共生細菌が昆虫の体色を変える
土田 努
(理化学研究所基幹研究所 松本分子昆虫学研究室)
email:土田 努
DOI: 10.7875/first.author.2010.061
Symbiotic bacterium modifies aphid body color.
Tsutomu Tsuchida, Ryuichi Koga, Mitsuyo Horikawa, Tetsuto Tsunoda, Takashi Maoka, Shogo Matsumoto, Jean-Christophe Simon, Takema Fukatsu
Science, 330, 1102-1104 (2010)
動物の体色は自然界において多彩な機能を担うきわめて重要な性質である.欧米に分布するエンドウヒゲナガアブラムシでは同種内に赤色と緑色の体色をもつものが存在し,それらは捕食者あるいは寄生者からの逃れやすさが異なることが知られていた.筆者らは,このアブラムシから新規の共生細菌Rickettsiellaを発見し,その感染により赤色のアブラムシが緑色に変化することを見い出した.種々の実験により,Rickettsiellaはアブラムシの成長や繁殖にはほとんど影響をあたえることなくアブラムシの緑色系色素の生産を活性化させることが示唆された.Rickettsiellaによるアブラムシの体色変化の影響は単に感染個体のみにとどまらず,生態系におけるアブラムシの捕食-被食関係をも変化させうるものである.
多くの動物は色覚をもち,色を手がかりとして環境,住処,食べ物,天敵,競争者,配偶者を認識している.すなわち,動物の体色は種の認識,配偶者をめぐる競争,天敵に対する隠蔽色・警告色・擬態など,生態的に重要な性質を反映した結果であると考えられている.このように生態的にも重要な役割をはたす体色は,当然のことながら,その生物自体の性質であると考えられてきた.
アブラムシ類は世界で4000種以上が知られており,多くの重要な農業害虫種を含んでいるが,同一種内にさまざまな体色をもつものも少なくない.たとえば,欧米では赤色と緑色のエンドウヒゲナガアブラムシが同一の地域に分布している.なぜさまざまな体色が同種内で進化してきたのかという問題については,これまでおもに生態学的な観点から多くの研究が行われてきた.
アブラムシの主要な捕食者にはテントウムシや寄生バチがいるが,テントウムシは緑の植物のうえでめだつ赤色のアブラムシをより高頻度で補食し1),寄生バチは緑色のアブラムシに高頻度で産卵する2),という習性がある.したがって,テントウムシが多い環境では緑色のアブラムシが生存に有利になり,逆に,寄生バチが多い環境では赤色のアブラムシが有利となる.このように,異なる捕食者がそれぞれ異なる色彩への選好性を示すことにより,生存に有利なアブラムシの体色は1種類だけに固定されず複数種類の体色が進化・維持されてきたのだと考えられている1).
古くからの遺伝学的な研究によってエンドウヒゲナガアブラムシの体色は赤色が緑色に対して優性であることが知られていた3).また,最近になり発表されたエンドウヒゲナガアブラムシの概要ゲノム配列4) に基づいた研究の結果,驚いたことに,菌類から遺伝子水平転移によって獲得されたカロテノイド不飽和化酵素がアブラムシの体色を決めていることが判明した5).つまり,赤色系カロテノイド色素を合成するこの遺伝子をもつアブラムシは赤色になるが,この遺伝子を欠くアブラムシは赤色カロテノイドを合成できないため緑色を呈するのである.
約4400種のアブラムシ類のほとんどには体内に菌細胞(mycetocyte,または,bacteriocyte)とよばれる肥大化した細胞が存在し,その細胞質にBuchnera aphidicola(以下,属名のみでBuchneraと記載)という共生細菌を生育させている.Buchneraは,宿主のアブラムシが生涯唯一の餌とする植物師管液に不足する必須アミノ酸などを合成・供給するため,アブラムシの生存・繁殖に不可欠な存在である.Buchneraは卵巣で胚に感染することで次世代へと伝えられていく(このような親から子へと伝わる様式を“垂直伝播”とよぶ).その共生の歴史は1億年以上まえにさかのぼるものと推定されている6).
Buchneraにくわえて,多くのアブラムシでは同一体内に1~2種類の別の種類の細菌が共存しているのが観察される.これまで,エンドウヒゲナガアブラムシからは総計6種類の任意共生細菌(Serratia symbiotica,Hamiltonella defensa,Regiella insecticola,Rickettsia sp.,Spiroplasma sp.,PAXS,以下,属名のみで記載)の存在が報告されてきた7).これらはBuchneraとは異なる細胞や体液に存在しており,Buchneraと同様に卵巣の胚に感染することで次世代に安定して伝播されていく.近年,これらの任意共生細菌はアブラムシの生存には必須でないものの,アブラムシの環境適応にさまざまな影響をあたえることが明らかにされてきた.たとえば,感染によりアブラムシに高温耐性を賦与する共生細菌Serratia 8),アブラムシに寄生バチへの抵抗性をもたらす共生細菌HamiltonellaおよびSerratia 9),アブラムシの植物適応や病原菌耐性にかかわる共生細菌Regiella 10,11),などが報告されている.しかし,共生微生物が正常個体の体色を大きく変えてしまうという報告はこれまで皆無であった.
ヨーロッパ野外集団からアブラムシ系統を収集していた筆者らは,いくつかの系統では緑色の母虫から赤色の幼虫が産み出されるのを発見した.これらの系統の体色は成長するにつれてだんだん緑色っぽくなり,4令幼虫から成虫にいたると完全に緑色になった.この体色の変わるアブラムシの体液を赤色系統のアブラムシに導入したところ,体色が緑に変化する個体が現れた(未発表データ).このことは,アブラムシの体液に存在するなんらかの因子が体色を変えていることを示唆していた.そこで,体液に存在する任意共生細菌が体色変化の要因である可能性を検証した.
体色の変化するアブラムシ系統の体内に生息する共生細菌群集を細菌16S rRNA遺伝子塩基配列にもとづいて解析したところ,生存に必須な共生細菌Buchnera以外に2種類の共生細菌が検出された.ひとつは既知の共生細菌HamiltonellaもしくはSerratiaで,アブラムシ系統によっていずれかが感染していた.さらに,これらの系統すべてにこれまでアブラムシからは報告のなかったRickettsiella属の共生細菌が共感染していた.Rickettsiellaはほかの昆虫類では病原細菌として知られ,進化的にヒトのQ熱病原体(Coxiella)やレジオネラ症病原体(Legionella)と近縁である.
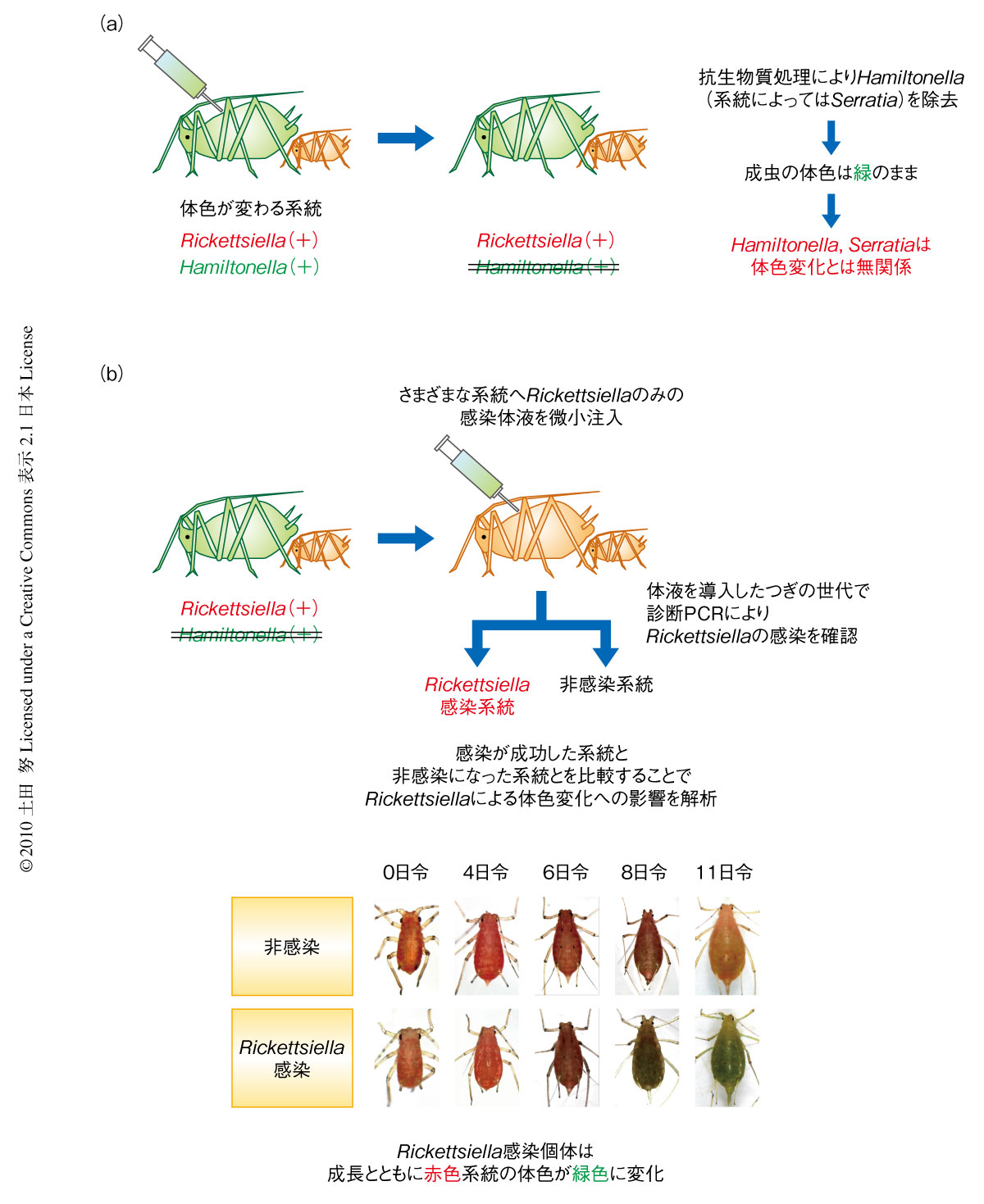
共存する共生細菌のうちいずれが体色の変化にかかわっているのかを調べるため,抗生物質を処理して共生細菌の除去を試みた(図1a).その結果,HamiltonellaおよびSerratiaを除去することに成功したが,これらの共生細菌を除去しても成虫の体色は緑色のままであった.つぎに,Rickettsiellaに感染しているアブラムシ系統の体液を採取して非感染系統に微小注入しその子孫を個別飼育することで,遺伝的な背景は同一でありながらRickettsiella感染あるいは非感染だけが異なる多数のアブラムシ系統を作製した(図1b).すると,Rickettsiellaの感染した赤色系統ではすべて体色が緑色に変化した(図1b).もともと緑色だった系統ではRickettsiellaを感染させても特段の変化はみられなかった.定量PCRを用いた解析の結果は,アブラムシ体内のRickettsiellaの感染密度が高くなるほど体色が緑色に変化するという有意な関係を示した.これらの結果は,Rickettsiellaがエンドウヒゲナガアブラムシの野外集団における緑色個体の形成にかかわっていることを示していた.
さきに述べたような方法で3系統のアブラムシについてRickettsiella感染個体と非感染個体とを作製し,体重,成長速度,産子数,寿命を比較した.その結果,感染による有意な違いはほとんど検出されなかった.
蛍光in situハイブリダイゼーションや電子顕微鏡による観察の結果,Rickettsiellaはアブラムシのほかの任意共生細菌と同様に,アブラムシ体内の二次菌細胞や鞘細胞,体液中に存在していることが判明した.
ヨーロッパ野外集団に由来するエンドウヒゲナガアブラムシ353個体につき診断PCR法を用いて調べたところ,28個体(7.9%)がRickettsiellaに感染しており,自然界における広範な分布が判明した.
自然界における体色とRickettsiella感染との関係を調べると,緑色の個体がすべてRickettsiellaに感染しているわけではなく,また,Rickettsiellaに感染していても赤色のままの個体も存在していた.このことは,体色はアブラムシとRickettsiellaとの相互作用によって決まることを意味していた.
アブラムシの体色はおもに,黄色~赤色のカロテノイド系色素と,緑色~青色などさまざまな色のaphin(または,aphinin)と総称される多環式キノン系色素から構成されている12).Rickettsiellaに感染したアブラムシ(緑色)と非感染のアブラムシ(赤色)の色素分析を行ったところ,カロテノイド系色素の組成や量には大きな違いはみられなかった.また,さきに述べたように,アブラムシの赤色カロテノイドの合成をつかさどるカロテノイド不飽和化酵素の遺伝子発現を定量RT-PCR法により解析した結果からも,Rickettsiella感染が赤色カロテノイドの合成に変化をもたらしているという証拠は得られなかった.一方,多環式キノン配糖体であると推定される緑色色素を逆相薄相クロマトグラフィーにより解析した結果,新規な成分の出現や組成の変化はないものの,その量が感染アブラムシでは非感染アブラムシの3倍以上に増加していた.すなわち,Rickettsiellaが感染することで宿主のアブラムシの緑色色素の生産がなんらかのかたちで活性化され,その結果,体色の変化が生じているものと推察された.
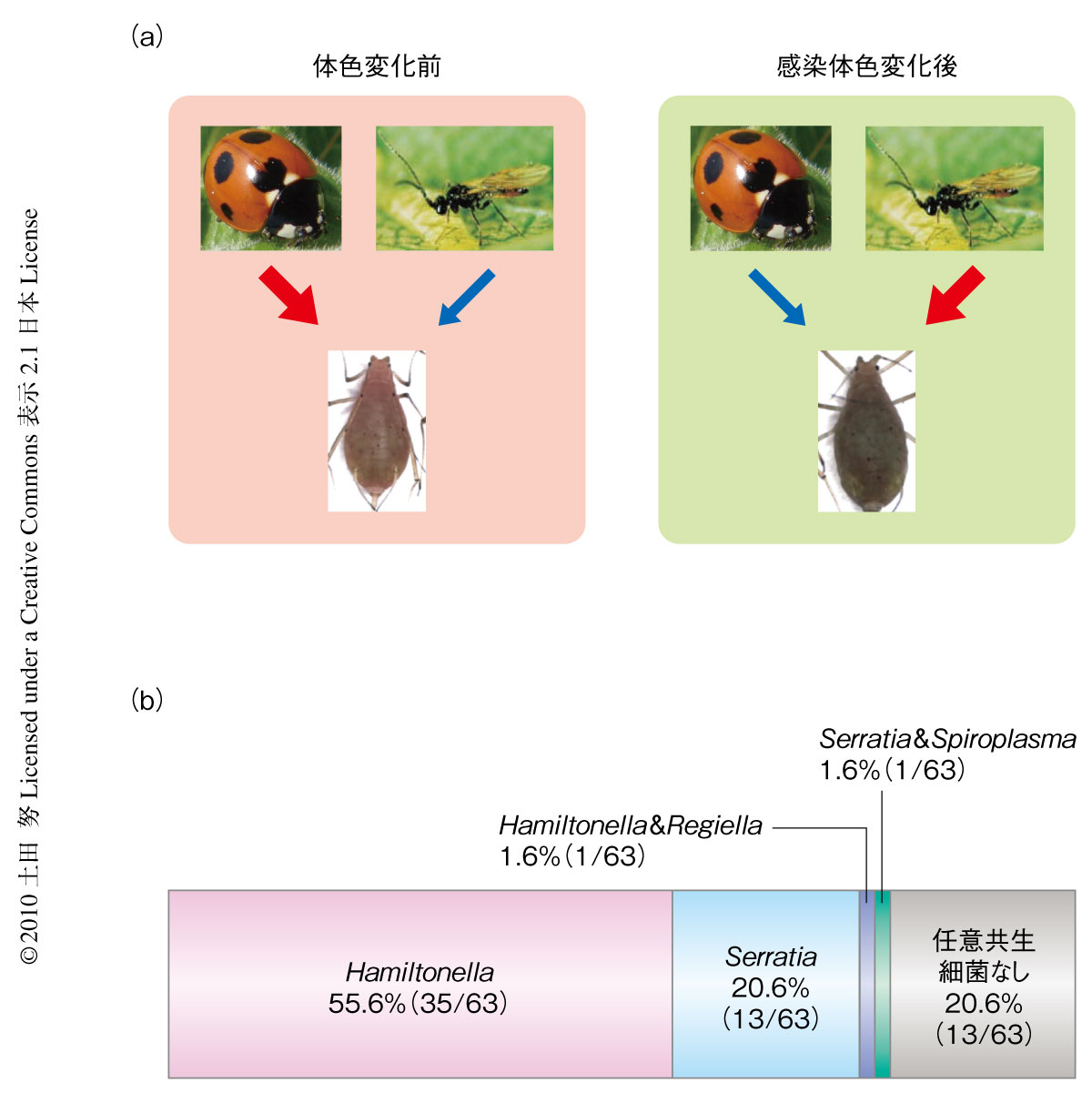
はじめに述べたように,アブラムシの体色は捕食あるいは寄生の回避に重要な役割をはたしている.ゆえに,Rickettsiellaに感染して体色が赤色から緑色に変化すると,テントウムシには食べられにくくなる一方,寄生バチの攻撃はうけやすくなることが予想された1,2)(図2a).興味深いことに,Rickettsiellaに感染しているアブラムシの大部分(約80%)はHamiltonellaもしくはSerratiaという共生細菌にも感染していた(図2b).HamiltonellaやSerratiaは産みつけられた卵や幼虫を殺すことにより寄生バチに対する耐性を賦与する共生細菌として知られているのはさきに述べたとおりである9).このことは,Rickettsiellaはアブラムシの体色を緑色に変えてテントウムシに補食されにくくすると同時に,(緑色のアブラムシに好んで産卵する)寄生バチへの耐性をあたえる共生細菌と共感染することにより宿主アブラムシの生存率,ひいては,自らの生存率を上げている可能性を示唆していた.
この研究では,自然界においてさまざまな役割をはたしている“生物の体の色”に体内の共生微生物が大きく影響している場合のあることを世界ではじめて実証した.この研究成果は生物の生態や環境適応の理解に新たな観点を提示するものといえよう.
現在,Rickettsiellaの感染によって生産が活性化される緑色色素の化学構造の詳細な分析とともに,体色変化をひき起こすRickettsiellaの全ゲノム配列の解読,さらには,感染によって活性化(あるいは,抑制)される宿主アブラムシの遺伝子の網羅的な発現解析などに取り組んでいる.この興味深い生命現象を題材にした研究の進展により,生命原理に関する新たな知見の得られることがおおいに期待される.また,これらの解析で明らかになった色素生産の活性化の機構を応用することで,将来的には,色素の効率的な生産方法や生物の色彩や外観を制御するための新規技術の開発につながる可能性もある.
略歴:2003年 東京大学大学院総合文化研究科 修了,同年 英国York大学 博士研究員,2004年 産業技術総合研究所生物機能工学研究部門 博士研究員を経て,2008年より理化学研究所基幹研究所 基礎科学特別研究員.
研究テーマ:昆虫と共生微生物の生物間相互作用を成立させる生理・分子機構の解明,共生微生物が生態系ではたす役割の解明.
抱負:さまざまな手法を有機的に組み合わせた解析により,生物の体内に住む小さな共生微生物の大きな役割を包括的に明らかにすることをめざしています.
© 2010 土田 努 Licensed under CC 表示 2.1 日本
(理化学研究所基幹研究所 松本分子昆虫学研究室)
email:土田 努
DOI: 10.7875/first.author.2010.061
Symbiotic bacterium modifies aphid body color.
Tsutomu Tsuchida, Ryuichi Koga, Mitsuyo Horikawa, Tetsuto Tsunoda, Takashi Maoka, Shogo Matsumoto, Jean-Christophe Simon, Takema Fukatsu
Science, 330, 1102-1104 (2010)
要 約
動物の体色は自然界において多彩な機能を担うきわめて重要な性質である.欧米に分布するエンドウヒゲナガアブラムシでは同種内に赤色と緑色の体色をもつものが存在し,それらは捕食者あるいは寄生者からの逃れやすさが異なることが知られていた.筆者らは,このアブラムシから新規の共生細菌Rickettsiellaを発見し,その感染により赤色のアブラムシが緑色に変化することを見い出した.種々の実験により,Rickettsiellaはアブラムシの成長や繁殖にはほとんど影響をあたえることなくアブラムシの緑色系色素の生産を活性化させることが示唆された.Rickettsiellaによるアブラムシの体色変化の影響は単に感染個体のみにとどまらず,生態系におけるアブラムシの捕食-被食関係をも変化させうるものである.
はじめに
多くの動物は色覚をもち,色を手がかりとして環境,住処,食べ物,天敵,競争者,配偶者を認識している.すなわち,動物の体色は種の認識,配偶者をめぐる競争,天敵に対する隠蔽色・警告色・擬態など,生態的に重要な性質を反映した結果であると考えられている.このように生態的にも重要な役割をはたす体色は,当然のことながら,その生物自体の性質であると考えられてきた.
アブラムシ類は世界で4000種以上が知られており,多くの重要な農業害虫種を含んでいるが,同一種内にさまざまな体色をもつものも少なくない.たとえば,欧米では赤色と緑色のエンドウヒゲナガアブラムシが同一の地域に分布している.なぜさまざまな体色が同種内で進化してきたのかという問題については,これまでおもに生態学的な観点から多くの研究が行われてきた.
アブラムシの主要な捕食者にはテントウムシや寄生バチがいるが,テントウムシは緑の植物のうえでめだつ赤色のアブラムシをより高頻度で補食し1),寄生バチは緑色のアブラムシに高頻度で産卵する2),という習性がある.したがって,テントウムシが多い環境では緑色のアブラムシが生存に有利になり,逆に,寄生バチが多い環境では赤色のアブラムシが有利となる.このように,異なる捕食者がそれぞれ異なる色彩への選好性を示すことにより,生存に有利なアブラムシの体色は1種類だけに固定されず複数種類の体色が進化・維持されてきたのだと考えられている1).
古くからの遺伝学的な研究によってエンドウヒゲナガアブラムシの体色は赤色が緑色に対して優性であることが知られていた3).また,最近になり発表されたエンドウヒゲナガアブラムシの概要ゲノム配列4) に基づいた研究の結果,驚いたことに,菌類から遺伝子水平転移によって獲得されたカロテノイド不飽和化酵素がアブラムシの体色を決めていることが判明した5).つまり,赤色系カロテノイド色素を合成するこの遺伝子をもつアブラムシは赤色になるが,この遺伝子を欠くアブラムシは赤色カロテノイドを合成できないため緑色を呈するのである.
約4400種のアブラムシ類のほとんどには体内に菌細胞(mycetocyte,または,bacteriocyte)とよばれる肥大化した細胞が存在し,その細胞質にBuchnera aphidicola(以下,属名のみでBuchneraと記載)という共生細菌を生育させている.Buchneraは,宿主のアブラムシが生涯唯一の餌とする植物師管液に不足する必須アミノ酸などを合成・供給するため,アブラムシの生存・繁殖に不可欠な存在である.Buchneraは卵巣で胚に感染することで次世代へと伝えられていく(このような親から子へと伝わる様式を“垂直伝播”とよぶ).その共生の歴史は1億年以上まえにさかのぼるものと推定されている6).
Buchneraにくわえて,多くのアブラムシでは同一体内に1~2種類の別の種類の細菌が共存しているのが観察される.これまで,エンドウヒゲナガアブラムシからは総計6種類の任意共生細菌(Serratia symbiotica,Hamiltonella defensa,Regiella insecticola,Rickettsia sp.,Spiroplasma sp.,PAXS,以下,属名のみで記載)の存在が報告されてきた7).これらはBuchneraとは異なる細胞や体液に存在しており,Buchneraと同様に卵巣の胚に感染することで次世代に安定して伝播されていく.近年,これらの任意共生細菌はアブラムシの生存には必須でないものの,アブラムシの環境適応にさまざまな影響をあたえることが明らかにされてきた.たとえば,感染によりアブラムシに高温耐性を賦与する共生細菌Serratia 8),アブラムシに寄生バチへの抵抗性をもたらす共生細菌HamiltonellaおよびSerratia 9),アブラムシの植物適応や病原菌耐性にかかわる共生細菌Regiella 10,11),などが報告されている.しかし,共生微生物が正常個体の体色を大きく変えてしまうという報告はこれまで皆無であった.
1.体色の変わるアブラムシの発見
ヨーロッパ野外集団からアブラムシ系統を収集していた筆者らは,いくつかの系統では緑色の母虫から赤色の幼虫が産み出されるのを発見した.これらの系統の体色は成長するにつれてだんだん緑色っぽくなり,4令幼虫から成虫にいたると完全に緑色になった.この体色の変わるアブラムシの体液を赤色系統のアブラムシに導入したところ,体色が緑に変化する個体が現れた(未発表データ).このことは,アブラムシの体液に存在するなんらかの因子が体色を変えていることを示唆していた.そこで,体液に存在する任意共生細菌が体色変化の要因である可能性を検証した.
2.体色の変化するアブラムシから検出された新規の共生細菌
体色の変化するアブラムシ系統の体内に生息する共生細菌群集を細菌16S rRNA遺伝子塩基配列にもとづいて解析したところ,生存に必須な共生細菌Buchnera以外に2種類の共生細菌が検出された.ひとつは既知の共生細菌HamiltonellaもしくはSerratiaで,アブラムシ系統によっていずれかが感染していた.さらに,これらの系統すべてにこれまでアブラムシからは報告のなかったRickettsiella属の共生細菌が共感染していた.Rickettsiellaはほかの昆虫類では病原細菌として知られ,進化的にヒトのQ熱病原体(Coxiella)やレジオネラ症病原体(Legionella)と近縁である.
3.体色を変える共生細菌
共存する共生細菌のうちいずれが体色の変化にかかわっているのかを調べるため,抗生物質を処理して共生細菌の除去を試みた(図1a).その結果,HamiltonellaおよびSerratiaを除去することに成功したが,これらの共生細菌を除去しても成虫の体色は緑色のままであった.つぎに,Rickettsiellaに感染しているアブラムシ系統の体液を採取して非感染系統に微小注入しその子孫を個別飼育することで,遺伝的な背景は同一でありながらRickettsiella感染あるいは非感染だけが異なる多数のアブラムシ系統を作製した(図1b).すると,Rickettsiellaの感染した赤色系統ではすべて体色が緑色に変化した(図1b).もともと緑色だった系統ではRickettsiellaを感染させても特段の変化はみられなかった.定量PCRを用いた解析の結果は,アブラムシ体内のRickettsiellaの感染密度が高くなるほど体色が緑色に変化するという有意な関係を示した.これらの結果は,Rickettsiellaがエンドウヒゲナガアブラムシの野外集団における緑色個体の形成にかかわっていることを示していた.
4.Rickettsiellaの適応度形質にあたえる影響,体内局在,野外における感染実態
さきに述べたような方法で3系統のアブラムシについてRickettsiella感染個体と非感染個体とを作製し,体重,成長速度,産子数,寿命を比較した.その結果,感染による有意な違いはほとんど検出されなかった.
蛍光in situハイブリダイゼーションや電子顕微鏡による観察の結果,Rickettsiellaはアブラムシのほかの任意共生細菌と同様に,アブラムシ体内の二次菌細胞や鞘細胞,体液中に存在していることが判明した.
ヨーロッパ野外集団に由来するエンドウヒゲナガアブラムシ353個体につき診断PCR法を用いて調べたところ,28個体(7.9%)がRickettsiellaに感染しており,自然界における広範な分布が判明した.
自然界における体色とRickettsiella感染との関係を調べると,緑色の個体がすべてRickettsiellaに感染しているわけではなく,また,Rickettsiellaに感染していても赤色のままの個体も存在していた.このことは,体色はアブラムシとRickettsiellaとの相互作用によって決まることを意味していた.
5.Rickettsiellaがアブラムシの色素生産にあたえる影響
アブラムシの体色はおもに,黄色~赤色のカロテノイド系色素と,緑色~青色などさまざまな色のaphin(または,aphinin)と総称される多環式キノン系色素から構成されている12).Rickettsiellaに感染したアブラムシ(緑色)と非感染のアブラムシ(赤色)の色素分析を行ったところ,カロテノイド系色素の組成や量には大きな違いはみられなかった.また,さきに述べたように,アブラムシの赤色カロテノイドの合成をつかさどるカロテノイド不飽和化酵素の遺伝子発現を定量RT-PCR法により解析した結果からも,Rickettsiella感染が赤色カロテノイドの合成に変化をもたらしているという証拠は得られなかった.一方,多環式キノン配糖体であると推定される緑色色素を逆相薄相クロマトグラフィーにより解析した結果,新規な成分の出現や組成の変化はないものの,その量が感染アブラムシでは非感染アブラムシの3倍以上に増加していた.すなわち,Rickettsiellaが感染することで宿主のアブラムシの緑色色素の生産がなんらかのかたちで活性化され,その結果,体色の変化が生じているものと推察された.
6.Rickettsiella感染と捕食-被食関係への影響
はじめに述べたように,アブラムシの体色は捕食あるいは寄生の回避に重要な役割をはたしている.ゆえに,Rickettsiellaに感染して体色が赤色から緑色に変化すると,テントウムシには食べられにくくなる一方,寄生バチの攻撃はうけやすくなることが予想された1,2)(図2a).興味深いことに,Rickettsiellaに感染しているアブラムシの大部分(約80%)はHamiltonellaもしくはSerratiaという共生細菌にも感染していた(図2b).HamiltonellaやSerratiaは産みつけられた卵や幼虫を殺すことにより寄生バチに対する耐性を賦与する共生細菌として知られているのはさきに述べたとおりである9).このことは,Rickettsiellaはアブラムシの体色を緑色に変えてテントウムシに補食されにくくすると同時に,(緑色のアブラムシに好んで産卵する)寄生バチへの耐性をあたえる共生細菌と共感染することにより宿主アブラムシの生存率,ひいては,自らの生存率を上げている可能性を示唆していた.
おわりに
この研究では,自然界においてさまざまな役割をはたしている“生物の体の色”に体内の共生微生物が大きく影響している場合のあることを世界ではじめて実証した.この研究成果は生物の生態や環境適応の理解に新たな観点を提示するものといえよう.
現在,Rickettsiellaの感染によって生産が活性化される緑色色素の化学構造の詳細な分析とともに,体色変化をひき起こすRickettsiellaの全ゲノム配列の解読,さらには,感染によって活性化(あるいは,抑制)される宿主アブラムシの遺伝子の網羅的な発現解析などに取り組んでいる.この興味深い生命現象を題材にした研究の進展により,生命原理に関する新たな知見の得られることがおおいに期待される.また,これらの解析で明らかになった色素生産の活性化の機構を応用することで,将来的には,色素の効率的な生産方法や生物の色彩や外観を制御するための新規技術の開発につながる可能性もある.
文 献
- Losey, J. E., Harmon, J., Ballantyne, F. et al.: A polymorphism maintained by opposite patterns of parasitism and predation. Nature, 388, 269-272 (1997)
- Libbrecht, R., Gwynn, D. M. & Fellowes, M. D. E.: Aphidius ervi preferentially attacks the green morph of the pea aphid, Acyrthosiphon pisum. J. Insect Behav., 20, 25-32 (2007)
- Muller, F. P.: Z. Pflanzenkr., 69, 129 (1962)
- The International Aphid Genomics Consortium: Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol., 8, e1000313 (2010)[PubMed]
- Moran, N. A. & Jarvik, T.: Lateral transfer of genes from fungi underlies carotenoid production in aphids. Science, 328, 624-627 (2010)[PubMed]
- Baumann, P.: Biology bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu Rev Microbiol., 59, 155-189 (2005)[PubMed]
- Oliver, K. M., Degnan, P. H., Burke, G. R. et al.: Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol., 55, 247-266 (2010)[PubMed]
- Montllor, C. B., Maxmen, A. & Purcell, A. H.: Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol. Entomol., 27, 189-195 (2002)
- Oliver, K. M., Russell, J. A., Moran, N. A. et al.: Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA, 100, 1803-1807 (2003)[PubMed]
- Tsuchida, T., Koga, R. & Fukatsu, T.: Host plant specialization governed by facultative symbiont. Science, 303, 1989 (2004)[PubMed]
- Scarborough, C. L., Ferrari, J. & Godfray H. C. J.: Aphid protected from pathogen by endosymbiont. Science, 310, 1781 (2005)[PubMed]
- Brown, K. S.: The chemistry of aphids and scale insects. Chem. Soc. Rev., 4, 263-288 (1975)
著者プロフィール
略歴:2003年 東京大学大学院総合文化研究科 修了,同年 英国York大学 博士研究員,2004年 産業技術総合研究所生物機能工学研究部門 博士研究員を経て,2008年より理化学研究所基幹研究所 基礎科学特別研究員.
研究テーマ:昆虫と共生微生物の生物間相互作用を成立させる生理・分子機構の解明,共生微生物が生態系ではたす役割の解明.
抱負:さまざまな手法を有機的に組み合わせた解析により,生物の体内に住む小さな共生微生物の大きな役割を包括的に明らかにすることをめざしています.
© 2010 土田 努 Licensed under CC 表示 2.1 日本
