運動学習にともなう大脳皮質におけるマクロな神経活動の変化
牧野 浩史
(米国California大学San Diego校Department of Neurosciences)
email:牧野浩史
DOI: 10.7875/first.author.2017.050
Transformation of cortex-wide emergent properties during motor learning.
Hiroshi Makino, Chi Ren, Haixin Liu, An Na Kim, Neehar Kondapaneni, Xin Liu, Duygu Kuzum, Takaki Komiyama
Neuron, 94, 880-890.e8 (2017)
個々のニューロンを理解することからは推測できない脳の全体のふるまいは,学習により変化することが予想される.この研究において,運動学習をしているマウスの大脳皮質におけるマクロな神経活動の変化を広視野Ca2+イメージング法により長期間にわたり測定した.運動の際に大脳皮質のおのおのの領域は順に活性化されたが,運動学習をつうじこの一連の神経活動の速度は上昇しばらつきは低下した.また,運動学習が進むにつれ2次運動野(運動前野)から新たな神経活動の流れが現われ,ほかの領域に影響をおよぼした.さらに,2次運動野の第2/3層および第5層において2光子Ca2+イメージング法により運動学習における興奮性ニューロンの神経活動について調べたところ,とくに第5層において運動の準備段階で神経活動をはじめるニューロンの数が多くなった.これらの結果から,運動学習における大脳皮質のマクロなレベルでの新たな動作の原理が明らかにされた.
脳などの複雑なシステムのふるまいは,個々の要素であるニューロンを完全に理解したとしても容易に予測することはできない.このようなシステムとして起こる挙動は学習により変化することが予想されるが,これまでの技術ではこれを検証することは困難であり解析は進んでいなかった.脳はおのおのの領域が独自の機能をもち,生物が行動する際にはおのおのの領域がそれぞれ活性化されて神経活動の連鎖を形成する1).最近,この脳における情報の流れは課題の種類によりさまざまなパターンを示すことが明らかにされたが2),とくに,げっ歯類におけるマクロなレベルでの学習による変化についてはあまり研究されていなかった.
運動学習とは新たな運動スキルを獲得しその正確性を高めることである.運動学習における脳の可塑性については,たとえば,樹状突起スパイン,単一のニューロン,ニューロンの集合といったさまざまなレベルにおいて解析されてきたが3,4),大脳皮質の全体といったよりマクロなレベルでの研究はまったく進んでいない.この研究においては,広視野Ca2+イメージング法を大脳皮質の大部分に適用することにより,運動学習における大脳皮質の全体にわたる挙動の変化にせまった.
運動学習におけるマウスの大脳皮質の神経活動の変化について調べるため,興奮性ニューロンにおいてCa2+指示薬であるGCaMP6を発現するトランスジェニックマウス5) を用い,広視野Ca2+イメージング法により数週間にわたり神経活動を測定した.そのあいだ,マウスに音の提示されるあいだにレバーを押すと報酬がもらえるというレバー押し運動6) を学習させたところ,2週間後には正答率が90%以上になり,また,レバー押しの正確性も向上した.そして,運動学習により,主成分分析および独立成分分析により区画化された1次運動野および2次運動野(運動前野),体性感覚野,後部頭頂皮質,脳梁膨大後部皮質,視覚野など,それぞれの領域が順に活性化されていくようすが確認された.
運動学習に関連したマクロな脳の挙動の変化を長期的に調べた.マウスがレバー押し運動をするときには脳のおのおのの領域の神経活動がそれぞれの時間的なパターンをともない活性化するが,運動学習により,神経活動のピークに達するまでの速度が変化した.神経活動のピークの半分に達する時間を指標としてその速度を調べたところ,ほとんどの領域においてこの時間が短くなった.また,運動学習のまえに神経活動が遅かった領域は速かった領域に比べより速度が上昇するなど,全体として,一連の神経活動の連鎖に対し時間圧縮が起こった.また,大脳皮質の全体での神経活動のばらつきも徐々に減少した(図1).これらの現象は行動の変化により説明のつくものではなく,脳の可塑性によるものであることもわかった.
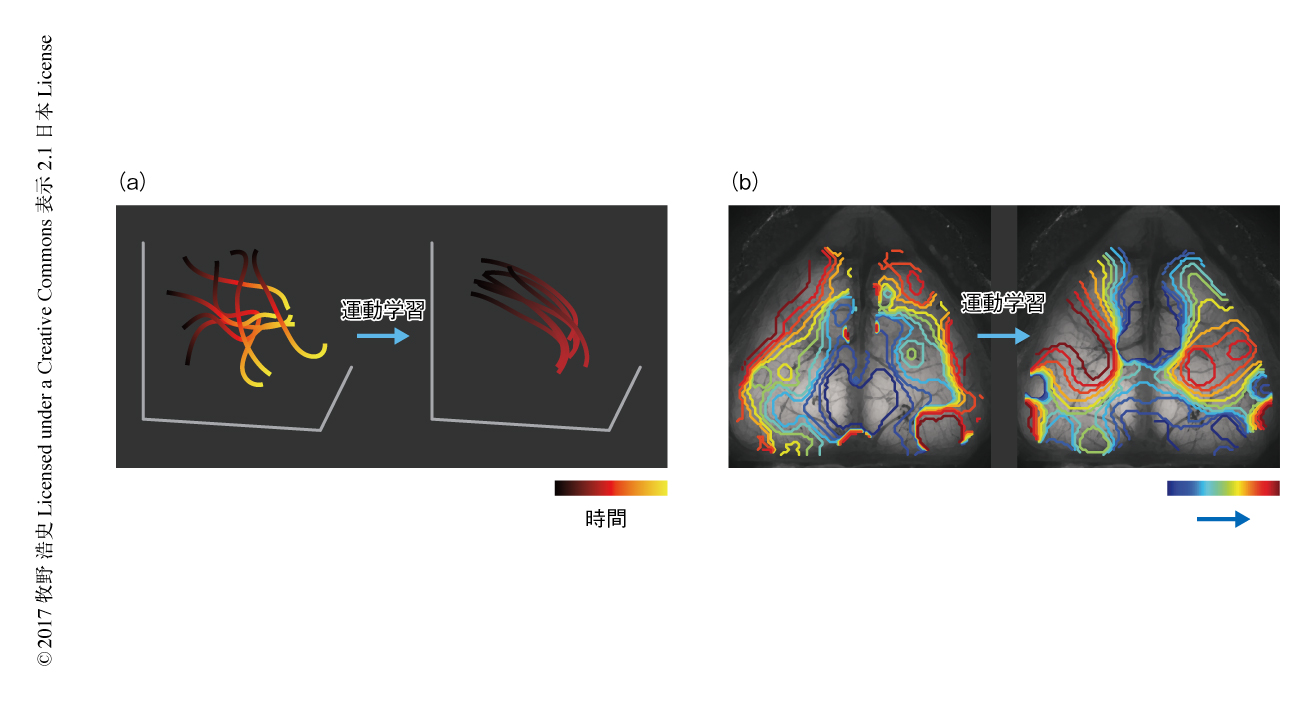
運動学習により大脳皮質における神経活動の伝達するようすがどのように変化するのか調べた.この解析においては,大脳皮質におけるCa2+シグナルを波として扱い7),位相のずれを調べることにより神経活動がどのように伝達するのか検証した.その結果,運動学習のまえには脳梁膨大後部皮質から神経活動が放射状に伝達するようすがみられたが,運動学習が進むにつれ2次運動野から新しい神経活動の流れが現われた(図1).
この神経活動の伝達は領域のあいだの相互作用によるものなのか,それとも,それぞれの領域の独自の神経活動の時間差による結果なのかを調べるため,Granger因果8) により時系列データの因果を推定したところ,運動学習のまえにはそれぞれの領域の因果関係はほとんどなかったのに対し,運動学習により2次運動野からほかの領域への因果関係が現われた.つまり,運動学習により2次運動野がほかの領域への影響を強めることが示された.
運動学習により2次運動野から新たな神経活動の伝達がはじまりほかの領域に影響をおよぼすようになることが明らかにされたが,実際に,2次運動野は運動学習により起こる脳の変化に必要なのかどうかを検証するため,運動学習ののちに2次運動野における神経活動を抑制したところ,マウスの行動および大脳皮質の神経活動のどちらもあらゆる点について運動学習のまえの状態にもどった.具体的には,正答率およびレバー押しの正確性は低下し,大脳皮質の全体にわたる神経活動の連鎖の時間圧縮ももとにもどりそのばらつきも増加した.さらに,神経活動の連鎖のはじまる領域である脳梁膨大後部皮質において同様に神経活動を抑制したところ,マウスの行動および大脳皮質の神経活動のどちらにもあまり変化はみられなかった.これは,脳梁膨大後部皮質からほかの領域への因果関係がほとんどなかったこととも合致した.これらの結果から,2次運動野が運動学習により起こる脳の可塑性に必要であることが示唆された.
2次運動野における個々のニューロンの神経活動について調べるため,興奮性ニューロンにおいてCa2+指示薬であるGCaMP6を発現するトランスジェニックマウス9) を用い,2光子Ca2+イメージング法によりレバー押し運動に関連する第2/3層および5層の興奮性ニューロンの神経活動をそれぞれ別々に2週間にわたり測定した.すると,レバー押し運動の際に,活性化されるニューロンと抑制されるニューロンのあることがわかった.また,とくに第5層においてレバー押し運動の準備段階で神経活動をはじめるニューロンの数が多くなった.さらに,新たに活性化あるいは抑制されるニューロンがもともと活動していたニューロンにとって代わり,全体として神経活動のばらつきも徐々に減少した.これらの結果から,広視野Ca2+イメージング法による結果が支持されたとともに,2次運動野における個々のニューロンの可塑性についても明らかにされた.
これまで,マウスの運動学習についてはシナプスやニューロンといったミクロなレベルでさかんに研究されてきたが,この研究においては,大脳皮質の全体にわたるマクロな挙動の変化について検証した.その結果,神経活動の伝達するようすが変化することがわかった.これは,筆者らが以前に示した,学習による情報の伝達機構の変化10) に類似しており,運動学習は高次機能をつかさどる2次運動野からのトップダウン入力を高めるものであるといえる.今後は,大脳皮質だけでなく,大脳基底核や小脳といったほかの領域との相互作用,あるいは,大脳皮質の全体にわたる神経活動の連鎖の時間圧縮の機構について,さらなる研究が期待される.
略歴:2010年 米国Cold Spring Harbor Laboratory博士課程 修了,2011年 米国California大学San Diego校 博士研究員を経て,2016年よりシンガポールNanyang Technological大学School of Medicine助教授.
研究テーマ:複雑系科学と大脳皮質の動作の原理.
© 2017 牧野 浩史 Licensed under CC 表示 2.1 日本
(米国California大学San Diego校Department of Neurosciences)
email:牧野浩史
DOI: 10.7875/first.author.2017.050
Transformation of cortex-wide emergent properties during motor learning.
Hiroshi Makino, Chi Ren, Haixin Liu, An Na Kim, Neehar Kondapaneni, Xin Liu, Duygu Kuzum, Takaki Komiyama
Neuron, 94, 880-890.e8 (2017)
要 約
個々のニューロンを理解することからは推測できない脳の全体のふるまいは,学習により変化することが予想される.この研究において,運動学習をしているマウスの大脳皮質におけるマクロな神経活動の変化を広視野Ca2+イメージング法により長期間にわたり測定した.運動の際に大脳皮質のおのおのの領域は順に活性化されたが,運動学習をつうじこの一連の神経活動の速度は上昇しばらつきは低下した.また,運動学習が進むにつれ2次運動野(運動前野)から新たな神経活動の流れが現われ,ほかの領域に影響をおよぼした.さらに,2次運動野の第2/3層および第5層において2光子Ca2+イメージング法により運動学習における興奮性ニューロンの神経活動について調べたところ,とくに第5層において運動の準備段階で神経活動をはじめるニューロンの数が多くなった.これらの結果から,運動学習における大脳皮質のマクロなレベルでの新たな動作の原理が明らかにされた.
はじめに
脳などの複雑なシステムのふるまいは,個々の要素であるニューロンを完全に理解したとしても容易に予測することはできない.このようなシステムとして起こる挙動は学習により変化することが予想されるが,これまでの技術ではこれを検証することは困難であり解析は進んでいなかった.脳はおのおのの領域が独自の機能をもち,生物が行動する際にはおのおのの領域がそれぞれ活性化されて神経活動の連鎖を形成する1).最近,この脳における情報の流れは課題の種類によりさまざまなパターンを示すことが明らかにされたが2),とくに,げっ歯類におけるマクロなレベルでの学習による変化についてはあまり研究されていなかった.
運動学習とは新たな運動スキルを獲得しその正確性を高めることである.運動学習における脳の可塑性については,たとえば,樹状突起スパイン,単一のニューロン,ニューロンの集合といったさまざまなレベルにおいて解析されてきたが3,4),大脳皮質の全体といったよりマクロなレベルでの研究はまったく進んでいない.この研究においては,広視野Ca2+イメージング法を大脳皮質の大部分に適用することにより,運動学習における大脳皮質の全体にわたる挙動の変化にせまった.
1.広視野Ca2+イメージング法による大脳皮質におけるマクロな神経活動の測定
運動学習におけるマウスの大脳皮質の神経活動の変化について調べるため,興奮性ニューロンにおいてCa2+指示薬であるGCaMP6を発現するトランスジェニックマウス5) を用い,広視野Ca2+イメージング法により数週間にわたり神経活動を測定した.そのあいだ,マウスに音の提示されるあいだにレバーを押すと報酬がもらえるというレバー押し運動6) を学習させたところ,2週間後には正答率が90%以上になり,また,レバー押しの正確性も向上した.そして,運動学習により,主成分分析および独立成分分析により区画化された1次運動野および2次運動野(運動前野),体性感覚野,後部頭頂皮質,脳梁膨大後部皮質,視覚野など,それぞれの領域が順に活性化されていくようすが確認された.
2.運動学習は大脳皮質のおのおのの領域にわたる神経活動の連鎖の速度を上げばらつきを減らす
運動学習に関連したマクロな脳の挙動の変化を長期的に調べた.マウスがレバー押し運動をするときには脳のおのおのの領域の神経活動がそれぞれの時間的なパターンをともない活性化するが,運動学習により,神経活動のピークに達するまでの速度が変化した.神経活動のピークの半分に達する時間を指標としてその速度を調べたところ,ほとんどの領域においてこの時間が短くなった.また,運動学習のまえに神経活動が遅かった領域は速かった領域に比べより速度が上昇するなど,全体として,一連の神経活動の連鎖に対し時間圧縮が起こった.また,大脳皮質の全体での神経活動のばらつきも徐々に減少した(図1).これらの現象は行動の変化により説明のつくものではなく,脳の可塑性によるものであることもわかった.
3.運動学習の際に2次運動野から新たな神経活動の流れが現われる
運動学習により大脳皮質における神経活動の伝達するようすがどのように変化するのか調べた.この解析においては,大脳皮質におけるCa2+シグナルを波として扱い7),位相のずれを調べることにより神経活動がどのように伝達するのか検証した.その結果,運動学習のまえには脳梁膨大後部皮質から神経活動が放射状に伝達するようすがみられたが,運動学習が進むにつれ2次運動野から新しい神経活動の流れが現われた(図1).
この神経活動の伝達は領域のあいだの相互作用によるものなのか,それとも,それぞれの領域の独自の神経活動の時間差による結果なのかを調べるため,Granger因果8) により時系列データの因果を推定したところ,運動学習のまえにはそれぞれの領域の因果関係はほとんどなかったのに対し,運動学習により2次運動野からほかの領域への因果関係が現われた.つまり,運動学習により2次運動野がほかの領域への影響を強めることが示された.
4.2次運動野は運動学習による大脳皮質の変化に必要である
運動学習により2次運動野から新たな神経活動の伝達がはじまりほかの領域に影響をおよぼすようになることが明らかにされたが,実際に,2次運動野は運動学習により起こる脳の変化に必要なのかどうかを検証するため,運動学習ののちに2次運動野における神経活動を抑制したところ,マウスの行動および大脳皮質の神経活動のどちらもあらゆる点について運動学習のまえの状態にもどった.具体的には,正答率およびレバー押しの正確性は低下し,大脳皮質の全体にわたる神経活動の連鎖の時間圧縮ももとにもどりそのばらつきも増加した.さらに,神経活動の連鎖のはじまる領域である脳梁膨大後部皮質において同様に神経活動を抑制したところ,マウスの行動および大脳皮質の神経活動のどちらにもあまり変化はみられなかった.これは,脳梁膨大後部皮質からほかの領域への因果関係がほとんどなかったこととも合致した.これらの結果から,2次運動野が運動学習により起こる脳の可塑性に必要であることが示唆された.
5.2光子Ca2+イメージング法による個々のニューロンの神経活動の測定
2次運動野における個々のニューロンの神経活動について調べるため,興奮性ニューロンにおいてCa2+指示薬であるGCaMP6を発現するトランスジェニックマウス9) を用い,2光子Ca2+イメージング法によりレバー押し運動に関連する第2/3層および5層の興奮性ニューロンの神経活動をそれぞれ別々に2週間にわたり測定した.すると,レバー押し運動の際に,活性化されるニューロンと抑制されるニューロンのあることがわかった.また,とくに第5層においてレバー押し運動の準備段階で神経活動をはじめるニューロンの数が多くなった.さらに,新たに活性化あるいは抑制されるニューロンがもともと活動していたニューロンにとって代わり,全体として神経活動のばらつきも徐々に減少した.これらの結果から,広視野Ca2+イメージング法による結果が支持されたとともに,2次運動野における個々のニューロンの可塑性についても明らかにされた.
おわりに
これまで,マウスの運動学習についてはシナプスやニューロンといったミクロなレベルでさかんに研究されてきたが,この研究においては,大脳皮質の全体にわたるマクロな挙動の変化について検証した.その結果,神経活動の伝達するようすが変化することがわかった.これは,筆者らが以前に示した,学習による情報の伝達機構の変化10) に類似しており,運動学習は高次機能をつかさどる2次運動野からのトップダウン入力を高めるものであるといえる.今後は,大脳皮質だけでなく,大脳基底核や小脳といったほかの領域との相互作用,あるいは,大脳皮質の全体にわたる神経活動の連鎖の時間圧縮の機構について,さらなる研究が期待される.
文 献
- Abeles, M.: Corticonics: Neural Circuits of the Cerebral Cortex. Cambridge University Press, Cambridge (1991)
- Siegel, M., Buschman, T. J. & Miller, E. K.: Cortical information flow during flexible sensorimotor decisions. Science, 348, 1352-1355 (2015)[PubMed]
- Makino, H., Hwang, E. J., Hedrick, N. G. et al.: Circuit mechanisms of sensorimotor learning. Neuron, 92, 705-721 (2016)[PubMed]
- Peters, A. J., Liu, H. & Komiyama, T.: Learning in the rodent motor cortex. Annu. Rev. Neurosci., DOI: 10.1146/annurev-neuro-072116-031407[PubMed]
- Dana, H., Chen, T. W., Hu, A. et al.: Thy1-GCaMP6 transgenic mice for neuronal population imaging in vivo. PLoS One, 9, e108697 (2014)[PubMed]
- Peters, A. J., Chen, S. X. & Komiyama, T.: Emergence of reproducible spatiotemporal activity during motor learning. Nature, 510, 263-267 (2014)[PubMed]
- Prechtl, J. C., Cohen, L. B., Pesaran, B. et al.: Visual stimuli induce waves of electrical activity in turtle cortex. Proc. Natl. Acad. Sci. USA, 94, 7621-7626 (1997)[PubMed]
- Barnett, L. & Seth, A. K.: The MVGC multivariate Granger causality toolbox: a new approach to Granger-causal inference. J. Neurosci. Methods, 223, 50-68 (2014)[PubMed]
- Wekselblatt, J. B., Flister, E. D., Piscopo, D.M. et al.: Large-scale imaging of cortical dynamics during sensory perception and behavior. J. Neurophysiol., 115, 2852-2866 (2016)[PubMed]
- Makino, H. & Komiyama, T.: Learning enhances the relative impact of top-down processing in the visual cortex. Nat. Neurosci., 18, 1116-1122 (2015)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2010年 米国Cold Spring Harbor Laboratory博士課程 修了,2011年 米国California大学San Diego校 博士研究員を経て,2016年よりシンガポールNanyang Technological大学School of Medicine助教授.
研究テーマ:複雑系科学と大脳皮質の動作の原理.
© 2017 牧野 浩史 Licensed under CC 表示 2.1 日本
