完全長ZP3のX線結晶構造からみえてきた卵被膜の組み立てと卵-精子相互作用の分子機構
奥村 裕紀
(名城大学農学部 応用生物化学科生物化学研究室)
email:奥村裕紀
DOI: 10.7875/first.author.2010.053
Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3.
Ling Han, Magnus Monné, Hiroki Okumura, Thomas Schwend, Amy L. Cherry, David Flot, Tsukasa Matsuda, Luca Jovine
Cell, 143, 404-415 (2010)
ZP3は哺乳類の卵細胞をおおう透明帯の主要な構成タンパク質であり,受精において精子が透明帯と相互作用する際に中心的な役割をはたしている.筆者らは,ZP3のニワトリホモログは糖鎖修飾の程度のきわめて低いことに着目し,組換えニワトリZP3の結晶化およびX線結晶構造解析を行った.その結果,完全長ZP3の立体構造を2.0Åの分解能で解明することに世界ではじめて成功した.ZP3の興味深い分子構造から,その分泌や卵被膜への組み込みの分子機構が明らかとなった.驚くべきことに,これまでの研究で精子との相互作用に関与していると示唆されてきたサイト1とサイト2およびC末端付近の領域はZP3の1次構造では離れて存在するが,解明された立体構造ではタンパク質表面の狭い範囲に集中していた.この領域がZP3の精子結合面であるものと考えられた.ニワトリZP3のサイト1にO-結合型糖鎖の存在したことから,この糖鎖が付加されないよう変異を導入した組換えニワトリZP3を調製し精子との結合実験を行った.その結果,サイト1のO-結合型糖鎖が精子との結合に重要である一方,ペプチド部分にも結合領域の存在する可能性が示された.
脊椎動物の卵細胞はある特殊な細胞外マトリックスにおおわれた状態で卵巣から排卵され,精子と出会い,受精する.その細胞外マトリックスは哺乳類では透明帯(zona pellucida:ZP),そのほかの脊椎動物では卵被膜(vitelline envelope:VE)などと命名されている.受精初期の卵-精子相互作用における透明帯の主要な役割は,精子が種特異的に卵と結合するための足場となり,精子先体反応(精子頭部の先体胞からその内容物が精子外へと分泌される反応で,精子が透明帯を貫通し卵の細胞膜へ達するために重要であると考えられる)を誘導することである1).また,透明帯は最初の精子が受容されたのちの多精子受精の防止機構にも関与している1).透明帯は,約260アミノ酸残基からなり高度に保存されたCys残基をもつZPドメイン(この論文では,ZPモジュールとよぶ)をもつ4種類の糖タンパク質ZP1,ZP2,ZP3,ZP4のうち,3~4種類が繊維状に重合した構造をしているものと考えられている2).
卵-精子相互作用においてZP1~ZP4はそれぞれが異なる機能をもっているものと考えられるが,とくに1980年,BleilとWassarmanが,マウスを対象とした生化学的な研究で示して以来1),哺乳類をはじめとする脊椎動物のZP3ホモログは精子が透明帯に最初に結合する際に中心的な役割をはたしているという研究結果が数多く報告されている3,4).また,1985年,FlormanとWassarmanは,マウスZP3ではO-結合型糖鎖が精子の結合に必須であることを示した1).近年になっても,精子結合におけるZP3とその糖鎖の重要性を支持する研究結果が発表されつづけている5).その一方で,ノックアウトマウスやトランスジェニックマウスを用いた一連の研究からは,卵-精子相互作用におけるZP3とそのO-結合型糖鎖の重要性に疑問を投げかけるような結果も得られている6).
筆者らは,このように実験系によって異なる結果が得られるような生命現象には,われわれの予想できなかった複雑あるいは巧妙な分子機構が関与しているのだろうと考えた.その実態を理解し,一見,矛盾する研究結果に共通の解釈をあたえるためには,ZP3の立体構造を解明し,その構造情報にもとづいて実験系を構築し,より直接的にその作用機構を解明する必要のあるものと考えた.
1980年に,ZP3の受精への関与が示されて以来,その詳細な立体構造を解明することは,生殖科学や,とくに,卵-精子相互作用における透明帯と精子との結合機構の研究にとって大きな目標のひとつであった.タンパク質の立体構造を解明するためのもっとも強力な手法のひとつはX線結晶構造解析である.ところが,哺乳類のZP3は高度に糖鎖修飾されており(マウスでは,分子重量の約50%),濃縮したり酵素的に糖鎖を除去したりすると凝集してしまう傾向があったためその構造解析は困難をきわめた.そこで,注目したのがニワトリZP3(cZP3)である.ニワトリZP3は哺乳類のZP3との相同性の高い(ヒトZP3との相同性は約53%)一方で,N-結合型糖鎖の付加配列を1か所しかもたず7),また,鳥類のZP3はチャイニーズハムスター卵巣に由来するCHO細胞に発現させると天然型と同一の修飾をうけ高濃度で培地に分泌されることが知られていた8).
ニワトリZP3の完全長cDNAをもとに,これまでの研究で蓄積されてきたマウスZP3における知見,2次構造予測や発現させた組換えニワトリZP3の限定トリプシン消化による溶解性の変化の結果などをふまえて,いくつかの変異を段階的に導入した発現コンストラクトを作製した.導入した変異は,1)C末端膜貫通領域の6Hisタグへの置換,2)Asn159のGlnへの置換によるN-結合型糖鎖の付加配列の不活性化,3)Arg359,Arg361,Arg362のAlaへの置換によるフューリン切断配列の不活性化,および,4)種間で多様性のみられるN末端領域の欠失,である.これらのコンストラクトでCHO細胞を形質転換して安定発現株を樹立し,培養して培地に分泌された組換えニワトリZP3をニッケルアフィニティークロマトグラフィーとゲルろ過クロマトグラフィーによって精製した.この変異1)~3)および1)~4)をそれぞれ導入した組換えタンパク質cZP3-3とcZP3-4(図1a)を限定トリプシン消化することによってcZP3-3TおよびcZP3-4Tを調製したところ,これらを結晶化することに成功した.cZP3-4TのX線結晶構造解析の結果,ニワトリZP3の3次元構造が分解能2.0Å(PDB ID:3NK4)と分解能2.6Å(PDB ID:3NK3)で解明された.分解能2.6Åの結晶構造から,のちに述べるO-結合型糖鎖が検出された.
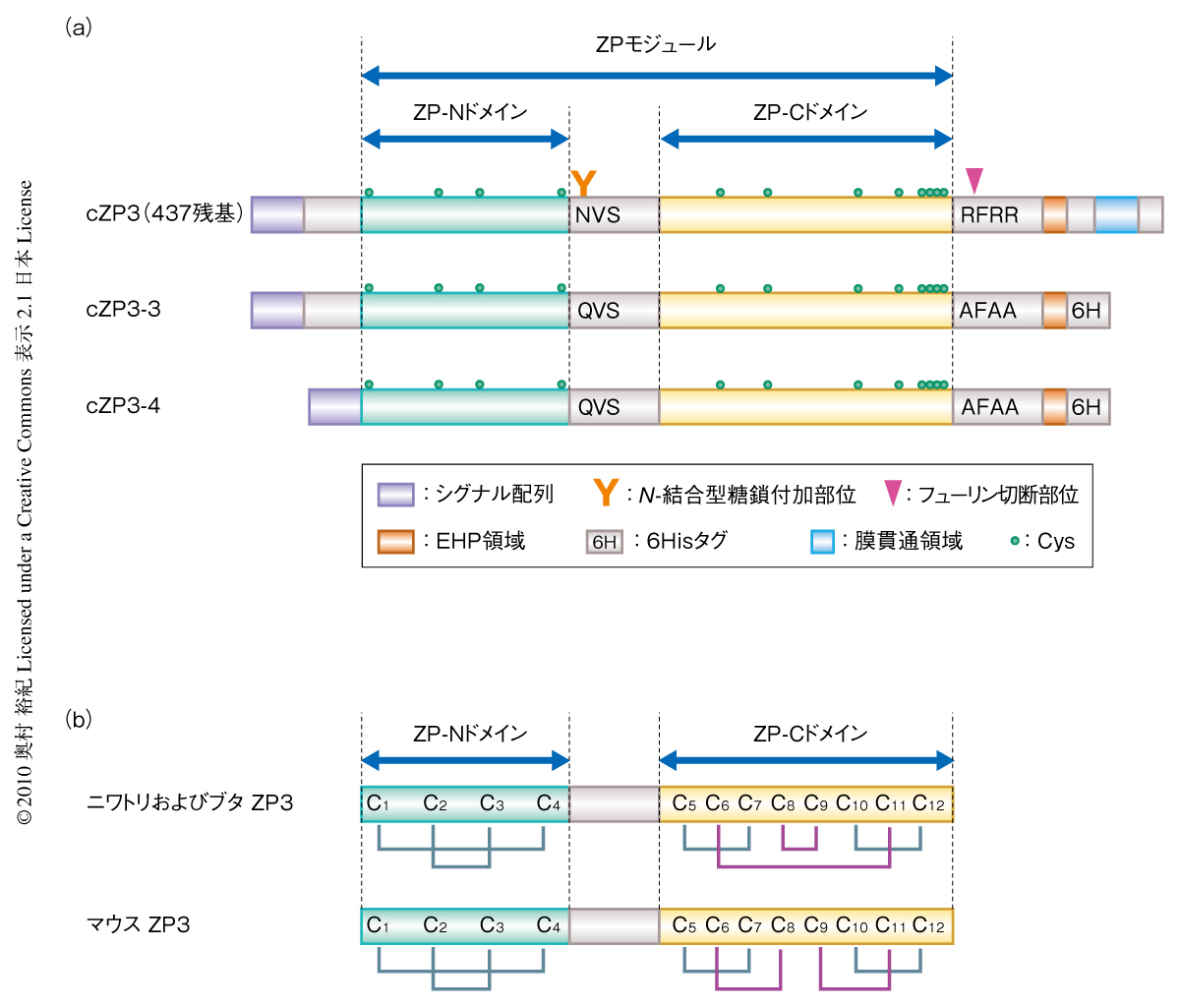
この論文ではZPドメイン(ZPモジュール)がZP-NドメインとZP-Cドメインの2つに分割できることが明確になったが,解明された結晶構造ではZP-NドメインとZP-Cドメインとを結ぶリンカー領域の一部とcZP3-4の限定トリプシン消化によって除去された領域(Arg348~Arg358,質量分析により同定)の前後が不明瞭であったものの,ニワトリZP3はそれぞれの単量体のあいだでZP-NドメインとZP-Cドメインとが非共有結合で会合したホモ2量体を形成していることが明らかとなった(図2).培地に分泌されたcZP3-4の大部分はホモ2量体を形成していることがゲルろ過クロマトグラフィーや架橋形成実験によって確認され,さらに,解明された構造情報にもとづいて2量体形成に関与していると考えられるアミノ酸残基に変異を導入したところ,その変異体は培地へ分泌されなくなった.よって,ニワトリZP3が卵細胞の周囲へ正常に分泌されるためにはホモ2量体を形成する必要のあることが示唆された.
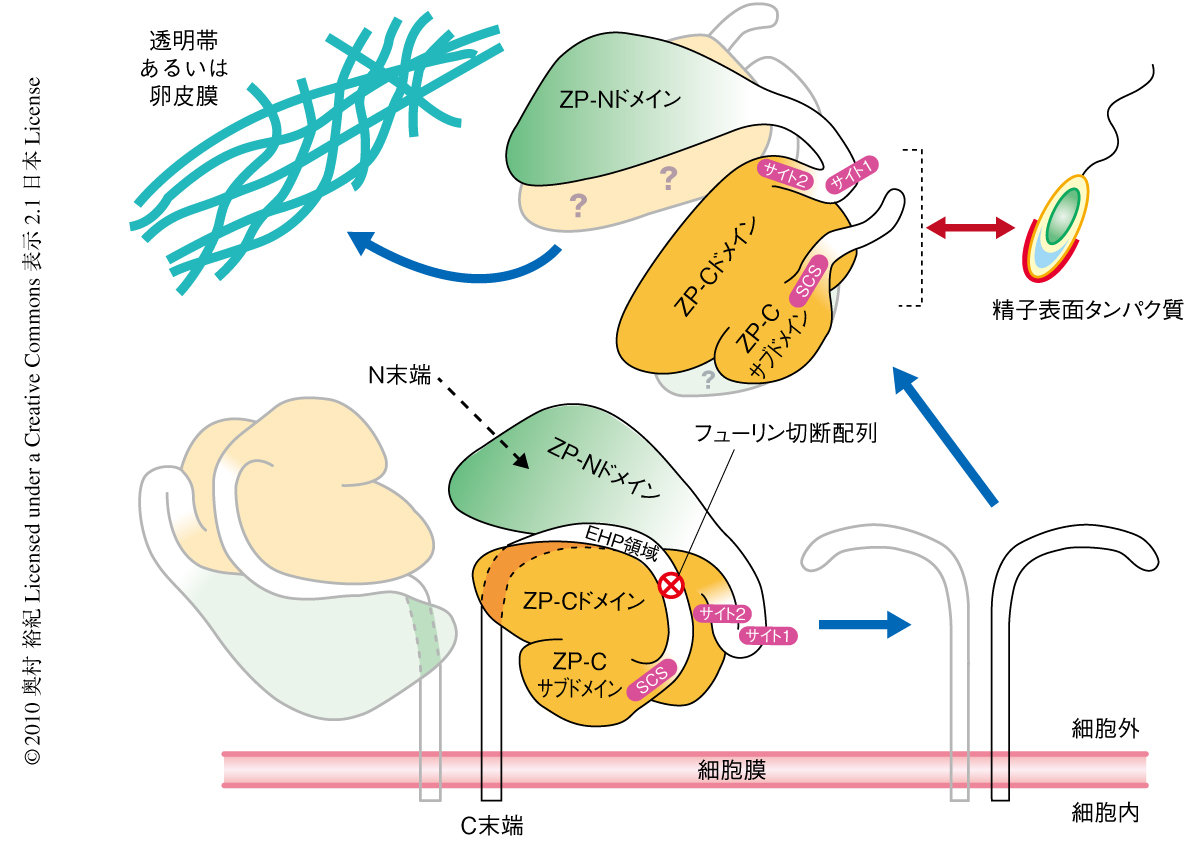
驚くべきことに,cZP3-4Tでは不活性化されたフューリン切断配列の直前に位置するArg348~Arg358が除去されているにもかかわらず,そのC末端側に位置するプロペプチドは組換えニワトリZP3の精製の際も遊離することなくZP-Cドメインの一部として存在していた.このプロペプチドはこれまでの研究でEHP(external hydrophobic patch)領域と命名された疎水性領域を含んでおり,疎水結合によってZP-NドメインとZP-Cドメインとを強固に結びつけていることが(図2),構造情報とそれにもとづくアミノ酸残基の置換実験から示唆された.さらに,cZP3-4Tをニワトリの平均体温である39℃で30時間インキュベーションしたところその約40%からプロペプチドの遊離していることが確認された.プロペプチドが除去されたタンパク質は天然型ニワトリZP3と同様に重合度が異なる何種類かのホモ多量体を形成していたのに対して,4℃で同様にインキュベーションしたあとで精製したプロペプチドをもたないタンパク質と,39℃で同様にインキュベーションしたトリプシン消化していないcZP3-4は,ほとんどが2量体を形成したままであった.
以上より,ZP3の分泌と卵被膜への組み込みについてつぎのような分子機構の存在が示唆された(図2).まず,ZP3はホモ2量体を形成して細胞外へ分泌される.分泌の過程あるいはその直後に単量体のZP-NドメインとZP-Cドメインとを強固に結びつけていたプロペプチド(および,そのC末端側につづく膜貫通領域)がフューリンによる切断をうけて時間をかけて自発的に遊離する.すると,ZP-NドメインとZP-Cドメインに存在する疎水性領域が露出し,ZP3はこれらの領域を介してZP3あるいはZPモジュール(ZPドメイン)をもつほかの卵被膜構成タンパク質と結合することにより透明帯(卵被膜)へ組み込まれるのである.
ZP3のZPモジュールには12か所の保存されたCys残基(C1~C12)が存在する.解明されたニワトリZP3の構造ではC1~C4はZP-Nドメインに存在しC1-C4およびC2-C3の組合せでS-S結合が形成されていた.また,ZP-Cドメインに存在するC5,C7,C10,C12についてはC5-C7およびC10-C12の組合せでS-S結合が形成されていた.これらの結果(図1b)は,これまでほかの脊椎動物のZP3について質量分析を利用して導き出された結果を確認するものであった9).ところが,残るC6,C8,C9,C11のあいだではC6-C11およびC8-C9の組合せでS-S結合が形成されており,ブタのZP3の質量分析によって示された結果とは一致していたものの,マウスのZP3などで同様に示された結果(C6-C8およびC9-C11)とは異なっていた(図1b).
ニワトリではZP3が卵被膜繊維の形成に重要であり10),また,ZP3と同様にS-S結合の組合せがマウスなどとは異なると考えられるニワトリのZP1では9),ZP-CドメインがZP3との複合体形成に関与していることが示唆されている11).S-S結合の多様性はZP-Cドメインの立体構造に多様性のあることを示唆しており,卵被膜構成タンパク質が卵被膜へ組み込まれる際の種特異性に関与しているものと考えられた.
なお,これまでの研究では,ZP3においてC9~C12を含む領域はZP-Cドメインとは独立したドメインを形成していると考えられてきた.しかし,今回,解明されたZP3の立体構造から,C6を含む領域とC8,C9~C12を含む領域とが近接してサブドメインを形成していることが明らかとなった.したがって,ZP3においてはC5~C12を含む領域をZP-Cドメインとし,C6,C8およびC9~C12を含むZP-Cサブドメインが形成されていると考えるのが適当だと思われた.
ニワトリZP3のO-結合型糖鎖はThr168に付加されたGalβ1-3GalNAc(T抗原)のみであることが明らかとなった.このThr残基は魚類,両生類,鳥類,哺乳類のZP3で高度に保存されておりサイト1とよばれている(図2).質量分析によってマウスZP3のサイト1(Thr155)にもO-結合型糖鎖の結合していることが確認されている12).マウスZP3ではサイト1のほかにサイト2とよばれるSer/Thr残基に富む領域(Arg160-Ser165,ニワトリZP3ではAsn173-Ala178に対応,図2)もO-結合型糖鎖をもっていることが示されている12).さきに述べたように,マウスZP3ではO-結合型糖鎖が精子の結合に必須であるため1),サイト1とサイト2は精子との結合に関与しているものと考えられている.また,精子結合領域とよばれる領域(Ile319-His339,ニワトリZP3ではVal331-Pro351に対応,図2)はO-結合型糖鎖をもっていないが12),精子との結合に重要であることが示されている1).驚くべきことに,サイト1,サイト2および精子結合領域は1次構造では離れている(ZP-NドメインとZP-Cドメインとのあいだのリンカー領域,および,フューリン切断配列の直前)が立体構造では分子表面の狭い領域に集合していることが明らかとなり(図2),この領域がニワトリZP3における精子との結合面であると考えられた.
解明された立体構造でサイト1に存在することの明らかとなった糖鎖の精子との結合への関与を確認するため,cZP3-4のサイト1をAlaに置換することによって糖鎖が付加されないcZP3-4T168Aを調製し,ニワトリ精子頭部先端への結合能をcZP3-4と比較した.その結果,ニワトリZP3が結合した精子の割合は約80%も減少し,サイト1に付加されたO-結合型糖鎖の精子結合における重要性が証明された.しかし,このcZP3-4T168Aも一定の割合で精子に結合したことから,ニワトリZP3のペプチド部分も少なからず精子との結合に関与しているものと考えられた.
以上より,ZP3の精子との結合には,サイト1,サイト2,精子結合領域のすべてが協調して関与しているものと考えられた.また,精子結合領域のC末端側半分からフューリン切断配列までの領域はcZP3-4Tの結晶構造では検出されなかったが,構造決定された部分との位置関係からサイト1,サイト2,精子結合領域と同じ面に存在することは確実であった.この領域の1次構造は種間での多様性が大きく,精子のZP3との結合の種特異性を調節し,また,精子結合領域とともに上記のペプチドを介した結合に関与しているものと考えられた.
1980年にZP3が同定されてから約30年間,その卵-精子相互作用への関与と精子との結合の分子機構について立体構造からのアプローチが強く望まれてきた.この研究では,ニワトリZP3を用いることによって困難をきわめたZP3完全長のX線結晶構造解析にはじめて成功し,その構造情報にもとづいて生理機能の解析を行うことが可能であることを示した.以上のように,ZP3の分泌機構と透明帯や卵被膜への組み込み機構,さらに,ZP3における精子との結合領域が明らかにされ,卵-精子相互作用の分子機構の解明にむけて大きな道が開かれた.
この研究で得られた成果は,ZP3の精子との結合面を標的とする低分子化合物の開発などホルモンによらず安全性のより高い避妊薬をデザインするのに利用することができるものと考えられる.そのほかにも,ZP3と精子との結合能を向上させることによる不妊症の治療,有害鳥獣の個体数の管理や希少動物の繁殖率の改善などに応用することができると期待される.また,ZPモジュール(ZPドメイン)をもつタンパク質は透明帯や卵被膜以外からも同定されているため1),ZP3の立体構造はそれらの構造解析や機能解析に対しても有用な情報を提供するであろう.
略歴:2006年 名古屋大学大学院生命農学研究科博士課程 修了,同 21世紀COEプログラム 研究員,スウェーデンKarolinska研究所 博士研究員を経て,2008年より名城大学農学部 助教.
研究テーマ:脊椎動物(とくに,鳥類)の卵-精子相互作用と卵被膜の形成の分子機構の解明.
抱負:高い好奇心と忍耐力を生かして幅広く研究・教育に励むとともに,卵-精子相互作用の謎の解明に取り組みつづける.
© 2010 奥村 裕紀 Licensed under CC 表示 2.1 日本
(名城大学農学部 応用生物化学科生物化学研究室)
email:奥村裕紀
DOI: 10.7875/first.author.2010.053
Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3.
Ling Han, Magnus Monné, Hiroki Okumura, Thomas Schwend, Amy L. Cherry, David Flot, Tsukasa Matsuda, Luca Jovine
Cell, 143, 404-415 (2010)
要 約
ZP3は哺乳類の卵細胞をおおう透明帯の主要な構成タンパク質であり,受精において精子が透明帯と相互作用する際に中心的な役割をはたしている.筆者らは,ZP3のニワトリホモログは糖鎖修飾の程度のきわめて低いことに着目し,組換えニワトリZP3の結晶化およびX線結晶構造解析を行った.その結果,完全長ZP3の立体構造を2.0Åの分解能で解明することに世界ではじめて成功した.ZP3の興味深い分子構造から,その分泌や卵被膜への組み込みの分子機構が明らかとなった.驚くべきことに,これまでの研究で精子との相互作用に関与していると示唆されてきたサイト1とサイト2およびC末端付近の領域はZP3の1次構造では離れて存在するが,解明された立体構造ではタンパク質表面の狭い範囲に集中していた.この領域がZP3の精子結合面であるものと考えられた.ニワトリZP3のサイト1にO-結合型糖鎖の存在したことから,この糖鎖が付加されないよう変異を導入した組換えニワトリZP3を調製し精子との結合実験を行った.その結果,サイト1のO-結合型糖鎖が精子との結合に重要である一方,ペプチド部分にも結合領域の存在する可能性が示された.
はじめに
脊椎動物の卵細胞はある特殊な細胞外マトリックスにおおわれた状態で卵巣から排卵され,精子と出会い,受精する.その細胞外マトリックスは哺乳類では透明帯(zona pellucida:ZP),そのほかの脊椎動物では卵被膜(vitelline envelope:VE)などと命名されている.受精初期の卵-精子相互作用における透明帯の主要な役割は,精子が種特異的に卵と結合するための足場となり,精子先体反応(精子頭部の先体胞からその内容物が精子外へと分泌される反応で,精子が透明帯を貫通し卵の細胞膜へ達するために重要であると考えられる)を誘導することである1).また,透明帯は最初の精子が受容されたのちの多精子受精の防止機構にも関与している1).透明帯は,約260アミノ酸残基からなり高度に保存されたCys残基をもつZPドメイン(この論文では,ZPモジュールとよぶ)をもつ4種類の糖タンパク質ZP1,ZP2,ZP3,ZP4のうち,3~4種類が繊維状に重合した構造をしているものと考えられている2).
卵-精子相互作用においてZP1~ZP4はそれぞれが異なる機能をもっているものと考えられるが,とくに1980年,BleilとWassarmanが,マウスを対象とした生化学的な研究で示して以来1),哺乳類をはじめとする脊椎動物のZP3ホモログは精子が透明帯に最初に結合する際に中心的な役割をはたしているという研究結果が数多く報告されている3,4).また,1985年,FlormanとWassarmanは,マウスZP3ではO-結合型糖鎖が精子の結合に必須であることを示した1).近年になっても,精子結合におけるZP3とその糖鎖の重要性を支持する研究結果が発表されつづけている5).その一方で,ノックアウトマウスやトランスジェニックマウスを用いた一連の研究からは,卵-精子相互作用におけるZP3とそのO-結合型糖鎖の重要性に疑問を投げかけるような結果も得られている6).
筆者らは,このように実験系によって異なる結果が得られるような生命現象には,われわれの予想できなかった複雑あるいは巧妙な分子機構が関与しているのだろうと考えた.その実態を理解し,一見,矛盾する研究結果に共通の解釈をあたえるためには,ZP3の立体構造を解明し,その構造情報にもとづいて実験系を構築し,より直接的にその作用機構を解明する必要のあるものと考えた.
1.組換えニワトリZP3の調製とX線結晶構造の解明
1980年に,ZP3の受精への関与が示されて以来,その詳細な立体構造を解明することは,生殖科学や,とくに,卵-精子相互作用における透明帯と精子との結合機構の研究にとって大きな目標のひとつであった.タンパク質の立体構造を解明するためのもっとも強力な手法のひとつはX線結晶構造解析である.ところが,哺乳類のZP3は高度に糖鎖修飾されており(マウスでは,分子重量の約50%),濃縮したり酵素的に糖鎖を除去したりすると凝集してしまう傾向があったためその構造解析は困難をきわめた.そこで,注目したのがニワトリZP3(cZP3)である.ニワトリZP3は哺乳類のZP3との相同性の高い(ヒトZP3との相同性は約53%)一方で,N-結合型糖鎖の付加配列を1か所しかもたず7),また,鳥類のZP3はチャイニーズハムスター卵巣に由来するCHO細胞に発現させると天然型と同一の修飾をうけ高濃度で培地に分泌されることが知られていた8).
ニワトリZP3の完全長cDNAをもとに,これまでの研究で蓄積されてきたマウスZP3における知見,2次構造予測や発現させた組換えニワトリZP3の限定トリプシン消化による溶解性の変化の結果などをふまえて,いくつかの変異を段階的に導入した発現コンストラクトを作製した.導入した変異は,1)C末端膜貫通領域の6Hisタグへの置換,2)Asn159のGlnへの置換によるN-結合型糖鎖の付加配列の不活性化,3)Arg359,Arg361,Arg362のAlaへの置換によるフューリン切断配列の不活性化,および,4)種間で多様性のみられるN末端領域の欠失,である.これらのコンストラクトでCHO細胞を形質転換して安定発現株を樹立し,培養して培地に分泌された組換えニワトリZP3をニッケルアフィニティークロマトグラフィーとゲルろ過クロマトグラフィーによって精製した.この変異1)~3)および1)~4)をそれぞれ導入した組換えタンパク質cZP3-3とcZP3-4(図1a)を限定トリプシン消化することによってcZP3-3TおよびcZP3-4Tを調製したところ,これらを結晶化することに成功した.cZP3-4TのX線結晶構造解析の結果,ニワトリZP3の3次元構造が分解能2.0Å(PDB ID:3NK4)と分解能2.6Å(PDB ID:3NK3)で解明された.分解能2.6Åの結晶構造から,のちに述べるO-結合型糖鎖が検出された.
2.分泌機構および卵被膜への組み込み機構との関連
この論文ではZPドメイン(ZPモジュール)がZP-NドメインとZP-Cドメインの2つに分割できることが明確になったが,解明された結晶構造ではZP-NドメインとZP-Cドメインとを結ぶリンカー領域の一部とcZP3-4の限定トリプシン消化によって除去された領域(Arg348~Arg358,質量分析により同定)の前後が不明瞭であったものの,ニワトリZP3はそれぞれの単量体のあいだでZP-NドメインとZP-Cドメインとが非共有結合で会合したホモ2量体を形成していることが明らかとなった(図2).培地に分泌されたcZP3-4の大部分はホモ2量体を形成していることがゲルろ過クロマトグラフィーや架橋形成実験によって確認され,さらに,解明された構造情報にもとづいて2量体形成に関与していると考えられるアミノ酸残基に変異を導入したところ,その変異体は培地へ分泌されなくなった.よって,ニワトリZP3が卵細胞の周囲へ正常に分泌されるためにはホモ2量体を形成する必要のあることが示唆された.
驚くべきことに,cZP3-4Tでは不活性化されたフューリン切断配列の直前に位置するArg348~Arg358が除去されているにもかかわらず,そのC末端側に位置するプロペプチドは組換えニワトリZP3の精製の際も遊離することなくZP-Cドメインの一部として存在していた.このプロペプチドはこれまでの研究でEHP(external hydrophobic patch)領域と命名された疎水性領域を含んでおり,疎水結合によってZP-NドメインとZP-Cドメインとを強固に結びつけていることが(図2),構造情報とそれにもとづくアミノ酸残基の置換実験から示唆された.さらに,cZP3-4Tをニワトリの平均体温である39℃で30時間インキュベーションしたところその約40%からプロペプチドの遊離していることが確認された.プロペプチドが除去されたタンパク質は天然型ニワトリZP3と同様に重合度が異なる何種類かのホモ多量体を形成していたのに対して,4℃で同様にインキュベーションしたあとで精製したプロペプチドをもたないタンパク質と,39℃で同様にインキュベーションしたトリプシン消化していないcZP3-4は,ほとんどが2量体を形成したままであった.
以上より,ZP3の分泌と卵被膜への組み込みについてつぎのような分子機構の存在が示唆された(図2).まず,ZP3はホモ2量体を形成して細胞外へ分泌される.分泌の過程あるいはその直後に単量体のZP-NドメインとZP-Cドメインとを強固に結びつけていたプロペプチド(および,そのC末端側につづく膜貫通領域)がフューリンによる切断をうけて時間をかけて自発的に遊離する.すると,ZP-NドメインとZP-Cドメインに存在する疎水性領域が露出し,ZP3はこれらの領域を介してZP3あるいはZPモジュール(ZPドメイン)をもつほかの卵被膜構成タンパク質と結合することにより透明帯(卵被膜)へ組み込まれるのである.
3.分子内S-S結合の多様性とZPモジュールの構造
ZP3のZPモジュールには12か所の保存されたCys残基(C1~C12)が存在する.解明されたニワトリZP3の構造ではC1~C4はZP-Nドメインに存在しC1-C4およびC2-C3の組合せでS-S結合が形成されていた.また,ZP-Cドメインに存在するC5,C7,C10,C12についてはC5-C7およびC10-C12の組合せでS-S結合が形成されていた.これらの結果(図1b)は,これまでほかの脊椎動物のZP3について質量分析を利用して導き出された結果を確認するものであった9).ところが,残るC6,C8,C9,C11のあいだではC6-C11およびC8-C9の組合せでS-S結合が形成されており,ブタのZP3の質量分析によって示された結果とは一致していたものの,マウスのZP3などで同様に示された結果(C6-C8およびC9-C11)とは異なっていた(図1b).
ニワトリではZP3が卵被膜繊維の形成に重要であり10),また,ZP3と同様にS-S結合の組合せがマウスなどとは異なると考えられるニワトリのZP1では9),ZP-CドメインがZP3との複合体形成に関与していることが示唆されている11).S-S結合の多様性はZP-Cドメインの立体構造に多様性のあることを示唆しており,卵被膜構成タンパク質が卵被膜へ組み込まれる際の種特異性に関与しているものと考えられた.
なお,これまでの研究では,ZP3においてC9~C12を含む領域はZP-Cドメインとは独立したドメインを形成していると考えられてきた.しかし,今回,解明されたZP3の立体構造から,C6を含む領域とC8,C9~C12を含む領域とが近接してサブドメインを形成していることが明らかとなった.したがって,ZP3においてはC5~C12を含む領域をZP-Cドメインとし,C6,C8およびC9~C12を含むZP-Cサブドメインが形成されていると考えるのが適当だと思われた.
4.ZP3と精子との結合機構
ニワトリZP3のO-結合型糖鎖はThr168に付加されたGalβ1-3GalNAc(T抗原)のみであることが明らかとなった.このThr残基は魚類,両生類,鳥類,哺乳類のZP3で高度に保存されておりサイト1とよばれている(図2).質量分析によってマウスZP3のサイト1(Thr155)にもO-結合型糖鎖の結合していることが確認されている12).マウスZP3ではサイト1のほかにサイト2とよばれるSer/Thr残基に富む領域(Arg160-Ser165,ニワトリZP3ではAsn173-Ala178に対応,図2)もO-結合型糖鎖をもっていることが示されている12).さきに述べたように,マウスZP3ではO-結合型糖鎖が精子の結合に必須であるため1),サイト1とサイト2は精子との結合に関与しているものと考えられている.また,精子結合領域とよばれる領域(Ile319-His339,ニワトリZP3ではVal331-Pro351に対応,図2)はO-結合型糖鎖をもっていないが12),精子との結合に重要であることが示されている1).驚くべきことに,サイト1,サイト2および精子結合領域は1次構造では離れている(ZP-NドメインとZP-Cドメインとのあいだのリンカー領域,および,フューリン切断配列の直前)が立体構造では分子表面の狭い領域に集合していることが明らかとなり(図2),この領域がニワトリZP3における精子との結合面であると考えられた.
解明された立体構造でサイト1に存在することの明らかとなった糖鎖の精子との結合への関与を確認するため,cZP3-4のサイト1をAlaに置換することによって糖鎖が付加されないcZP3-4T168Aを調製し,ニワトリ精子頭部先端への結合能をcZP3-4と比較した.その結果,ニワトリZP3が結合した精子の割合は約80%も減少し,サイト1に付加されたO-結合型糖鎖の精子結合における重要性が証明された.しかし,このcZP3-4T168Aも一定の割合で精子に結合したことから,ニワトリZP3のペプチド部分も少なからず精子との結合に関与しているものと考えられた.
以上より,ZP3の精子との結合には,サイト1,サイト2,精子結合領域のすべてが協調して関与しているものと考えられた.また,精子結合領域のC末端側半分からフューリン切断配列までの領域はcZP3-4Tの結晶構造では検出されなかったが,構造決定された部分との位置関係からサイト1,サイト2,精子結合領域と同じ面に存在することは確実であった.この領域の1次構造は種間での多様性が大きく,精子のZP3との結合の種特異性を調節し,また,精子結合領域とともに上記のペプチドを介した結合に関与しているものと考えられた.
おわりに
1980年にZP3が同定されてから約30年間,その卵-精子相互作用への関与と精子との結合の分子機構について立体構造からのアプローチが強く望まれてきた.この研究では,ニワトリZP3を用いることによって困難をきわめたZP3完全長のX線結晶構造解析にはじめて成功し,その構造情報にもとづいて生理機能の解析を行うことが可能であることを示した.以上のように,ZP3の分泌機構と透明帯や卵被膜への組み込み機構,さらに,ZP3における精子との結合領域が明らかにされ,卵-精子相互作用の分子機構の解明にむけて大きな道が開かれた.
この研究で得られた成果は,ZP3の精子との結合面を標的とする低分子化合物の開発などホルモンによらず安全性のより高い避妊薬をデザインするのに利用することができるものと考えられる.そのほかにも,ZP3と精子との結合能を向上させることによる不妊症の治療,有害鳥獣の個体数の管理や希少動物の繁殖率の改善などに応用することができると期待される.また,ZPモジュール(ZPドメイン)をもつタンパク質は透明帯や卵被膜以外からも同定されているため1),ZP3の立体構造はそれらの構造解析や機能解析に対しても有用な情報を提供するであろう.
文 献
- Wassarman, P. M., Jovine, L. & Litscher, E. S.: A profile of fertilization in mammals. Nat. Cell Biol., 3, E59-64 (2001)[PubMed]
- Goudet, G., Mugnier, S., Callebaut, I. et al.: Phylogenetic analysis and identification of pseudogenes reveal a progressive loss of zona pellucida genes during evolution of vertebrates. Biol. Reprod., 78, 796-806 (2008)[PubMed]
- Yurewicz, E. C., Sacco, A. G., Gupta, S. K. et. al.: Hetero-oligomerization-dependent binding of pig oocyte zona pellucida glycoproteins ZPB and ZPC to boar sperm membrane vesicles. J. Biol. Chem., 273, 7488-7494 (1998)[PubMed]
- Bausek, N., Ruckenbauer, H. H., Pfeifer, S. et al.: Interaction of sperm with purified native chicken ZP1 and ZPC proteins. Biol. Reprod., 71, 684-690 (2004)[PubMed]
- Bansal, P., Chakrabarti, K. & Gupta, S. K.: Functional activity of human ZP3 primary sperm receptor resides toward its C-terminus. Biol. Reprod., 81, 7-15 (2009)[PubMed]
- Williams, S. A., Xia, L., Cummings, R. D. et al.: Fertilization in mouse does not require terminal galactose or N-acetylglucosamine on the zona pellucida glycans. J. Cell Sci., 120, 1341-1349 (2007)[PubMed]
- Takeuchi, Y., Nishimura, K., Aoki, N. et al.: A 42-kDa glycoprotein from chicken egg-envelope, an avian homolog of the ZPC family glycoproteins in mammalian zona pellucida. Its first identification, cDNA cloning and granulosa cell-specific expression. Eur. J. Biochem., 260, 736-742 (1999)[PubMed]
- Sasanami, T., Toriyama, M. & Mori, M.: Carboxy-terminal proteolytic processing at a consensus furin cleavage site is a prerequisite event for quail ZPC secretion. Biol. Reprod., 68, 1613-1619 (2003)[PubMed]
- Kanai, S., Kitayama, T., Yonezawa, N. et al.: Disulfide linkage patterns of pig zona pellucida glycoproteins ZP3 and ZP4. Mol. Reprod. Dev., 75, 847-856 (2008)[PubMed]
- Okumura, H., Okajima, T., Nadano, D. et al.: Association of chicken zona pellucida glycoprotein (ZP) B1 with ZPC induces formation of ZPB1-ZPC fibrous aggregates containing disulfide-bridged ZPB1 dimer. Biochem. Biophys. Res. Commun., 364, 682-688 (2007)[PubMed]
- Okumura, H., Aoki, N., Sato, C. et al.: Heterocomplex formation and cell-surface accumulation of hen's serum zona pellucida B1 (ZPB1) with ZPC expressed by a mammalian cell line (COS-7): a possible initiating step of egg-envelope matrix construction. Biol. Reprod., 76, 9-18 (2007)[PubMed]
- Chalabi, S., Panico, M., Sutton-Smith, M. et al.: Differential O-glycosylation of a conserved domain expressed in murine and human ZP3. Biochemistry, 45, 637-647 (2006)[PubMed]
著者プロフィール
略歴:2006年 名古屋大学大学院生命農学研究科博士課程 修了,同 21世紀COEプログラム 研究員,スウェーデンKarolinska研究所 博士研究員を経て,2008年より名城大学農学部 助教.
研究テーマ:脊椎動物(とくに,鳥類)の卵-精子相互作用と卵被膜の形成の分子機構の解明.
抱負:高い好奇心と忍耐力を生かして幅広く研究・教育に励むとともに,卵-精子相互作用の謎の解明に取り組みつづける.
© 2010 奥村 裕紀 Licensed under CC 表示 2.1 日本
