大脳皮質においてニューロンにより構築される格子構造
丸岡久人・細谷俊彦
(理化学研究所脳科学総合研究センター 局所神経回路研究チーム)
email:細谷俊彦
DOI: 10.7875/first.author.2017.131
Lattice system of functionally distinct cell types in the neocortex.
Hisato Maruoka, Nao Nakagawa, Shun Tsuruno, Seiichiro Sakai, Taisuke Yoneda, Toshihiko Hosoya
Science, 358, 610-615 (2017)
大脳皮質に単一の神経回路がくり返した構造が存在するのかどうかは不明であった.この研究において,筆者らは,マウスにおいて大脳皮質の6層ある細胞層のひとつである第5層を解析し,大部分のニューロンがニューロンのタイプに特異的なカラム状の小さなクラスターを形成することを発見した.マイクロカラムとよばれるこの構造は,六方格子状の規則的な配置をとっており,大脳皮質のさまざまな領野に共通に存在した.同一のマイクロカラムに含まれるニューロンは同期した神経活動を示し,方位選択性や眼優位性など刺激に対する応答性が類似していた.さらに,神経回路が形成される時期にはマイクロカラムのニューロンはギャップ結合により結合しており,のちに共通な入力をうけることがわかった.以上の結果から,第5層はマイクロカラムがくり返した神経回路に組織化されており,個々のマイクロカラムは機能単位として動作することが示された.したがって,多数のマイクロカラムによる並列処理が大脳皮質の多様な機能において共通な情報処理を担うと考えられた.
ヒトの大脳は100億個以上のニューロンからなるきわめて複雑な組織である.大脳はさまざまな領野に分かれており,それぞれ,感覚処理,運動制御,言語,思考など異なる機能をつかさどる.大脳皮質における神経回路の構造については100年以上にわたり研究がなされているが,その複雑さからまだ不明な点が多く,脳の機能の理解において大きなさまたげになっている.なかでも,大脳皮質に単一の神経回路がくり返した構造が存在するのかどうかは不明であった.1960年代ごろから,ネコやサルなどにおいて,方位選択性カラムなどのいわゆる皮質カラムが発見されたが,これらは大脳皮質の視覚野のみにかぎられ,さらに,ラットやマウスなどではみられないことから,大脳皮質の普遍的な機能単位ではないと考えられている1,2).
大脳のニューロンは厚さ1~2 mmの大脳皮質を形成しており,さらに,大脳皮質は機能の異なる6つの層に分かれている.大脳皮質の深部に位置する第5層はニューロンの分類が比較的進んでおり,皮質下投射細胞および皮質投射細胞の2種類の興奮性ニューロンと,パルブアルブミン陽性細胞およびソマトスタチン陽性細胞の2種類の抑制性ニューロンの計4種類のニューロンからおもに構成されることがわかっている.
2011年,筆者らの研究室は,マウスの脳において皮質下投射細胞が幅1~2細胞,高さ数細胞の細長いクラスターを形成すること,および,多数の皮質下投射細胞マイクロカラムとよばれるこの構造が第5層において並んでいることを発見した3).これとは独立に,2012年,別の研究グループも,ヒトの脳の言語野において同様な構造を発見している4).一方,これまで,皮質下投射細胞マイクロカラムの機能やほかのニューロンの空間的な配置については不明であった.
これまでの研究においては脳をスライスして解析していたが,今回は,新しく開発された,脳を丸ごと3次元解析する技術を導入した5-7).神経終末への色素の注入や抗体染色などを用いて個々のニューロンのタイプを可視化し,脳の全体を透明化して3次元撮影することにより,第5層において1個体あたり数千~数万個のニューロンの位置座標を決定した.その結果,マウスの大脳皮質の第5層においては,1)皮質下投射細胞マイクロカラムは視覚野,体性感覚野,運動野といった機能の異なる複数の領野において共通にみられ,ハニカム状の六方格子配列をとり並んでいること,2)皮質投射細胞もマイクロカラムを形成し,この皮質投射細胞マイクロカラムは皮質下投射細胞マイクロカラムとは互い違いに並んでいること,3)パルブアルブミン陽性細胞およびソマトスタチン陽性細胞は皮質下投射細胞マイクロカラムに選択的に含まれ,皮質投射細胞マイクロカラムには含まれないこと,が明らかにされた(図1).
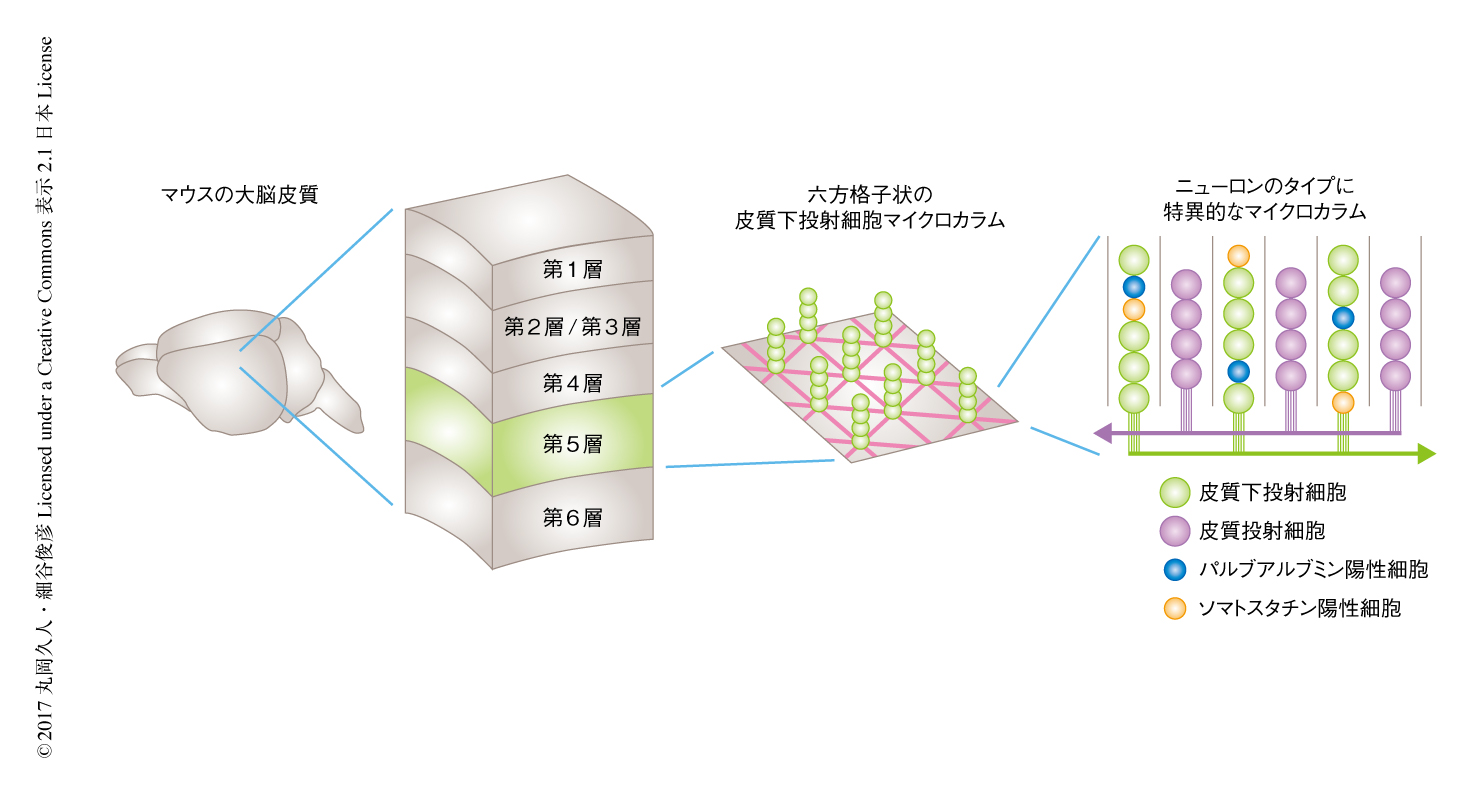
マイクロカラムの機能を調べるため,皮質下投射細胞の神経活動を解析した.ニューロンが発火するとCa2+の濃度が上昇する.そこで,皮質下投射細胞にCa2+の濃度の上昇により蛍光強度が上昇するCa2+センサー8) を導入し,生体脳イメージングを実施した.その結果,同一のマイクロカラムに含まれるニューロンは同期した神経活動を示すことが明らかにされた.この同期した神経活動は視覚野,体性感覚野,運動野の皮質下投射細胞マイクロカラムにおいて共通にみられた.さらに,視覚野においては,同一のマイクロカラムに含まれるニューロンは方位選択性および眼優位性が類似していた.以上より,個々のマイクロカラムはそれぞれ特定の情報を処理しており,大脳皮質の幅広い領野の共通な機能単位として動作することが明らかにされた.
電気生理学的な解析により,同じマイクロカラムに含まれるニューロンは同じニューロンからの入力をうけていることが明らかにされ,この入力が同期した神経活動をもたらす可能性が示された.また,大脳皮質において神経回路が形成される時期には,同じマイクロカラムに含まれるニューロンはギャップ結合により電気的に結合していることがわかった.このギャップ結合は,マイクロカラムに特異的な神経回路の形成を誘導する可能性があると考えられた.
以上の結果から,大脳皮質の幅広い領域において第5層はマイクロカラムがくり返した神経回路に組織化されていることが示された(図1).個々のマイクロカラムは要素的な情報処理を担う機能モジュールであると考えられ,多数のマイクロカラムによる並列処理が第5層における情報処理を担うことが示された.この神経回路の構造は大脳皮質のさまざまな領野に存在したことから,感覚処理,運動制御,言語処理などの大脳の多様な機能に共通する情報処理を担うと考えられた.
第5層がマイクロカラムのくり返しから構成されていたことから,10個程度のニューロンからなる単一のマイクロカラムの機能が明らかにされれば,第5層の機能について深い理解が得られる可能性がある.さらに,マイクロカラムは大脳皮質のさまざまな領野において共通な機能単位であるため,感覚,運動制御,言語など脳の広範な機能に共通する普遍的な情報処理の機構が明らかにされることが期待される.
今回の研究においては,ニューロンの分類が比較的進んでいる第5層を対象とした.近年,大脳皮質のほかの層においても遺伝子の発現などを指標としたニューロンの分類が進められているため,この情報を用いてマイクロカラム様の構造が探索されることが期待される.実際,ギャップ結合により結合したカラム状のクラスターは第5層のほかにも存在するため9,10),大脳皮質のほかの層にもマイクロカラムが存在する可能性がある.
個々のマイクロカラムに含まれる細胞は特定の方位選択性や眼優位性をもつ.一方,方位選択性カラムや眼優位性カラムなど古典的な皮質カラムは細胞数十個分ほどの幅があり,マイクロカラムより幅広い構造をもつ.したがって,ネコやサルなどにおいては方位選択性や眼優位性の類似した多数のマイクロカラムが並ぶことにより皮質カラムが形成されている可能性がある.もし,このことが示されれば,皮質カラムとそれ以外のさまざまな機能をもつ大脳皮質の領野の統一的な理解が可能になると期待される.
略歴:2006年 大阪大学大学院医学系研究科博士課程 修了,同年より理化学研究所脳科学総合研究センター 博士研究員.
研究テーマ:大脳皮質から出力する神経回路における情報処理の機構.
関心事:マイクロカラムが生体においてどのように機能するのか.
細谷 俊彦(Toshihiko Hosoya)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://t-hosoya.brain.riken.jp/
© 2017 丸岡久人・細谷俊彦 Licensed under CC 表示 2.1 日本
(理化学研究所脳科学総合研究センター 局所神経回路研究チーム)
email:細谷俊彦
DOI: 10.7875/first.author.2017.131
Lattice system of functionally distinct cell types in the neocortex.
Hisato Maruoka, Nao Nakagawa, Shun Tsuruno, Seiichiro Sakai, Taisuke Yoneda, Toshihiko Hosoya
Science, 358, 610-615 (2017)
要 約
大脳皮質に単一の神経回路がくり返した構造が存在するのかどうかは不明であった.この研究において,筆者らは,マウスにおいて大脳皮質の6層ある細胞層のひとつである第5層を解析し,大部分のニューロンがニューロンのタイプに特異的なカラム状の小さなクラスターを形成することを発見した.マイクロカラムとよばれるこの構造は,六方格子状の規則的な配置をとっており,大脳皮質のさまざまな領野に共通に存在した.同一のマイクロカラムに含まれるニューロンは同期した神経活動を示し,方位選択性や眼優位性など刺激に対する応答性が類似していた.さらに,神経回路が形成される時期にはマイクロカラムのニューロンはギャップ結合により結合しており,のちに共通な入力をうけることがわかった.以上の結果から,第5層はマイクロカラムがくり返した神経回路に組織化されており,個々のマイクロカラムは機能単位として動作することが示された.したがって,多数のマイクロカラムによる並列処理が大脳皮質の多様な機能において共通な情報処理を担うと考えられた.
はじめに
ヒトの大脳は100億個以上のニューロンからなるきわめて複雑な組織である.大脳はさまざまな領野に分かれており,それぞれ,感覚処理,運動制御,言語,思考など異なる機能をつかさどる.大脳皮質における神経回路の構造については100年以上にわたり研究がなされているが,その複雑さからまだ不明な点が多く,脳の機能の理解において大きなさまたげになっている.なかでも,大脳皮質に単一の神経回路がくり返した構造が存在するのかどうかは不明であった.1960年代ごろから,ネコやサルなどにおいて,方位選択性カラムなどのいわゆる皮質カラムが発見されたが,これらは大脳皮質の視覚野のみにかぎられ,さらに,ラットやマウスなどではみられないことから,大脳皮質の普遍的な機能単位ではないと考えられている1,2).
大脳のニューロンは厚さ1~2 mmの大脳皮質を形成しており,さらに,大脳皮質は機能の異なる6つの層に分かれている.大脳皮質の深部に位置する第5層はニューロンの分類が比較的進んでおり,皮質下投射細胞および皮質投射細胞の2種類の興奮性ニューロンと,パルブアルブミン陽性細胞およびソマトスタチン陽性細胞の2種類の抑制性ニューロンの計4種類のニューロンからおもに構成されることがわかっている.
2011年,筆者らの研究室は,マウスの脳において皮質下投射細胞が幅1~2細胞,高さ数細胞の細長いクラスターを形成すること,および,多数の皮質下投射細胞マイクロカラムとよばれるこの構造が第5層において並んでいることを発見した3).これとは独立に,2012年,別の研究グループも,ヒトの脳の言語野において同様な構造を発見している4).一方,これまで,皮質下投射細胞マイクロカラムの機能やほかのニューロンの空間的な配置については不明であった.
1.第5層の構造解析
これまでの研究においては脳をスライスして解析していたが,今回は,新しく開発された,脳を丸ごと3次元解析する技術を導入した5-7).神経終末への色素の注入や抗体染色などを用いて個々のニューロンのタイプを可視化し,脳の全体を透明化して3次元撮影することにより,第5層において1個体あたり数千~数万個のニューロンの位置座標を決定した.その結果,マウスの大脳皮質の第5層においては,1)皮質下投射細胞マイクロカラムは視覚野,体性感覚野,運動野といった機能の異なる複数の領野において共通にみられ,ハニカム状の六方格子配列をとり並んでいること,2)皮質投射細胞もマイクロカラムを形成し,この皮質投射細胞マイクロカラムは皮質下投射細胞マイクロカラムとは互い違いに並んでいること,3)パルブアルブミン陽性細胞およびソマトスタチン陽性細胞は皮質下投射細胞マイクロカラムに選択的に含まれ,皮質投射細胞マイクロカラムには含まれないこと,が明らかにされた(図1).
2.マイクロカラムの機能解析
マイクロカラムの機能を調べるため,皮質下投射細胞の神経活動を解析した.ニューロンが発火するとCa2+の濃度が上昇する.そこで,皮質下投射細胞にCa2+の濃度の上昇により蛍光強度が上昇するCa2+センサー8) を導入し,生体脳イメージングを実施した.その結果,同一のマイクロカラムに含まれるニューロンは同期した神経活動を示すことが明らかにされた.この同期した神経活動は視覚野,体性感覚野,運動野の皮質下投射細胞マイクロカラムにおいて共通にみられた.さらに,視覚野においては,同一のマイクロカラムに含まれるニューロンは方位選択性および眼優位性が類似していた.以上より,個々のマイクロカラムはそれぞれ特定の情報を処理しており,大脳皮質の幅広い領野の共通な機能単位として動作することが明らかにされた.
3.マイクロカラムに特異的なギャップ結合およびシナプス結合
電気生理学的な解析により,同じマイクロカラムに含まれるニューロンは同じニューロンからの入力をうけていることが明らかにされ,この入力が同期した神経活動をもたらす可能性が示された.また,大脳皮質において神経回路が形成される時期には,同じマイクロカラムに含まれるニューロンはギャップ結合により電気的に結合していることがわかった.このギャップ結合は,マイクロカラムに特異的な神経回路の形成を誘導する可能性があると考えられた.
おわりに
以上の結果から,大脳皮質の幅広い領域において第5層はマイクロカラムがくり返した神経回路に組織化されていることが示された(図1).個々のマイクロカラムは要素的な情報処理を担う機能モジュールであると考えられ,多数のマイクロカラムによる並列処理が第5層における情報処理を担うことが示された.この神経回路の構造は大脳皮質のさまざまな領野に存在したことから,感覚処理,運動制御,言語処理などの大脳の多様な機能に共通する情報処理を担うと考えられた.
第5層がマイクロカラムのくり返しから構成されていたことから,10個程度のニューロンからなる単一のマイクロカラムの機能が明らかにされれば,第5層の機能について深い理解が得られる可能性がある.さらに,マイクロカラムは大脳皮質のさまざまな領野において共通な機能単位であるため,感覚,運動制御,言語など脳の広範な機能に共通する普遍的な情報処理の機構が明らかにされることが期待される.
今回の研究においては,ニューロンの分類が比較的進んでいる第5層を対象とした.近年,大脳皮質のほかの層においても遺伝子の発現などを指標としたニューロンの分類が進められているため,この情報を用いてマイクロカラム様の構造が探索されることが期待される.実際,ギャップ結合により結合したカラム状のクラスターは第5層のほかにも存在するため9,10),大脳皮質のほかの層にもマイクロカラムが存在する可能性がある.
個々のマイクロカラムに含まれる細胞は特定の方位選択性や眼優位性をもつ.一方,方位選択性カラムや眼優位性カラムなど古典的な皮質カラムは細胞数十個分ほどの幅があり,マイクロカラムより幅広い構造をもつ.したがって,ネコやサルなどにおいては方位選択性や眼優位性の類似した多数のマイクロカラムが並ぶことにより皮質カラムが形成されている可能性がある.もし,このことが示されれば,皮質カラムとそれ以外のさまざまな機能をもつ大脳皮質の領野の統一的な理解が可能になると期待される.
文 献
- Horton, J. C. & Adams, D. L.: The cortical column: a structure without a function. Philos. Trans. R. Soc. Lond. B. Biol. Sci., 360, 837-862 (2005)[PubMed]
- da Costa, N. M. & Martin, K. A.: Whose cortical column would that be? Front. Neuroanat., 4, 16 (2010)[PubMed]
- Maruoka, H., Kubota, K., Kurokawa, R. et al.: Periodic organization of a major subtype of pyramidal neurons in neocortical layer V. J. Neurosci., 31, 18522-18542 (2011)[PubMed]
- Kwan, K. Y., Lam, M. M., Johnson, M. B. et al.: Species-dependent posttranscriptional regulation of NOS1 by FMRP in the developing cerebral cortex. Cell, 149, 899-911 (2012)[PubMed]
- Ke, M. T., Fujimoto, S. & Imai, T.: SeeDB: a simple and morphology-preserving optical clearing agent for neuronal circuit reconstruction. Nat. Neurosci., 16, 1154-1161 (2013)[PubMed]
- Hama, H., Hioki, H., Namiki, K. et al.: ScaleS: an optical clearing palette for biological imaging. Nat. Neurosci., 18, 1518-1529 (2015)[PubMed]
- Susaki, E. A., Tainaka, K., Perrin, D. et al.: Advanced CUBIC protocols for whole-brain and whole-body clearing and imaging. Nat. Protoc., 10, 1709-1727 (2015)[PubMed]
- Ohkura, M., Sasaki, T., Sadakari, J. et al.: Genetically encoded green fluorescent Ca2+ indicators with improved detectability for neuronal Ca2+ signals. PLoS One, 7, e51286 (2012)[PubMed]
- Yuste, R., Peinado, A. & Katz, L. C.: Neuronal domains in developing neocortex. Science, 257, 665-669 (1992)[PubMed]
- Peinado, A., Yuste, R. & Katz, L. C.: Extensive dye coupling between rat neocortical neurons during the period of circuit formation. Neuron, 10, 103-114 (1993)[PubMed]
著者プロフィール
略歴:2006年 大阪大学大学院医学系研究科博士課程 修了,同年より理化学研究所脳科学総合研究センター 博士研究員.
研究テーマ:大脳皮質から出力する神経回路における情報処理の機構.
関心事:マイクロカラムが生体においてどのように機能するのか.
細谷 俊彦(Toshihiko Hosoya)
理化学研究所脳科学総合研究センター チームリーダー.
研究室URL:http://t-hosoya.brain.riken.jp/
© 2017 丸岡久人・細谷俊彦 Licensed under CC 表示 2.1 日本
