脳弓下器官において水分および塩分への欲求を制御する神経機構
松田隆志・野田昌晴
(基礎生物学研究所 統合神経生物学研究部門)
email:松田隆志,野田昌晴
DOI: 10.7875/first.author.2017.002
Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ.
Takashi Matsuda, Takeshi Y. Hiyama, Fumio Niimura, Taiji Matsusaka, Akiyoshi Fukamizu, Kenta Kobayashi, Kazuto Kobayashi, Masaharu Noda
Nature Neuroscience, 20, 230-241 (2017)
水分あるいは塩分への欲求を正しく制御するため,脳はつねに体液の状態をモニターしている.アンジオテンシンIIは水分および塩分への両方の欲求を司令するが,体液の状態に応じて,選択的にそれらのうちの一方への欲求を,あるいは,両方への欲求を亢進する神経機構は明らかにされていない.この研究において,筆者らは,光遺伝学的な手法を用いて,水分および塩分への欲求が脳弓下器官のアンジオテンシンII受容体陽性の異なる2つの興奮性ニューロンの集団により制御されることを明らかにした.水分への欲求は終板脈管器官に投射するニューロンが,一方,塩分への欲求は腹側分界条床核に投射するニューロンが担っていた.水分への欲求を制御するニューロンは,塩分の欠乏した状態においてコレシストキニンにより活性化されるGABAニューロンを介して抑制され,塩分への欲求を制御するニューロンは,水分の欠乏した状態においてNa+チャンネルのひとつであるNaxのシグナルにより別のGABAニューロンを介して抑制されていた.脳弓下器官におけるこのような水分と塩分の選択的な摂取の制御機構は,生命の維持に必要な体液の恒常性の実現に貢献すると考えられる.
ヒトを含む陸生動物はつねに水分あるいは塩分の欠乏という危険にさらされている.水分の欠乏した状態においては体液のNa+の濃度が上昇する結果,動物は渇きをおぼえるとともに塩分を避ける1,2).一方,塩分が欠乏すると塩分への欲求が高まるが,これはCl-でなくNa+の摂取によってのみ充足される3).体液の状態に応じたこれらの欲求の正しい制御は生命の維持にとり必須のしくみとなっている.
Na+は体液の主要な陽イオンであり,浸透圧を決める主要な因子である.体液におけるNa+の濃度は体液の浸透圧を生理的な範囲に維持するため脳においてつねにモニターされている.脳におけるモニターの場は血液-脳関門を欠く脳室周囲器官であると推測されていた4).筆者らは,これまでに,Na+チャンネルのひとつであるNaxが感覚性の脳室周囲器官である脳弓下器官および終板脈管器官の特殊なグリア細胞に特異的に発現しており,これがNa+の濃度の上昇を感知する脳のNa+センサーであることを明らかにしてきた2,5).Naxノックアウトマウスを用いた一連の解析から,脳弓下器官が塩分への欲求の制御のためのNa+の濃度のモニターの中枢であり,Naxの活性化はグリア細胞からの乳酸の放出の増加につながり,これがGABAニューロンの活性化を誘導することが明らかにされた6).筆者らは,このGABAニューロンが塩分の摂取の行動をつかさどるニューロンの活性を制御すると推定していた7).
アンジオテンシンIIは血管の収縮を促進するペプチドホルモンであり,血圧の制御を担うホルモンのひとつである.一方で,アンジオテンシンIIを脳室に投与すると水分および塩分の両方の摂取を促進することが知られていた8).脳室周囲器官にはアンジオテンシンII受容体(ヒトにおいてはAT1およびAT2,マウスにおいてはAT1a,AT1b,AT2)が豊富に分布しており,血液におけるアンジオテンシンIIシグナルの中枢における受容部位と考えられてきた9).
マウスを水分,塩分,あるいは,その両方の欠乏した状態にした.水分の欠乏した状態としては2日間にわたる絶水,水分および塩分の両方が欠乏した状態としては利尿剤の処理,塩分の欠乏した状態としては利尿剤の処理ののち水分のみを補給した.すべての条件において,血液中のアンジオテンシンIIは同じ程度に上昇した.野生型マウスおよびAT1aノックアウトマウスにおいて水分および塩分の摂取の行動を調べた結果,塩分の摂取は完全にAT1aに依存的であったのに対し,水分の摂取は約半分がAT1aに依存性であることが判明した.AT2ノックアウトマウスにおいてもアンジオテンシンIIに依存的に水分の摂取が減少することから10),水分の摂取の残りの半分はAT2陽性のニューロンにより担われると推測された.
塩分の欠乏した状態の脳弓下器官および終板脈管器官において,ニューロンの活性化のマーカーであるFos陽性のニューロンの数が有意に増加し,そのほとんどがAT1a陽性であった.脳室にAT1の阻害剤であるロサルタンを投与したところ,脳弓下器官あるいは終板脈管器官においてニューロンの活性化は起こらず,同時に,塩分の摂取の行動も抑制された.水分の欠乏した状態においては,塩分の欠乏した状態よりもさらにFos陽性のニューロンの数が増加したが,そこにしめるAT1a陽性のニューロンの割合は塩分の欠乏した状態よりも少なかった.また,AT1aノックアウトマウスにおいても,水分の欠乏した状態においてはFos陽性のニューロンが増加し水分の摂取も増加した.脳弓下器官あるいは終板脈管器官のいずれからAT1aを欠失させても,水分への欲求は大きく減少した.脳弓下器官において選択的にAT1aを欠失させると塩分への欲求が消失したが,終板脈管器官における選択的な欠失により塩分への欲求性はまったく影響されなかった.これらの結果から,脳弓下器官のAT1aシグナルが水分および塩分の両方の摂取に関与するのに対し,終板脈管器官のAT1aシグナルは水分の摂取だけに関与することが示された.
脳弓下器官のAT1a陽性のニューロンはVglut2陽性およびnNOS陽性であったことから,グルタミン酸作動性の興奮性ニューロンであることがわかった.その投射先を調べたところ,終板脈管器官,正中視索前核,腹側分界条床核,室傍核,視索上核と,脳の多くの部位が標的であった.逆行性の標識により調べたところ,脳弓下器官から終板脈管器官に投射するニューロンの約半分がAT1a陽性のニューロンであった.また,水分の欠乏した状態においてFos陽性になるニューロンは脳弓下器官から終板脈管器官に投射するニューロンとよく一致したことから,脳弓下器官から終板脈管器官に投射するニューロンが水分への欲求を担うであろうと推定された.光遺伝学的な手法により脳弓下器官から終板脈管器官に投射するニューロンを抑制したところ,水分への欲求が有意に減少した.対照的に,塩分への欲求はまったく影響されなかった.光遺伝学的な手法により脳弓下器官から終板脈管器官に投射するニューロンを活性化したところ,水分が足りている状態でも急速な水分の摂取の行動が誘発された.しかしながら,塩分の摂取に対しては影響がなかった.これらの結果から,脳弓下器官から終板脈管器官に投射するニューロンが特異的に水分への欲求を制御することが示された(図1).
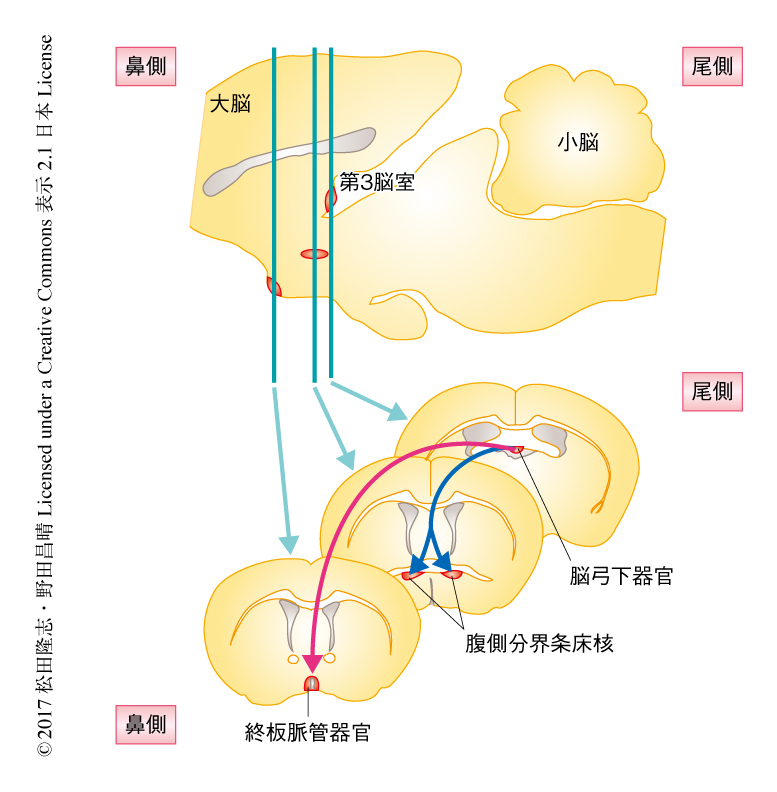
グルタミン酸作動性の脳弓下器官のニューロンの一部は腹側分界条床核に投射していたが,これらはほとんどすべてがAT1a陽性であり,塩分の欠乏したときにFos陽性になるニューロンであった.両側の腹側分界条床核を電気的に障害したところ,塩分の欠乏した状態における塩分の摂取がいちじるしく減少した.また,脳弓下器官から腹側分界条床核に投射するニューロンにおいて選択的にAT1aを欠失させたところ,欠失の程度に応じて塩分の摂取が減少した.光遺伝学的な手法により脳弓下器官から腹側分界条床核に投射するニューロンを抑制したところ,塩分の摂取が選択的に抑制された.反対に,光遺伝学的な手法により脳弓下器官から腹側分界条床核に投射するニューロンを活性化したところ,水分の欠乏した状態においても塩分の摂取が選択的に上昇した.このとき,NaClに代えてKClを用いた場合には塩分の摂取の増加は認められなかった.したがって,脳弓下器官から腹側分界条床核に投射するニューロンはNaClの摂取を選択的に制御すると結論された(図1).
Na+チャンネルのひとつであるNaxのシグナルとAT1aに依存的な塩分への欲求の制御との関係について解析した.0.3 M NaClと純水による2瓶法により測定したところ,野生型のマウスにおいては水分の欠乏した状態では体液のNa+の濃度が上昇する結果,塩分の摂取がいちじるしく減少した.すでに報告されたように,Naxノックアウトマウスにおいては水分の欠乏した状態においてもこの塩分の忌避は起こらず,0.3 M NaClの摂取の量は水と同じ程度である11)(新着論文レビュー でも掲載).対照的に,AT1aノックアウトマウスおよびNaxとAT1aのダブルノックアウトマウスにおいては,0.3 M NaClはほとんど摂取されなかった.このことから,AT1aの欠損により塩分への欲求は消失することが示された.Naxノックアウトマウスにおいては水分の欠乏した状態にてFos陽性である脳弓下器官から腹側分界条床核に投射するニューロンの数が野生型のマウスと比べ有意に多かった.このことから,脳弓下器官から腹側分界条床核に投射するニューロンの活性がNaxシグナルにより負に制御されることが示唆された(図2).

脳弓下器官から腹側分界条床核に投射するニューロンおよびその近傍のGABAニューロンの活性を電気生理学的に調べたところ,Na+の濃度が145 mMの条件において,脳弓下器官から腹側分界条床核に投射するニューロンはアンジオテンシンIIにより活性化したのに対し,GABAニューロンの活性は影響されなかった.Na+の濃度が160 mMの条件においてはGABAニューロンの活性の頻度が上昇したが,この上昇につれてアンジオテンシンIIにより誘導される脳弓下器官から腹側分界条床核に投射するニューロンの活性化は抑制された.このGABAニューロンによる脳弓下器官から腹側分界条床核に投射するニューロンの活性の制御はNaxノックアウトマウスにおいてはみられなかった.また,光遺伝学的な手法によりGABAニューロンを活性化したところ,塩分の欠乏した状態において塩分の摂取が有意に減少した.
GABAニューロンの活性化は水分の摂取も少ないながら有意に減少させたことから,脳弓下器官から終板脈管器官に投射するニューロンのGABAニューロンによる制御の可能性が推定された.脳弓下器官から終板脈管器官に投射するニューロンとそれとシナプス結合するGABAニューロンとの関係について調べたところ,Na+の濃度が145 mMの条件において脳弓下器官から終板脈管器官に投射するニューロンの活性は停止していたが,このニューロンの約半数についてはアンジオテンシンIIにより活性化することがわかった.さらに,コレシストキニンがGABAニューロンの活性化を誘導し,脳弓下器官から終板脈管器官に投射するニューロンの活性を制御することが判明した.脳弓下器官におけるコレシストキニンの濃度を調べたところ,Naの欠乏した状態でのみ約9倍の濃度の上昇がみられた.コレシストキニンは脳弓下器官から腹側分界条床核に投射するニューロンと結合するGABAニューロンの活性には影響をおよぼさなかったことから,脳弓下器官から腹側分界条床核に投射するニューロンと,脳弓下器官から終板脈管器官に投射するニューロンとをそれぞれ制御する別のGABAニューロンがあると考えられた(図2).
この研究において,筆者らが“水ニューロン”“塩ニューロン”とよぶ脳弓下器官の異なるAT1a陽性のニューロンの集団が,それぞれ水分および塩分への欲求を司令していることが明らかにされた.水ニューロンは脳弓下器官のAT1a陽性の興奮性ニューロンであり,終板脈管器官に投射していた.一方,塩ニューロンは同様のニューロンであるが,腹側分界条床核に投射していた.水ニューロンの活性はNa+の欠乏した状態にて脳弓下器官において濃度の上昇するコレシストキニンがGABAニューロンを介して制御していた.一方,塩ニューロンの活性は水の欠乏した状態においてNa+チャンネルのひとつであるNaxのシグナルが別のGABAニューロンを介して制御していた.このように,アンジオテンシンIIの2つの作用には,体液のNa+の濃度により別々の制御機構が存在していた(図2).水分の摂取については,終板脈管器官にNa+の濃度の上昇に依存するさらなる活性化の機構が存在すると推定されている12).この成果は,体液の状態により水分あるいは塩分への欲求が正しく制御される中枢機構を明らかにしたものであり,体液の制御の異常によりひき起こされるさまざまな疾患の発症機構の理解および治療のための標的の同定につながると考えられる.
略歴:総合研究大学院大学生命科学研究科博士課程 在学中.
研究テーマ:体液の恒常性の維持にかかわる神経機構.
野田 昌晴(Masaharu Noda)
基礎生物学研究所 教授.
研究室URL:http://niwww3.nibb.ac.jp/
© 2017 松田隆志・野田昌晴 Licensed under CC 表示 2.1 日本
(基礎生物学研究所 統合神経生物学研究部門)
email:松田隆志,野田昌晴
DOI: 10.7875/first.author.2017.002
Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ.
Takashi Matsuda, Takeshi Y. Hiyama, Fumio Niimura, Taiji Matsusaka, Akiyoshi Fukamizu, Kenta Kobayashi, Kazuto Kobayashi, Masaharu Noda
Nature Neuroscience, 20, 230-241 (2017)
要 約
水分あるいは塩分への欲求を正しく制御するため,脳はつねに体液の状態をモニターしている.アンジオテンシンIIは水分および塩分への両方の欲求を司令するが,体液の状態に応じて,選択的にそれらのうちの一方への欲求を,あるいは,両方への欲求を亢進する神経機構は明らかにされていない.この研究において,筆者らは,光遺伝学的な手法を用いて,水分および塩分への欲求が脳弓下器官のアンジオテンシンII受容体陽性の異なる2つの興奮性ニューロンの集団により制御されることを明らかにした.水分への欲求は終板脈管器官に投射するニューロンが,一方,塩分への欲求は腹側分界条床核に投射するニューロンが担っていた.水分への欲求を制御するニューロンは,塩分の欠乏した状態においてコレシストキニンにより活性化されるGABAニューロンを介して抑制され,塩分への欲求を制御するニューロンは,水分の欠乏した状態においてNa+チャンネルのひとつであるNaxのシグナルにより別のGABAニューロンを介して抑制されていた.脳弓下器官におけるこのような水分と塩分の選択的な摂取の制御機構は,生命の維持に必要な体液の恒常性の実現に貢献すると考えられる.
はじめに
ヒトを含む陸生動物はつねに水分あるいは塩分の欠乏という危険にさらされている.水分の欠乏した状態においては体液のNa+の濃度が上昇する結果,動物は渇きをおぼえるとともに塩分を避ける1,2).一方,塩分が欠乏すると塩分への欲求が高まるが,これはCl-でなくNa+の摂取によってのみ充足される3).体液の状態に応じたこれらの欲求の正しい制御は生命の維持にとり必須のしくみとなっている.
Na+は体液の主要な陽イオンであり,浸透圧を決める主要な因子である.体液におけるNa+の濃度は体液の浸透圧を生理的な範囲に維持するため脳においてつねにモニターされている.脳におけるモニターの場は血液-脳関門を欠く脳室周囲器官であると推測されていた4).筆者らは,これまでに,Na+チャンネルのひとつであるNaxが感覚性の脳室周囲器官である脳弓下器官および終板脈管器官の特殊なグリア細胞に特異的に発現しており,これがNa+の濃度の上昇を感知する脳のNa+センサーであることを明らかにしてきた2,5).Naxノックアウトマウスを用いた一連の解析から,脳弓下器官が塩分への欲求の制御のためのNa+の濃度のモニターの中枢であり,Naxの活性化はグリア細胞からの乳酸の放出の増加につながり,これがGABAニューロンの活性化を誘導することが明らかにされた6).筆者らは,このGABAニューロンが塩分の摂取の行動をつかさどるニューロンの活性を制御すると推定していた7).
アンジオテンシンIIは血管の収縮を促進するペプチドホルモンであり,血圧の制御を担うホルモンのひとつである.一方で,アンジオテンシンIIを脳室に投与すると水分および塩分の両方の摂取を促進することが知られていた8).脳室周囲器官にはアンジオテンシンII受容体(ヒトにおいてはAT1およびAT2,マウスにおいてはAT1a,AT1b,AT2)が豊富に分布しており,血液におけるアンジオテンシンIIシグナルの中枢における受容部位と考えられてきた9).
1.AT1aシグナルの水分あるいは塩分への欲求における役割
マウスを水分,塩分,あるいは,その両方の欠乏した状態にした.水分の欠乏した状態としては2日間にわたる絶水,水分および塩分の両方が欠乏した状態としては利尿剤の処理,塩分の欠乏した状態としては利尿剤の処理ののち水分のみを補給した.すべての条件において,血液中のアンジオテンシンIIは同じ程度に上昇した.野生型マウスおよびAT1aノックアウトマウスにおいて水分および塩分の摂取の行動を調べた結果,塩分の摂取は完全にAT1aに依存的であったのに対し,水分の摂取は約半分がAT1aに依存性であることが判明した.AT2ノックアウトマウスにおいてもアンジオテンシンIIに依存的に水分の摂取が減少することから10),水分の摂取の残りの半分はAT2陽性のニューロンにより担われると推測された.
2.水分あるいは塩分への欲求と脳弓下器官および終板脈管器官のAT1a陽性のニューロン
塩分の欠乏した状態の脳弓下器官および終板脈管器官において,ニューロンの活性化のマーカーであるFos陽性のニューロンの数が有意に増加し,そのほとんどがAT1a陽性であった.脳室にAT1の阻害剤であるロサルタンを投与したところ,脳弓下器官あるいは終板脈管器官においてニューロンの活性化は起こらず,同時に,塩分の摂取の行動も抑制された.水分の欠乏した状態においては,塩分の欠乏した状態よりもさらにFos陽性のニューロンの数が増加したが,そこにしめるAT1a陽性のニューロンの割合は塩分の欠乏した状態よりも少なかった.また,AT1aノックアウトマウスにおいても,水分の欠乏した状態においてはFos陽性のニューロンが増加し水分の摂取も増加した.脳弓下器官あるいは終板脈管器官のいずれからAT1aを欠失させても,水分への欲求は大きく減少した.脳弓下器官において選択的にAT1aを欠失させると塩分への欲求が消失したが,終板脈管器官における選択的な欠失により塩分への欲求性はまったく影響されなかった.これらの結果から,脳弓下器官のAT1aシグナルが水分および塩分の両方の摂取に関与するのに対し,終板脈管器官のAT1aシグナルは水分の摂取だけに関与することが示された.
3.脳弓下器官から終板脈管器官に投射するニューロンは水分への欲求を制御する
脳弓下器官のAT1a陽性のニューロンはVglut2陽性およびnNOS陽性であったことから,グルタミン酸作動性の興奮性ニューロンであることがわかった.その投射先を調べたところ,終板脈管器官,正中視索前核,腹側分界条床核,室傍核,視索上核と,脳の多くの部位が標的であった.逆行性の標識により調べたところ,脳弓下器官から終板脈管器官に投射するニューロンの約半分がAT1a陽性のニューロンであった.また,水分の欠乏した状態においてFos陽性になるニューロンは脳弓下器官から終板脈管器官に投射するニューロンとよく一致したことから,脳弓下器官から終板脈管器官に投射するニューロンが水分への欲求を担うであろうと推定された.光遺伝学的な手法により脳弓下器官から終板脈管器官に投射するニューロンを抑制したところ,水分への欲求が有意に減少した.対照的に,塩分への欲求はまったく影響されなかった.光遺伝学的な手法により脳弓下器官から終板脈管器官に投射するニューロンを活性化したところ,水分が足りている状態でも急速な水分の摂取の行動が誘発された.しかしながら,塩分の摂取に対しては影響がなかった.これらの結果から,脳弓下器官から終板脈管器官に投射するニューロンが特異的に水分への欲求を制御することが示された(図1).
4.脳弓下器官から腹側分界条床核に投射するニューロンは塩分への欲求を制御する
グルタミン酸作動性の脳弓下器官のニューロンの一部は腹側分界条床核に投射していたが,これらはほとんどすべてがAT1a陽性であり,塩分の欠乏したときにFos陽性になるニューロンであった.両側の腹側分界条床核を電気的に障害したところ,塩分の欠乏した状態における塩分の摂取がいちじるしく減少した.また,脳弓下器官から腹側分界条床核に投射するニューロンにおいて選択的にAT1aを欠失させたところ,欠失の程度に応じて塩分の摂取が減少した.光遺伝学的な手法により脳弓下器官から腹側分界条床核に投射するニューロンを抑制したところ,塩分の摂取が選択的に抑制された.反対に,光遺伝学的な手法により脳弓下器官から腹側分界条床核に投射するニューロンを活性化したところ,水分の欠乏した状態においても塩分の摂取が選択的に上昇した.このとき,NaClに代えてKClを用いた場合には塩分の摂取の増加は認められなかった.したがって,脳弓下器官から腹側分界条床核に投射するニューロンはNaClの摂取を選択的に制御すると結論された(図1).
5.Naxシグナルは脳弓下器官から腹側分界条床核に投射するニューロンの制御に関与する
Na+チャンネルのひとつであるNaxのシグナルとAT1aに依存的な塩分への欲求の制御との関係について解析した.0.3 M NaClと純水による2瓶法により測定したところ,野生型のマウスにおいては水分の欠乏した状態では体液のNa+の濃度が上昇する結果,塩分の摂取がいちじるしく減少した.すでに報告されたように,Naxノックアウトマウスにおいては水分の欠乏した状態においてもこの塩分の忌避は起こらず,0.3 M NaClの摂取の量は水と同じ程度である11)(新着論文レビュー でも掲載).対照的に,AT1aノックアウトマウスおよびNaxとAT1aのダブルノックアウトマウスにおいては,0.3 M NaClはほとんど摂取されなかった.このことから,AT1aの欠損により塩分への欲求は消失することが示された.Naxノックアウトマウスにおいては水分の欠乏した状態にてFos陽性である脳弓下器官から腹側分界条床核に投射するニューロンの数が野生型のマウスと比べ有意に多かった.このことから,脳弓下器官から腹側分界条床核に投射するニューロンの活性がNaxシグナルにより負に制御されることが示唆された(図2).
6.脳弓下器官から腹側分界条床核に投射するニューロンのNa+の濃度による制御
脳弓下器官から腹側分界条床核に投射するニューロンおよびその近傍のGABAニューロンの活性を電気生理学的に調べたところ,Na+の濃度が145 mMの条件において,脳弓下器官から腹側分界条床核に投射するニューロンはアンジオテンシンIIにより活性化したのに対し,GABAニューロンの活性は影響されなかった.Na+の濃度が160 mMの条件においてはGABAニューロンの活性の頻度が上昇したが,この上昇につれてアンジオテンシンIIにより誘導される脳弓下器官から腹側分界条床核に投射するニューロンの活性化は抑制された.このGABAニューロンによる脳弓下器官から腹側分界条床核に投射するニューロンの活性の制御はNaxノックアウトマウスにおいてはみられなかった.また,光遺伝学的な手法によりGABAニューロンを活性化したところ,塩分の欠乏した状態において塩分の摂取が有意に減少した.
7.脳弓下器官から終板脈管器官に投射するニューロンのコレシストキニンによる制御
GABAニューロンの活性化は水分の摂取も少ないながら有意に減少させたことから,脳弓下器官から終板脈管器官に投射するニューロンのGABAニューロンによる制御の可能性が推定された.脳弓下器官から終板脈管器官に投射するニューロンとそれとシナプス結合するGABAニューロンとの関係について調べたところ,Na+の濃度が145 mMの条件において脳弓下器官から終板脈管器官に投射するニューロンの活性は停止していたが,このニューロンの約半数についてはアンジオテンシンIIにより活性化することがわかった.さらに,コレシストキニンがGABAニューロンの活性化を誘導し,脳弓下器官から終板脈管器官に投射するニューロンの活性を制御することが判明した.脳弓下器官におけるコレシストキニンの濃度を調べたところ,Naの欠乏した状態でのみ約9倍の濃度の上昇がみられた.コレシストキニンは脳弓下器官から腹側分界条床核に投射するニューロンと結合するGABAニューロンの活性には影響をおよぼさなかったことから,脳弓下器官から腹側分界条床核に投射するニューロンと,脳弓下器官から終板脈管器官に投射するニューロンとをそれぞれ制御する別のGABAニューロンがあると考えられた(図2).
おわりに
この研究において,筆者らが“水ニューロン”“塩ニューロン”とよぶ脳弓下器官の異なるAT1a陽性のニューロンの集団が,それぞれ水分および塩分への欲求を司令していることが明らかにされた.水ニューロンは脳弓下器官のAT1a陽性の興奮性ニューロンであり,終板脈管器官に投射していた.一方,塩ニューロンは同様のニューロンであるが,腹側分界条床核に投射していた.水ニューロンの活性はNa+の欠乏した状態にて脳弓下器官において濃度の上昇するコレシストキニンがGABAニューロンを介して制御していた.一方,塩ニューロンの活性は水の欠乏した状態においてNa+チャンネルのひとつであるNaxのシグナルが別のGABAニューロンを介して制御していた.このように,アンジオテンシンIIの2つの作用には,体液のNa+の濃度により別々の制御機構が存在していた(図2).水分の摂取については,終板脈管器官にNa+の濃度の上昇に依存するさらなる活性化の機構が存在すると推定されている12).この成果は,体液の状態により水分あるいは塩分への欲求が正しく制御される中枢機構を明らかにしたものであり,体液の制御の異常によりひき起こされるさまざまな疾患の発症機構の理解および治療のための標的の同定につながると考えられる.
文 献
- Stricker, E. M. & Sved, A. F.: Thirst. Nutrition, 16, 821-826 (2000)[PubMed]
- Noda, M. & Sakuta, H.: Central regulation of body-fluid homeostasis. Trends Neurosci., 36, 661-673 (2013)[PubMed]
- Rowland, N. E., Farnbauch, L. J. & Crews, E. C.: Sodium deficiency and salt appetite in ICR:CD1 mice. Physiol. Behav., 80, 629-635 (2004)[PubMed]
- McKinley, M. J., McAllen, R. M., Davern, P. et al.: The sensory circumventricular organs of the mammalian brain. Adv. Anat. Embryol. Cell Biol., 172, III-XII, 1-122, back cover (2003)[PubMed]
- Hiyama, T. Y., Watanabe, E., Ono, K. et al.: Nax channel involved in CNS sodium-level sensing. Nat. Neurosci., 5, 511-512 (2002)[PubMed]
- Shimizu, H., Watanabe, E., Hiyama, T. Y. et al.: Glial Nax channels control lactate signaling to neurons for brain [Na+] sensing. Neuron, 54, 59-72 (2007)[PubMed]
- Noda, M. & Hiyama, T. Y.: The Nax channel: What it is and what it does. Neuroscientist, 21, 399-412 (2015)[PubMed]
- Buggy, J. & Fisher, A. E.: Evidence for a dual central role for angiotensin in water and sodium intake. Nature, 250, 733-72 (1974)[PubMed]
- Premer, C., Lamondin, C., Mitzey, A. et al.: Immunohistochemical localization of AT1a, AT1b, and AT2 angiotensin II receptor subtypes in the rat adrenal, pituitary, and brain with a perspective commentary. Int. J. Hypertens., 2013, 175428 (2013)[PubMed]
- Li, Z., Iwai, M., Wu, L. et al.: Role of AT2 receptor in the brain in regulation of blood pressure and water intake. Am. J. Physiol. Heart Circ. Physiol., 284, H116-H121 (2003)[PubMed]
- Hiyama, T. Y., Yoshida, M., Matsumoto, M. et al.: Endothelin-3 expression in the subfornical organ enhances the sensitivity of Nax, the brain sodium-level sensor, to suppress salt intake. Cell Metab., 17, 507-519 (2013)[PubMed] [新着論文レビュー]
- Sakuta, H., Nishihara, E., Hiyama, T. Y. et al.: Nax signaling evoked by an increase in [Na+] in CSF induces water intake via EET-mediated TRPV4 activation. Am. J. Physiol. Regul. Integr. Comp. Physiol., 311, R299-R306 (2016)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:総合研究大学院大学生命科学研究科博士課程 在学中.
研究テーマ:体液の恒常性の維持にかかわる神経機構.
野田 昌晴(Masaharu Noda)
基礎生物学研究所 教授.
研究室URL:http://niwww3.nibb.ac.jp/
© 2017 松田隆志・野田昌晴 Licensed under CC 表示 2.1 日本
