海馬の歯状回に存在する顆粒細胞と苔状細胞の生理学的な特徴および場所情報の表現の違い
千歳雄大・György Buzsáki
(米国New York大学Langone Medical Center,Neuroscience Institute)
email:千歳雄大
DOI: 10.7875/first.author.2017.017
Physiological properties and behavioral correlates of hippocampal granule cells and mossy cells.
Yuta Senzai, György Buzsáki
Neuron, 93, 691-704.e5 (2017)
海馬の歯状回はパターン分離に重要であると考えられている.しかし,この考えを支持するこれまでの生理学的な研究においては,歯状回の顆粒細胞と苔状細胞とが区別されていなかったため解釈に混乱をきたしていた.この研究においては,細胞外電気生理記録法により得られた生理学的な指標にもとづく顆粒細胞と苔状細胞との分類法を確立し,さらに,苔状細胞の光遺伝学的な手法による同定によりこの分類が妥当であることを確認した.その結果,苔状細胞はより高い発火の頻度を示し複数の場所受容野をもつ一方,顆粒細胞はより低い発火の頻度を示し多くは場所受容野をもたないあるいは1つだけもっていた.この顆粒細胞の性質は歯状回におけるパターン分離にとり好都合であったが,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離しか示さなかった.さらに,シナプス結合の推定される顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関を調べたところ,シナプス前の顆粒細胞の場所受容野がシナプス後の苔状細胞へと受け継がれることはまれであった.これらの結果から,顆粒細胞だけでなくその下流の苔状細胞やCA3錐体細胞も含めた神経回路の全体がパターン分離にとり重要であることが示唆された.
ニューヨークの地下鉄の駅は違う駅でもよく似ていることがあるが,そこがどこの駅であるかを区別することは日常生活においてしばしば重要である.このように,外界のわずかな差異を区別する“パターン分離”にとり海馬の歯状回が重要であることが提唱され1,2),いくつかの実験により支持されている3-5).パターン分離においては歯状回のおもな出力細胞である顆粒細胞の疎な活動が重要な役割をはたすと考えられている1,2).しかし,顆粒細胞の発火の頻度や場所情報の表現は疎でないという報告もあり5),歯状回におけるパターン分離の神経回路のレベルでの機構については不明な点も多い.筆者らは,これまでの行動している動物からのin vivoでの生理学的な研究において,歯状回に存在する2種類の興奮性ニューロンがうまく区別されていないことに,この混乱は起因するのではないかと考えた.
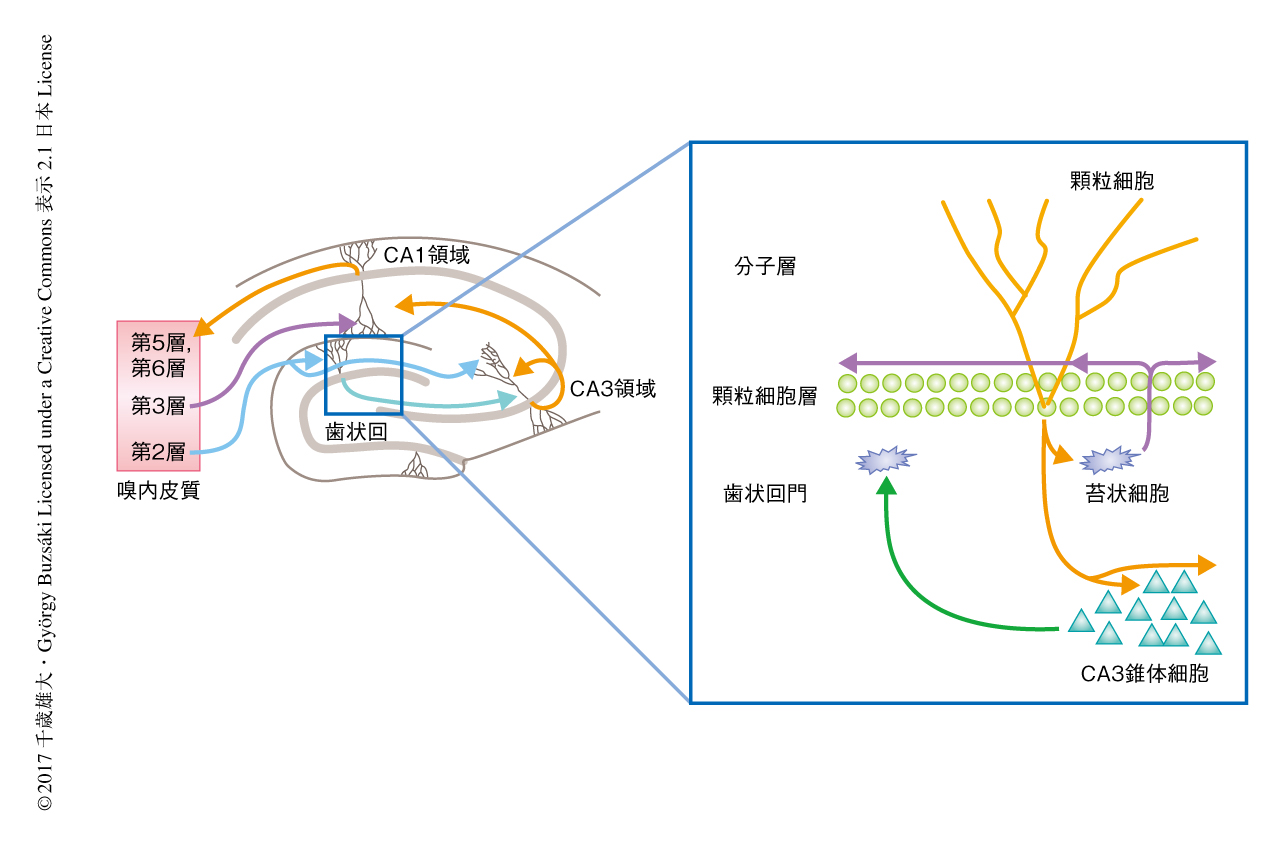
歯状回にはCA3領域への出力を担う興奮性ニューロンである顆粒細胞のほか,苔状細胞とよばれる興奮性ニューロンが存在する.苔状細胞は顆粒細胞から入力をうけ,顆粒細胞へと出力を返し,歯状回において再帰的な回路を形成する6)(図1).苔状細胞は歯状回の機能に重要であることが示されているが7),行動している動物における苔状細胞の挙動や情報の表現についてはほとんどわかっていない.
海馬の歯状回に存在するニューロンを細胞外電気生理記録法により得られた波形およびバースト発火にもとづき興奮性ニューロンと抑制性ニューロンとに分類した.広い波形をもちバースト発火を示すものが興奮性ニューロンと分類されたが,これらをさらに顆粒細胞と苔状細胞とに分類するため,顆粒細胞あるいは苔状細胞の生理学的な特徴から抽出した3つの指標を用いた.1つ目の指標としては,ニューロンの活動が記録されたチャネルにおける2型dentate spikeとよばれる局所の電位パターンの大きさを用いた.2型dentate spikeは歯状回においてノンレム睡眠中などにみられる特徴的な局所の電位パターンであり,その極性が歯状回門ではプラス,分子層ではマイナスとなり,顆粒細胞層において反転することが知られている8).よって,この2型dentate spikeの大きさをもって活動の記録されたニューロンの解剖学的な位置を推定した.2つ目の指標としては,ニューロンの覚醒中の発火の頻度とノンレム睡眠中の発火の頻度の比を用いた.3つ目の指標としては,それぞれのニューロンの波形の特徴をとらえるため,波形の2次導関数の主成分分析から得られた第1主成分および第2主成分の2つを用いた.これら3つの指標から得られた4つのパラメーターにより規定される空間において,歯状回の興奮性ニューロンは2つのクラスターを形成したので,k平均法を用いてこれらを2つのクラスターへと分類した.顆粒細胞は顆粒細胞層に,苔状細胞は歯状回門に存在することが知られていたので,分類されたクラスターのうちプラスのより大きな2型dentate spikeをもつもの,つまり,歯状回門に存在すると推定されるものを苔状細胞,もう一方を顆粒細胞とした.
この生理学的な指標にもとづく分類が妥当であることを示すため,光遺伝学的な手法により苔状細胞を同定した.歯状回においてはドーパミンD2受容体がおもに苔状細胞に特異的に発現し,顆粒細胞においては発現しないことが知られている9).このことを利用し,ドーパミンD2受容体をコードする遺伝子のプロモーターに依存して光感受性プロトンポンプであるアーキロドプシンを発現するマウスを用いて,光遺伝学的な手法により苔状細胞を同定した.その結果,同定された苔状細胞はすべて生理学的な指標にもとづき分類された苔状細胞のクラスターに含まれ,分類の妥当性が確認された.
以上の分類により,顆粒細胞はより低い発火の頻度を示し,とくに覚醒中はノンレム睡眠中と比べ発火の頻度が低下するためにほとんど発火しないことがわかった.一方,苔状細胞はより高い発火の頻度を示し,覚醒中でもその発火の頻度はノンレム睡眠中に比べ低下しなかった.また,顆粒細胞は苔状細胞に比べややとがった波形を示した.これらの生理学的な特徴にくわえて,顆粒細胞から苔状細胞へのシナプス結合を推定した.そのために,ニューロンの発火のタイミングの相互相関ヒストグラムを用い,1.5ミリ秒から5ミリ秒の遅延で有意なピークの認められた場合にシナプス結合があると推定した.その結果,歯状回においては顆粒細胞から苔状細胞への多数のシナプス結合が推定されたが,逆に,苔状細胞から顆粒細胞へのシナプス結合は推定されなかった.また,同じく発火のタイミングの相互相関ヒストグラムを用いた解析において±1ミリ秒の遅延での有意なピークにより検出される同期的な発火については,苔状細胞どうし,および,CA3錐体細胞どうしには認められたが,顆粒細胞どうしには認められなかった.さらに,このCA3錐体細胞の同期的な発火の1ミリ秒以下の遅延と顆粒細胞との距離とのあいだに有意な相関がみられ,歯状回に近いCA3錐体細胞は少しだけ早く発火した.よって,この同期的な発火は顆粒細胞からの共通の入力によるものである可能性が高いと考えられた.これらの結果から,顆粒細胞から苔状細胞およびCA3錐体細胞へのシナプス入力が非常に強いことが示唆された.
顆粒細胞,苔状細胞,CA3錐体細胞それぞれによる場所情報の表現について調べるため,T字迷路課題を遂行しているマウスの歯状回およびCA3領域から細胞外電気生理記録を行った.その結果,顆粒細胞の大半は場所受容野をもたないあるいは1つだけもつことがわかった.一方,大半の苔状細胞は場所受容野をもち多くは2つ以上もっていた.また,場所受容野における最大の発火の頻度も顆粒細胞では低かったが,苔状細胞では高かった.CA3錐体細胞の場所受容野の数は顆粒細胞より多かったが,苔状細胞よりは少なかった.また,T字迷路において右あるいは左に曲がるとき,分岐点よりまえの位置において場所情報の表現が有意に変わるかどうか調べたところ,顆粒細胞,苔状細胞,CA3錐体細胞のいずれにおいても約30%のニューロンにおいて場所情報の表現が分岐点における決断に依存して変化した.
顆粒細胞の低い発火の頻度や疎な場所情報の表現はパターン分離にとり好都合であることから,顆粒細胞が苔状細胞やCA3錐体細胞に比べ強いパターン分離を示すかどうか調べた.そのため,異なるテクスチャーや模様をもつが同じ部屋の同じ場所に置かれた同じサイズの2つの異なる迷路を探索しているマウスのニューロンの活動を記録し,それぞれの迷路における場所情報の表現について調べ,顆粒細胞,苔状細胞,CA3錐体細胞のそれぞれが異なる迷路においてどのように場所受容野を再配置するのか解析した.顆粒細胞,苔状細胞,CA3錐体細胞において2つの迷路における発火の頻度のマップの相関にもとづき場所受容野の再配置の強さを計算したところ,顆粒細胞は苔状細胞やCA3錐体細胞に比べ場所受容野の再配置が起こりにくいことがわかった.個々のニューロンの比較だけではなく,異なる2つの迷路における場所受容野の再配置をニューロンの集団のレベルで定量化して比較するため,それぞれの迷路におけるおのおのの位置においてポピュレーションベクターを構成し,このポピュレーションベクターどうしの相関にもとづき場所受容野の再配置の強さを計算した5).その結果,この方法によっても,顆粒細胞は苔状細胞やCA3錐体細胞に比べ場所受容野の再配置が起こりにくいことが示された.これらの結果から,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離を示すことが示唆された.
顆粒細胞が下流の苔状細胞やCA3錐体細胞に比べ弱いパターン分離を示したことから,苔状細胞やCA3錐体細胞において場所情報はどのように計算されているのか,また,場所受容野は顆粒細胞からの非常に強いシナプス入力をとおして受け継がれるのか,という疑問が生じた.この疑問に答えるため,相互相関ヒストグラムにもとづきシナプス結合の推定された顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関について調べた.その結果,シナプス結合が推定されるにもかかわらず,シナプス前の顆粒細胞の場所受容野とシナプス後の苔状細胞の場所受容野とが相関することはまれであった.また,顆粒細胞から苔状細胞へと場所受容野が受け継がれていることが確認された一例において,別の迷路においては場所受容野は受け継がれないことも確認された.これらの結果から,単独の顆粒細胞から苔状細胞へと場所受容野が受け継がれることはまれであることが示唆された.
この研究においては,細胞外電気生理記録法により得られた生理学的な指標をもとに顆粒細胞と苔状細胞とを分類した.その結果,顆粒細胞は苔状細胞に比べ低い発火の頻度や疎な場所情報の表現を示した.以前の研究において,顆粒細胞は高い発火の頻度や複数の場所受容野をもつと報告されていたが5),これは,苔状細胞の混入による可能性が高いと考えられる.顆粒細胞はそもそもの発火の頻度が低く,さらに,覚醒中にはほとんど発火しないため,覚醒中だけの細胞外電気生理記録により顆粒細胞を検出するのはむずかしい.以前の研究においては,このことにより苔状細胞が顆粒細胞として分類されてしまったと考えられる.
顆粒細胞の低い発火の頻度や疎な場所情報の表現はパターン分離にとり好都合である.しかし,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離しか示さなかった.さらに,シナプス結合の推定される顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関を調べたところ,シナプス前の顆粒細胞の場所受容野がシナプス後の苔状細胞へと受け継がれることはまれであった.これらの結果から,パターン分離は顆粒細胞の段階で完全に行われそれが下流の苔状細胞あるいはCA3錐体細胞へと受け継がれることではないことが示唆され,その機構としていくつかの可能性が考えられる.1つ目として,顆粒細胞から苔状細胞あるいはCA3錐体細胞へと収束するシナプス結合が顆粒細胞により行われたパターン分離を増幅する可能性が考えられる.また,顆粒細胞は歯状回およびCA3領域において多くの抑制性ニューロンを支配することが知られており,苔状細胞やCA3錐体細胞をとりまく抑制性ニューロンがパターン分離の増幅や場所情報の表現の計算に関与する可能性もある.あるいは,パターン分離が歯状回以外の神経回路においても行われる可能性も考えられる.CA3領域は歯状回と同様に嗅内皮質第2層からの直接の入力をうけており,苔状細胞はCA3領域からの投射をうけているという報告もあって10),これらの神経回路がパターン分離に関与する可能性もある.また,このCA3領域から苔状細胞への投射が苔状細胞の位置情報の計算に重要である可能性もある.いずれの場合においても,この研究からは,歯状回におけるパターン分離が顆粒細胞の単独のはたらきだけではなく,その下流の苔状細胞やCA3錐体細胞も含めたネットワークにおいて行われることが示唆された.その詳細な機構に関してさらなる研究が望まれる.
略歴:米国New York大学School of Medicine博士課程 在学中.
関心事:海馬や大脳皮質における神経回路のはたらき.
György Buzsáki
米国New York大学School of Medicine教授.
研究室URL:http://www.buzsakilab.com/
© 2017 千歳雄大・György Buzsáki Licensed under CC 表示 2.1 日本
(米国New York大学Langone Medical Center,Neuroscience Institute)
email:千歳雄大
DOI: 10.7875/first.author.2017.017
Physiological properties and behavioral correlates of hippocampal granule cells and mossy cells.
Yuta Senzai, György Buzsáki
Neuron, 93, 691-704.e5 (2017)
この論文に出現する遺伝子・タンパク質のUniprot ID
要 約
海馬の歯状回はパターン分離に重要であると考えられている.しかし,この考えを支持するこれまでの生理学的な研究においては,歯状回の顆粒細胞と苔状細胞とが区別されていなかったため解釈に混乱をきたしていた.この研究においては,細胞外電気生理記録法により得られた生理学的な指標にもとづく顆粒細胞と苔状細胞との分類法を確立し,さらに,苔状細胞の光遺伝学的な手法による同定によりこの分類が妥当であることを確認した.その結果,苔状細胞はより高い発火の頻度を示し複数の場所受容野をもつ一方,顆粒細胞はより低い発火の頻度を示し多くは場所受容野をもたないあるいは1つだけもっていた.この顆粒細胞の性質は歯状回におけるパターン分離にとり好都合であったが,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離しか示さなかった.さらに,シナプス結合の推定される顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関を調べたところ,シナプス前の顆粒細胞の場所受容野がシナプス後の苔状細胞へと受け継がれることはまれであった.これらの結果から,顆粒細胞だけでなくその下流の苔状細胞やCA3錐体細胞も含めた神経回路の全体がパターン分離にとり重要であることが示唆された.
はじめに
ニューヨークの地下鉄の駅は違う駅でもよく似ていることがあるが,そこがどこの駅であるかを区別することは日常生活においてしばしば重要である.このように,外界のわずかな差異を区別する“パターン分離”にとり海馬の歯状回が重要であることが提唱され1,2),いくつかの実験により支持されている3-5).パターン分離においては歯状回のおもな出力細胞である顆粒細胞の疎な活動が重要な役割をはたすと考えられている1,2).しかし,顆粒細胞の発火の頻度や場所情報の表現は疎でないという報告もあり5),歯状回におけるパターン分離の神経回路のレベルでの機構については不明な点も多い.筆者らは,これまでの行動している動物からのin vivoでの生理学的な研究において,歯状回に存在する2種類の興奮性ニューロンがうまく区別されていないことに,この混乱は起因するのではないかと考えた.
歯状回にはCA3領域への出力を担う興奮性ニューロンである顆粒細胞のほか,苔状細胞とよばれる興奮性ニューロンが存在する.苔状細胞は顆粒細胞から入力をうけ,顆粒細胞へと出力を返し,歯状回において再帰的な回路を形成する6)(図1).苔状細胞は歯状回の機能に重要であることが示されているが7),行動している動物における苔状細胞の挙動や情報の表現についてはほとんどわかっていない.
1.生理学的な指標にもとづく顆粒細胞と苔状細胞との分類および苔状細胞の光遺伝学的な手法による同定
海馬の歯状回に存在するニューロンを細胞外電気生理記録法により得られた波形およびバースト発火にもとづき興奮性ニューロンと抑制性ニューロンとに分類した.広い波形をもちバースト発火を示すものが興奮性ニューロンと分類されたが,これらをさらに顆粒細胞と苔状細胞とに分類するため,顆粒細胞あるいは苔状細胞の生理学的な特徴から抽出した3つの指標を用いた.1つ目の指標としては,ニューロンの活動が記録されたチャネルにおける2型dentate spikeとよばれる局所の電位パターンの大きさを用いた.2型dentate spikeは歯状回においてノンレム睡眠中などにみられる特徴的な局所の電位パターンであり,その極性が歯状回門ではプラス,分子層ではマイナスとなり,顆粒細胞層において反転することが知られている8).よって,この2型dentate spikeの大きさをもって活動の記録されたニューロンの解剖学的な位置を推定した.2つ目の指標としては,ニューロンの覚醒中の発火の頻度とノンレム睡眠中の発火の頻度の比を用いた.3つ目の指標としては,それぞれのニューロンの波形の特徴をとらえるため,波形の2次導関数の主成分分析から得られた第1主成分および第2主成分の2つを用いた.これら3つの指標から得られた4つのパラメーターにより規定される空間において,歯状回の興奮性ニューロンは2つのクラスターを形成したので,k平均法を用いてこれらを2つのクラスターへと分類した.顆粒細胞は顆粒細胞層に,苔状細胞は歯状回門に存在することが知られていたので,分類されたクラスターのうちプラスのより大きな2型dentate spikeをもつもの,つまり,歯状回門に存在すると推定されるものを苔状細胞,もう一方を顆粒細胞とした.
この生理学的な指標にもとづく分類が妥当であることを示すため,光遺伝学的な手法により苔状細胞を同定した.歯状回においてはドーパミンD2受容体がおもに苔状細胞に特異的に発現し,顆粒細胞においては発現しないことが知られている9).このことを利用し,ドーパミンD2受容体をコードする遺伝子のプロモーターに依存して光感受性プロトンポンプであるアーキロドプシンを発現するマウスを用いて,光遺伝学的な手法により苔状細胞を同定した.その結果,同定された苔状細胞はすべて生理学的な指標にもとづき分類された苔状細胞のクラスターに含まれ,分類の妥当性が確認された.
2.顆粒細胞から苔状細胞へのシナプス結合の推定および顆粒細胞とCA3錐体細胞との同期的な発火
以上の分類により,顆粒細胞はより低い発火の頻度を示し,とくに覚醒中はノンレム睡眠中と比べ発火の頻度が低下するためにほとんど発火しないことがわかった.一方,苔状細胞はより高い発火の頻度を示し,覚醒中でもその発火の頻度はノンレム睡眠中に比べ低下しなかった.また,顆粒細胞は苔状細胞に比べややとがった波形を示した.これらの生理学的な特徴にくわえて,顆粒細胞から苔状細胞へのシナプス結合を推定した.そのために,ニューロンの発火のタイミングの相互相関ヒストグラムを用い,1.5ミリ秒から5ミリ秒の遅延で有意なピークの認められた場合にシナプス結合があると推定した.その結果,歯状回においては顆粒細胞から苔状細胞への多数のシナプス結合が推定されたが,逆に,苔状細胞から顆粒細胞へのシナプス結合は推定されなかった.また,同じく発火のタイミングの相互相関ヒストグラムを用いた解析において±1ミリ秒の遅延での有意なピークにより検出される同期的な発火については,苔状細胞どうし,および,CA3錐体細胞どうしには認められたが,顆粒細胞どうしには認められなかった.さらに,このCA3錐体細胞の同期的な発火の1ミリ秒以下の遅延と顆粒細胞との距離とのあいだに有意な相関がみられ,歯状回に近いCA3錐体細胞は少しだけ早く発火した.よって,この同期的な発火は顆粒細胞からの共通の入力によるものである可能性が高いと考えられた.これらの結果から,顆粒細胞から苔状細胞およびCA3錐体細胞へのシナプス入力が非常に強いことが示唆された.
3.顆粒細胞,苔状細胞,CA3錐体細胞それぞれによる場所情報の表現
顆粒細胞,苔状細胞,CA3錐体細胞それぞれによる場所情報の表現について調べるため,T字迷路課題を遂行しているマウスの歯状回およびCA3領域から細胞外電気生理記録を行った.その結果,顆粒細胞の大半は場所受容野をもたないあるいは1つだけもつことがわかった.一方,大半の苔状細胞は場所受容野をもち多くは2つ以上もっていた.また,場所受容野における最大の発火の頻度も顆粒細胞では低かったが,苔状細胞では高かった.CA3錐体細胞の場所受容野の数は顆粒細胞より多かったが,苔状細胞よりは少なかった.また,T字迷路において右あるいは左に曲がるとき,分岐点よりまえの位置において場所情報の表現が有意に変わるかどうか調べたところ,顆粒細胞,苔状細胞,CA3錐体細胞のいずれにおいても約30%のニューロンにおいて場所情報の表現が分岐点における決断に依存して変化した.
4.顆粒細胞,苔状細胞,CA3錐体細胞それぞれのパターン分離への関与
顆粒細胞の低い発火の頻度や疎な場所情報の表現はパターン分離にとり好都合であることから,顆粒細胞が苔状細胞やCA3錐体細胞に比べ強いパターン分離を示すかどうか調べた.そのため,異なるテクスチャーや模様をもつが同じ部屋の同じ場所に置かれた同じサイズの2つの異なる迷路を探索しているマウスのニューロンの活動を記録し,それぞれの迷路における場所情報の表現について調べ,顆粒細胞,苔状細胞,CA3錐体細胞のそれぞれが異なる迷路においてどのように場所受容野を再配置するのか解析した.顆粒細胞,苔状細胞,CA3錐体細胞において2つの迷路における発火の頻度のマップの相関にもとづき場所受容野の再配置の強さを計算したところ,顆粒細胞は苔状細胞やCA3錐体細胞に比べ場所受容野の再配置が起こりにくいことがわかった.個々のニューロンの比較だけではなく,異なる2つの迷路における場所受容野の再配置をニューロンの集団のレベルで定量化して比較するため,それぞれの迷路におけるおのおのの位置においてポピュレーションベクターを構成し,このポピュレーションベクターどうしの相関にもとづき場所受容野の再配置の強さを計算した5).その結果,この方法によっても,顆粒細胞は苔状細胞やCA3錐体細胞に比べ場所受容野の再配置が起こりにくいことが示された.これらの結果から,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離を示すことが示唆された.
5.シナプス結合の推定される顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関
顆粒細胞が下流の苔状細胞やCA3錐体細胞に比べ弱いパターン分離を示したことから,苔状細胞やCA3錐体細胞において場所情報はどのように計算されているのか,また,場所受容野は顆粒細胞からの非常に強いシナプス入力をとおして受け継がれるのか,という疑問が生じた.この疑問に答えるため,相互相関ヒストグラムにもとづきシナプス結合の推定された顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関について調べた.その結果,シナプス結合が推定されるにもかかわらず,シナプス前の顆粒細胞の場所受容野とシナプス後の苔状細胞の場所受容野とが相関することはまれであった.また,顆粒細胞から苔状細胞へと場所受容野が受け継がれていることが確認された一例において,別の迷路においては場所受容野は受け継がれないことも確認された.これらの結果から,単独の顆粒細胞から苔状細胞へと場所受容野が受け継がれることはまれであることが示唆された.
おわりに
この研究においては,細胞外電気生理記録法により得られた生理学的な指標をもとに顆粒細胞と苔状細胞とを分類した.その結果,顆粒細胞は苔状細胞に比べ低い発火の頻度や疎な場所情報の表現を示した.以前の研究において,顆粒細胞は高い発火の頻度や複数の場所受容野をもつと報告されていたが5),これは,苔状細胞の混入による可能性が高いと考えられる.顆粒細胞はそもそもの発火の頻度が低く,さらに,覚醒中にはほとんど発火しないため,覚醒中だけの細胞外電気生理記録により顆粒細胞を検出するのはむずかしい.以前の研究においては,このことにより苔状細胞が顆粒細胞として分類されてしまったと考えられる.
顆粒細胞の低い発火の頻度や疎な場所情報の表現はパターン分離にとり好都合である.しかし,顆粒細胞はその下流に位置する苔状細胞やCA3錐体細胞に比べ弱いパターン分離しか示さなかった.さらに,シナプス結合の推定される顆粒細胞と苔状細胞とのあいだの場所情報の表現の相関を調べたところ,シナプス前の顆粒細胞の場所受容野がシナプス後の苔状細胞へと受け継がれることはまれであった.これらの結果から,パターン分離は顆粒細胞の段階で完全に行われそれが下流の苔状細胞あるいはCA3錐体細胞へと受け継がれることではないことが示唆され,その機構としていくつかの可能性が考えられる.1つ目として,顆粒細胞から苔状細胞あるいはCA3錐体細胞へと収束するシナプス結合が顆粒細胞により行われたパターン分離を増幅する可能性が考えられる.また,顆粒細胞は歯状回およびCA3領域において多くの抑制性ニューロンを支配することが知られており,苔状細胞やCA3錐体細胞をとりまく抑制性ニューロンがパターン分離の増幅や場所情報の表現の計算に関与する可能性もある.あるいは,パターン分離が歯状回以外の神経回路においても行われる可能性も考えられる.CA3領域は歯状回と同様に嗅内皮質第2層からの直接の入力をうけており,苔状細胞はCA3領域からの投射をうけているという報告もあって10),これらの神経回路がパターン分離に関与する可能性もある.また,このCA3領域から苔状細胞への投射が苔状細胞の位置情報の計算に重要である可能性もある.いずれの場合においても,この研究からは,歯状回におけるパターン分離が顆粒細胞の単独のはたらきだけではなく,その下流の苔状細胞やCA3錐体細胞も含めたネットワークにおいて行われることが示唆された.その詳細な機構に関してさらなる研究が望まれる.
文 献
- Marr, D.: Simple memory: a theory for archicortex. Philos. Trans. R. Soc. Lond. B Biol. Sci., 262, 23-81 (1971)[PubMed]
- McNaughton, B. L. & Morris, R. G. M.: Hippocampal synaptic enhancement and information storage within a distributed memory system. Trends Neurosci., 10, 408-415 (1987)
- Gilbert, P. E., Kesner, R. P. & Lee, I.: Dissociating hippocampal subregions: double dissociation between dentate gyrus and CA1. Hippocampus, 11, 626-636 (2001)[PubMed]
- McHugh, T. J., Jones, M. W., Quinn, J. J. et al.: Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science, 317, 94-99 (2007)[PubMed]
- Leutgeb, J. K., Leutgeb, S., Moser, M. et al.: Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science, 315, 961-966 (2007)[PubMed]
- Buckmaster, P. S., Wenzel, H. J., Kunkel, D. D. et al.: Axon arbors and synaptic connections of hippocampal mossy cells in the rat in vivo. J. Comp. Neurol., 366, 271-292 (1996)[PubMed]
- Jinde, S., Zsiros, V., Jiang, Z. et al.: Hilar mossy cell degeneration causes transient dentate granule cell hyperexcitability and impaired pattern separation. Neuron, 76, 1189-1200 (2012)[PubMed]
- Bragin, A., Jando, G., Nadasdy, Z. et al.: Dentate EEG spikes and associated interneuronal population bursts in the hippocampal hilar region of the rat. J. Neurophysiol., 73, 1691-1705 (1995)[PubMed]
- Puighermanal, E., Biever, A., Espallergues, J. et al.: drd2-cre:ribotag mouse line unravels the possible diversity of dopamine d2 receptor-expressing cells of the dorsal mouse hippocampus. Hippocampus, 25, 858-875 (2015)[PubMed]
- Scharfman, H. E.: Evidence from simultaneous intracellular recordings in rat hippocampal slices that area CA3 pyramidal cells innervate dentate hilar mossy cells. J. Neurophysiol., 72, 2167-2180 (1994)[PubMed]
著者プロフィール
略歴:米国New York大学School of Medicine博士課程 在学中.
関心事:海馬や大脳皮質における神経回路のはたらき.
György Buzsáki
米国New York大学School of Medicine教授.
研究室URL:http://www.buzsakilab.com/
© 2017 千歳雄大・György Buzsáki Licensed under CC 表示 2.1 日本
