微小管フィラメントのプラス端とマイナス端を含んだ無秩序な構造を形成し,PatroninとKataninとのあいだのフィードバックによる急速なリモデリングのもとにあった.陥入のまえ,Patroninにより頂端面に固定された微小管ネットワークはDyneinの駆動力により頂端面をささえる外向きの力を生じ,頂端面のドーム様構造の形成に貢献する.このDyneinに依存的な微小管による力学的なモデルは,開いた傘を骨組みが押しささえている状態にたとえられる.また,微小管ネットワークのリモデリングは組織に存在する力学的なノイズを緩和し,細胞の形態および大きさの恒常性を維持する.陥入期にはいるとこの微小管ネットワークは異なる用途に利用され,陥入先行細胞においては極性の変化に応答したPatroninの再分布を介して頂端面を押しあげささえる力が減少し頂端面の沈み込みをみちびく.
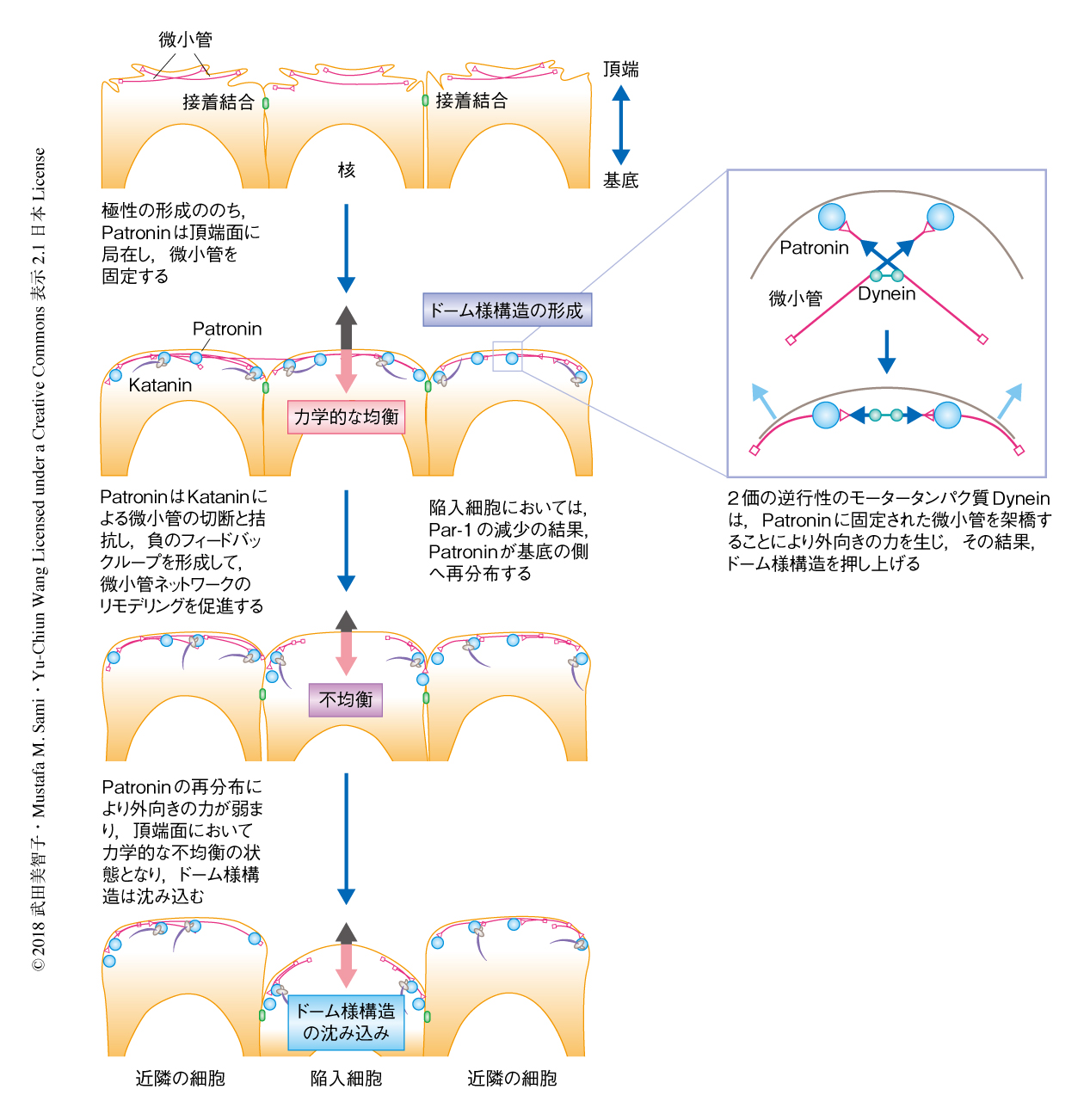
この力学的なモデルにおいては,陥入細胞の背丈の減少は微小管ネットワークの頂端面を押しあげる力が弱まることに起因し,力の方向はつねに外向きで,ミオシンによる直接的な頂端面の収縮とは逆である.筆者らは,陥入よりまえには微小管ネットワークが生じる力と均衡し,微小管ネットワークの力が減少する陥入のときに頂端面を下向きに押し込む,内向きの力がつねに存在する可能性を考えている(図2).このような微小管ネットワークに依存的に局所的に発生する力の不均衡が,組織において弱い領域,つまり,折りたたみ構造の開始点をつくりだすと思われる.
また,この研究により,上皮組織におけるPatronin/CAMSAPの細胞形態の制御に関する機能がはじめて報告された.頂端面の微小管ネットワークについては哺乳類の上皮細胞においても類似した構造が報告されており6),上皮細胞の共通な特徴である可能性がある.
この研究は,多種多様な発生の過程において普遍的にみられる細胞の陥入の過程をへたダイナミックな上皮組織の形態変化は,ミオシンの収縮のみでなく,能動的な微小管ネットワークにより制御される可能性もあることを示した.今後,上皮組織の形態形成を制御する多様な機構のさらなる解明につながることを期待する.
略歴:2012年 ドイツGoettingen大学大学院博士課程 修了,理化学研究所発生・再生科学総合研究センター(現 多細胞システム研究センター)テクニカルスタッフ.
研究テーマ:上皮組織の形態形成.
Mustafa M. Sami
理化学研究所多細胞システム研究センター テクニカルスタッフ.
Yu-Chiun Wang
理化学研究所多細胞システム研究センター チームリーダー.
研究室URL:http://www.cdb.riken.jp/epm/index-jp.html
© 2018 武田美智子・Mustafa M. Sami・Yu-Chiun Wang Licensed under CC 表示 2.1 日本
(理化学研究所多細胞システム形成研究センター 上皮形態形成研究チーム)
email:武田美智子
DOI: 10.7875/first.author.2018.002
A homeostatic apical microtubule network shortens cells for epithelial folding via a basal polarity shift.
Michiko Takeda, Mustafa M. Sami, Yu-Chiun Wang
Nature Cell Biology, 20, 36-45 (2018)
要 約
上皮組織に高次構造が形成されるしくみを理解するためには,個々の細胞の形態形成の機構を明らかにすることが必須である.これまでに,ミオシンの収縮による細胞の頂端面の収縮が広く知られているが,この研究において,筆者らは,微小管がネットワークを形成し能動的に細胞の形態変化を制御するしくみを見い出した.Patroninにより頂端膜に裏打ちされた微小管ネットワークは,モータータンパク質Dyneinを駆動力とした頂端面を押し上げる力を生じ,ドーム様構造の形成に貢献する.また,微小管ネットワークはPatroninとKataninとの拮抗による急速なリモデリングのもとにあり,組織に形態変化が起こるまでは細胞の大きさおよび形態の恒常性を維持する.のちに折りたたみ構造の形成されるときには頂端面が胚の表面から沈み込むが,この変形には細胞の極性の変化に応答したPatroninの再分布が重要な役割をはたすことが示唆された.この研究により,微小管ネットワークが細胞の極性の変化に依存して細胞の形態変化を制御する機構が明らかにされた.
はじめに
上皮組織の折りたたみ構造の形成は,2次元のシートが折りたたまれ3次元の構造へと変化する,発生の過程においてよくみられる基本的な形態形成の過程のひとつである.この形態の変化を起点として,内部組織の形成や機能的な器官の形成へと発生の過程はつづいていく.さまざまな3次元の形態形成の機構が研究されているが,単層の上皮組織が折りたたまれる過程は,陥入がはじまる領域において生じた局所的な力,あるいは,組織の近辺の不均一な力学的な環境から生じた広域的なストレスにより押し曲げられることによりはじまる.いずれの場合も,ミオシンに依存性の収縮の変化が深くかかわることが明らかにされてきた1).しかし,このようなミオシンの顕著な変化の起こらない上皮組織において,どのような機構により組織の変形が起こるのかは知られていない.
ショウジョウバエの原腸胚に形成されるdorsal foldは,細胞膜の形成をへて上皮細胞層が形成されたのち,背側中心線を横切って並ぶ2つの線に並んだ領域において細胞の背丈が減少することから形成がはじまる,上皮組織の折りたたみ構造である2)(図1).dorsal foldが形成される前後の背側の領域においては,ほかの領域とは異なり,ミオシンの活性は低く均一に保たれている2,3).そのため,dorsal foldの形成の過程はこれまでに知られていない折りたたみ構造の形成機構を解明するうえで重要なモデルである.以前に,筆者らは,細胞の背丈が減少するまえに,側底膜を特定するMARKファミリーキナーゼPar-1が減少しその局在の境界が基底側へと移動し,その結果,接着結合が基底側へと移動することを報告した2).陥入先行細胞においては,この極性の基底の方向への変化につづき頂端面は収縮し胚の表面から沈み込むが,これらのイベントを結ぶ機構は明らかにされていない.この研究においては,上皮組織の折りたたみ構造の形成において,極性の変化が細胞の形態変化をみちびく機構の解明をめざした.
1.Patroninは頂端膜を裏打ちする微小管ネットワークを組織し頂端面にドーム様構造を形成する
近年,ショウジョウバエの卵母細胞において,Par-1は微小管マイナス端結合タンパク質Patronin/CAMSAPの局在を制御することが報告された4).そこで,Par-1の減少につづき,Patroninに依存的な微小管の構造の制御がdorsal foldの形成の開始をみちびく可能性について検証した.
細胞の形態変化におけるPatroninの機能について調べるため,RNAi法によりPatroninをノックダウンし,微小管を蛍光により標識して細胞の頂端の側のライブイメージングを行った.このイメージングにより,単量体のチューブリンに由来すると考えられる中程度のシグナルから頂端面の形態を推定し,また,繊維状の強いシグナルより微小管フィラメントを観察することが可能である.野生型の胚においては,細胞膜が形成されるときには荒かった頂端面が,陥入が開始する10分ほどまえからなめらかに整い,細胞ごとに半球形のドーム様構造が形成された.また,このドーム様構造の底部にはリング状の微小管の構造がみられた.一方,Patroninをノックダウンした胚においては,繊維状に突出した頂端面の構造が現われ,ドーム様構造およびリング状構造ともに形成されなかった.さらに,頂端面の形態を,円形にどれほど近いかを示すcircularityおよびなめらかさを示すsolidityにより定量的に解析した結果,野生型の胚においては陥入の直前までにcircularityおよびsolidityは上昇したが,Patroninをノックダウンした胚では低いままであった.したがって,Patroninは頂端面におけるなめらかなドーム様構造の形成に必須であることが明らかにされた.
それでは,Patroninはどのように頂端面の形態形成に貢献するのだろうか? 頂端面における微小管の形状および構成について調べるため,不安定型の微小管および安定型の微小管を免疫染色し,超解像度でのイメージングを行った.野生型の胚のドーム様構造の内面においてはおもに不安定型の微小管から構成された非中心体微小管がフィラメント状のネットワークを形成していた.一方,中心体や核膜をかこんで基底側へと伸びる中心体微小管は不安定型と安定型とが混合して存在し,頂端面の微小管ネットワークとは性質が異なっていた.Patroninをノックダウンした胚の頂端面においては不安定型の微小管から構成されたフィラメントが増加し束状の構造になっていたことから,Patroninは頂端面において微小管ネットワークの構造の形成を制御し,ドーム様構造を形成することが示唆された.また,頂端面には微小管マイナス端結合タンパク質であるPatroninにくわえ,微小管のプラス端に集積するEB1の局在も確認され,無秩序な微小管ネットワークが形成されていることが示唆された.
2.モータータンパク質Dyneinは微小管ネットワークを押し上げる力を生じ頂端面のドーム様構造をささえる
微小管ネットワークに裏打ちされた頂端面にドーム様構造を形成する機構は何だろうか? 微小管フィラメントは湾曲に対する剛性をもつことが知られている.そこで,Patroninにより微小管のマイナス端を固定された微小管ネットワークを,微小管の逆行性モータータンパク質であるDyneinが架橋しつつ移動することにより,頂端面を外向きにささえる力が生じる,との仮説をたてた.Dyneinのノックダウンによる機能欠損解析において,Patroninによる微小管ネットワークの形成に影響はなかったが,頂端面の形態は繊維状に突出しドーム様構造は形成されなかった.頂端面の形態を定量的に示すcircularityおよびsolidityは低いままで,Patroninのノックダウンと同様であった.よって,微小管ネットワークにおいて,Dyneinに依存的に生じる外方向へと押し上げる力を駆動力として,なめらかなドーム様構造をささえる力学的なモデルが示唆された.
3.Par-1の減少に応答した頂端面のPatroninの基底の方向への再分布により頂端面は沈み込む
dorsal foldの形成が開始するときにPar-1の減少につづき頂端面が沈み込む機構について明らかにするため,細胞におけるPatroninの局在を経時的に観察した.細胞膜の形成の初期にはPatroninは中心体から頂端の側の細胞質に散在していたが,頂端面のドーム様構造の形成の直前に頂端面に局在を変え点状のパターンになった.頂端面に局在するaPKC,Par-6,Bazookaの阻害実験から,極性の特定化に依存してPatroninは頂端面に局在することが示された.以上より,極性化,Patroninの頂端面への移行,頂端面のドーム様構造の形成の機能的なつながりが示唆された.
dorsal foldの形成が開始したときの陥入先行細胞において,Par-1の減少につづく接着結合の基底の方向への移動にともない,Patroninの局在の境界も基底の方向へと移動し,拡張した頂端の側の領域に再分布した.また,Par-1の阻害実験において,Patroninの局在は基底の方向へ拡張したことから,Par-1はPatroninの局在を頂端の側に制限することが明らかにされた.さらに,野生型におけるPatroninの局在の定量的な解析から,陥入先行細胞のPatroninは,近隣の細胞と比較して,より基底の側へと局在が拡張していることが示された.これらの結果から,頂端面に局在化したPatroninはPar-1の減少に応答し基底の側へと再分布することが明らかにされた.
そこで,陥入細胞においてPatroninの再分布によりドーム様構造を押し上げる力が減少することが原因となり細胞の頂端面が沈み込む可能性について検証した.逆に,Patroninを均一に過剰発現すると,通常の陥入細胞の領域において接着結合の位置は基底の方向へと移動するにもかかわらず,折りたたみ構造は形成されなかった.頂端面の微小管ネットワークにおいては安定型および不安定型の微小管が増加し,太いアーチ状の構造が形成された.また,頂端面は波打つように動き,頂端面の沈下が阻害されたことから,周辺の組織からうける力学的なストレスに抵抗していると予想された.このように,Patroninが過剰になると微小管の安定性が増大し微小管ネットワークはまとまった束状になり,力学的な抵抗力が増強されることから,Patroninの再分布による陥入細胞と近隣の細胞とのあいだのPatroninの分布の不均衡が折りたたみ構造の形成に必要であると考えられた.
4.Patroninは微小管切断酵素Kataninを微小管のマイナス端に局在させ微小管ネットワークのリモデリングを促進する
頂端面における微小管ネットワークはおもに不安定型の微小管から構成され,極性の変化に応答し急速に再分布することから,つねに急速なリモデリングのもとにある可能性が考えられた.微小管切断酵素KataninとPatroninとの結合が報告されていたことから4,5),微小管ネットワークにおけるKataninの機能および局在について検証した.Kataninは頂端面に点状の局在を示し,一部はPatroninと共局在し,Patroninをノックダウンすると頂端面におけるKataninの局在量は減少した.微小管ネットワークのリモデリングにおける役割を微小管フィラメントのカイモグラフ解析により検証すると,PatroninあるいはKataninを阻害すると微小管フィラメントの長時間の安定化がみられた.したがって,Patroninにより頂端面の微小管ネットワークに局在化されたKataninは,Patroninと複合体を形成し,Patroninと拮抗して微小管ネットワークのリモデリングを促進することが示唆された.
Kataninの細胞形態に関する機能について解析した.Kataninを減少させた胚においては,dorsal foldの形成のときに細胞の背丈の減少および頂端面の沈み込みが進行せず,頂端面は胚の表面に残存した.頂端面の形態変化を測定した結果,Kataninを減少させた胚の陥入先行細胞における頂端の側の領域は伸長しており,そのため,細胞の背丈の減少が阻害されると考えられた.したがって,Kataninを介した急速なリモデリングを維持した微小管ネットワークは頂端面の沈み込みの促進に必要であることが示された.これにくわえ,本来,上皮組織の細胞の形態は頂端面および側面ともに均一であるが,Patroninをノックダウンした胚あるいはKataninを減少させた胚においては不均一となった.よって,PatroninおよびKataninにより制御される正常な微小管ネットワークのリモデリングが細胞の大きさの維持に必要であることが示された.以上より,頂端面における微小管ネットワークのリモデリングは2段階の機能を担い,陥入よりまえには細胞形態の恒常性の維持,それにつづく陥入のときには極性の変化に応答して頂端面の形態の変化をみちびくと考えられた.
おわりに
この研究において,ミオシンのレベルが低く均一に保たれた上皮組織における折りたたみ構造の形成機構のひとつとして,細胞の頂端面に裏打ちされた微小管ネットワークが細胞の形態形成を制御する分子的および力学的な機構が明らかにされた(図2).この微小管ネットワークはおもに不安定型の微小管から構成され,微小管フィラメントのプラス端とマイナス端を含んだ無秩序な構造を形成し,PatroninとKataninとのあいだのフィードバックによる急速なリモデリングのもとにあった.陥入のまえ,Patroninにより頂端面に固定された微小管ネットワークはDyneinの駆動力により頂端面をささえる外向きの力を生じ,頂端面のドーム様構造の形成に貢献する.このDyneinに依存的な微小管による力学的なモデルは,開いた傘を骨組みが押しささえている状態にたとえられる.また,微小管ネットワークのリモデリングは組織に存在する力学的なノイズを緩和し,細胞の形態および大きさの恒常性を維持する.陥入期にはいるとこの微小管ネットワークは異なる用途に利用され,陥入先行細胞においては極性の変化に応答したPatroninの再分布を介して頂端面を押しあげささえる力が減少し頂端面の沈み込みをみちびく.
この力学的なモデルにおいては,陥入細胞の背丈の減少は微小管ネットワークの頂端面を押しあげる力が弱まることに起因し,力の方向はつねに外向きで,ミオシンによる直接的な頂端面の収縮とは逆である.筆者らは,陥入よりまえには微小管ネットワークが生じる力と均衡し,微小管ネットワークの力が減少する陥入のときに頂端面を下向きに押し込む,内向きの力がつねに存在する可能性を考えている(図2).このような微小管ネットワークに依存的に局所的に発生する力の不均衡が,組織において弱い領域,つまり,折りたたみ構造の開始点をつくりだすと思われる.
また,この研究により,上皮組織におけるPatronin/CAMSAPの細胞形態の制御に関する機能がはじめて報告された.頂端面の微小管ネットワークについては哺乳類の上皮細胞においても類似した構造が報告されており6),上皮細胞の共通な特徴である可能性がある.
この研究は,多種多様な発生の過程において普遍的にみられる細胞の陥入の過程をへたダイナミックな上皮組織の形態変化は,ミオシンの収縮のみでなく,能動的な微小管ネットワークにより制御される可能性もあることを示した.今後,上皮組織の形態形成を制御する多様な機構のさらなる解明につながることを期待する.
文 献
- Murrell, M., Oakes, P. W., Lenz, M. et al.: Forcing cells into shape: the mechanics of actomyosin contractility. Nat. Rev. Mol. Cell Biol., 16, 486-498 (2015)[PubMed]
- Wang, Y. -C., Khan, Z., Kaschube, M. et al.: Differential positioning of adherens junctions is associated with initiation of epithelial folding. Nature, 484, 390-393 (2012)[PubMed]
- Rauzi, M., Krzic, U., Saunders, T. E. et al.: Embryo-scale tissue mechanics during Drosophila gastrulation movements. Nat. Commun., 6, 8677 (2015)[PubMed]
- Nashchekin, D., Fernandes, A. R. & Johnston, D. S.: Patronin/Shot cortical foci assemble the noncentrosomal microtubule array that specifies the Drosophila anterior-posterior axis. Dev. Cell, 38, 61-72 (2016)[PubMed]
- Jiang, K., Hua, S., Mohan, R. et al.: Microtubule minus-end stabilization by polymerization-driven CAMSAP deposition. Dev. Cell, 28, 295-309 (2014)[PubMed]
- Yano, T., Matsui, T., Tamura, A. et al.: The association of microtubules with tight junctions is promoted by cingulin phosphorylation by AMPK. J. Cell Biol., 203, 605-614 (2013)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2012年 ドイツGoettingen大学大学院博士課程 修了,理化学研究所発生・再生科学総合研究センター(現 多細胞システム研究センター)テクニカルスタッフ.
研究テーマ:上皮組織の形態形成.
Mustafa M. Sami
理化学研究所多細胞システム研究センター テクニカルスタッフ.
Yu-Chiun Wang
理化学研究所多細胞システム研究センター チームリーダー.
研究室URL:http://www.cdb.riken.jp/epm/index-jp.html
© 2018 武田美智子・Mustafa M. Sami・Yu-Chiun Wang Licensed under CC 表示 2.1 日本
