神経活動の活性化がIGF-Iの血液脳関門の通過をひき起こす
西島 壮
(首都大学東京大学院人間健康科学研究科 ヘルスプロモーションサイエンス学域行動生理学研究室)
email:西島 壮
DOI: 10.7875/first.author.2010.046
Neuronal activity drives localized blood-brain-barrier transport of serum insulin-like growth factor-I into the CNS.
Takeshi Nishijima*, Joaquin Piriz*, Sylvie Duflot*, Ana M. Fernandez, Gema Gaitan, Ulises Gomez-Pinedo, Jose M. Garcia Verdugo, Felix Leroy, Hideaki Soya, Angel Nuñez, Ignacio Torres-Aleman (* These authors contributed equally to this work.)
Neuron, 67, 834-846 (2010)
血液中のIGF-Iは中枢神経系に移行するとニューロンの成長や生存,細胞興奮性の調節といった多様な神経保護作用を発現する.しかしながら,IGF-Iの血液脳関門の通過をひき起こす機構は明らかにされていない.筆者らは,電気刺激,体性感覚刺激,そして,行動刺激により神経活動を活性化させると,活性化した脳部位へのIGF-Iの移行が促進されることを明らかにした.そして,この血液中のIGF-Iの脳内への移行は神経活動の活性化によって放出されるATP,アラキドン酸誘導体などの血管拡張物質により仲介されていた.これらの血管拡張性物質はMMP-9のプロテアーゼ活性を活性化し,IGF-Iの結合タンパク質であるIGFBP-3を分解する.そして,遊離したIGF-Iは血管内皮細胞におけるIGF-I受容体とLRP-1のはたらきによって血液脳関門を通過し脳内に移行する.この神経活動に依存したIGF-Iの脳内への移行は,てんかんの神経新生効果や,神経刺激のリハビリテーション効果,神経活動に対する脳血流の修飾作用といった現象を説明しうる.
脳毛細血管床および脈絡叢にある血液脳関門のはたらきによって脳実質および脳脊髄液への血液中の物質の移動は厳密に制御されている(図1).かつては,血液中のタンパク質はほとんど血液脳関門を通過できないと考えられてきたが,近年,ホルモンや成長因子は血管内皮細胞に発現する受容体やトランスポーターのはたらきによって血液脳関門を通過することが明らかになってきた1).しかしながら,ホルモンや成長因子が血液脳関門を通過することの生理的な意義やその調節機構はいまだ解明されていない.
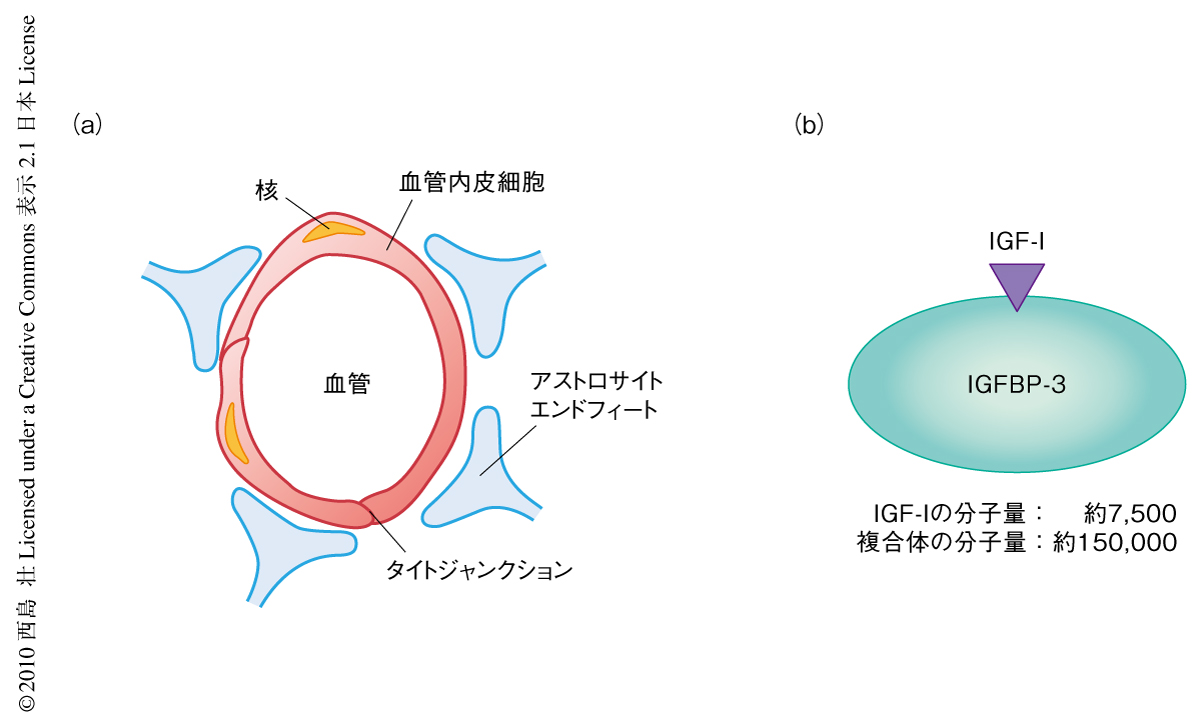
IGF-I(insulin-like growth factor-I,インスリン様成長因子-I)は血液から脳内に移行する成長ホルモンのひとつとして知られている.IGF-Iは多様な神経保護効果をもっており,血管新生,神経新生,ニューロンの興奮性,そして,認知機能などにかかわっている2).したがって,IGF-Iが血液脳関門を通過しなければならないことは明白であり,実際に,血管内皮細胞やアストロサイトのエンドフィートとよばれる血管周囲を取り囲む構造にはIGF-I受容体が発現していて3),IGF-Iが血液脳関門を通過しうる構造をしている.筆者らのグループは,以前,血液中のIGF-Iが脈絡叢から脳脊髄液に取り込まれることを明らかにしたが4),この研究ではさらに,神経活動に依存した積極的かつ局所的なIGF-Iの脳内への取り込み機構が存在するとの仮説をたてた.
この仮説を検証するにあたっては,以下のポイントに着目して研究を展開した.1)IGF-Iは神経活動の活性化により血液から脳内に移行するのか,複数の刺激方法(刺激部位)で実証する.2)神経活動が活性化した部位では局所脳血流量が増加する.したがって,神経活動に依存した血液中のIGF-Iの脳内への移行にはこの局所脳血流量の増加が関与しうる.さらに,局所脳血流量の増加の重要な仲介役であるアストロサイトが5),IGF-Iの脳内への移行の仲介役である可能性が高い.3)血液中ではIGF-IのほとんどがIGFBP-3(insulin-like growth factor-binding protein-3,インスリン様成長因子結合タンパク質3)と結合している.したがって,IGF-Iを脳内に移行させるためにはIGFBP-3を分解してIGF-Iを遊離させる過程が必要なはずである.4)血管内皮細胞にはIGF-I受容体とLRP-1(lipoprotein receptor related protein-1,リポタンパク質受容体関連タンパク質1)とが発現している.したがって,IGF-IはIGF-I受容体とLRP-1とを介したトランスサイトーシスにより血液脳関門を通過する可能性が高い.
まず,血液中のIGF-Iは神経活動が活性化した脳部位へ移行するのかどうかを検証した.ラット動脈にヒト組換えIGF-Iもしくはジゴキシゲニン標識IGF-Iを投与し,活性化させた脳部位でこれらが増加するのかどうか調べた.脳内で検出されたヒト組換えIGF-Iあるいはジゴキシゲニン標識IGF-Iはラットの内因性IGF-Iとは区別でき,血液に由来するものと断定できる.
まず,麻酔下でラット小脳脚を電気刺激したのち,小脳におけるヒト組換えIGF-I濃度を測定した.その結果,電気刺激によって小脳におけるヒト組換えIGF-Iが有意に増加することが認められた.さらに,ほかの脳部位においても同様に神経活動に応じて血液中のIGF-Iが取り込まれるのかどうか確認するため,体性感覚野(バレル皮質)を対側ヒゲ刺激によって刺激したところ,偽刺激群と比較してヒゲ刺激群においてヒト組換えIGF-Iの増加がみられた.なお,ヒゲ刺激はバレル皮質におけるIGF-IをコードするmRNAの発現を増加させなかったが,チロシン残基がリン酸化されたIGF-I受容体は増加しており,刺激部位においてIGF-Iの作用の亢進していることが確認された.さらに,生理的な条件下でも同様に刺激に依存した血液中のIGF-Iの取り込みが生じるかどうかを確認するため,マウスを豊かな環境で短期間(2時間)飼育した.その結果,海馬におけるIGF-I濃度は約60%も増加し,チロシン残基がリン酸化されたIGF-I受容体も増加していた.
以上,小脳脚の電気刺激,バレル皮質の体性感覚刺激,豊かな環境刺激,と刺激方法が異なっても,刺激をうけた脳部位ではIGF-Iが増加すること,さらに,そのIGF-Iは脳内で産生されたものではなく血液から供給されていることが示された.
神経活動に依存して血液中のIGF-Iを脳内に取り込む機構のひとつとして,まず,神経活動が活性化した部位では局所脳血流量が増加することに着目した.ヒゲ刺激によってバレル皮質におけるIGF-Iの取り込みが増加することはさきに述べた.同じ条件下において,バレル皮質の局所脳血流量の変化をレーザードップラー血流計で測定したところ,予想どおり,局所脳血流量は増加していた.この結果は,血液中のIGF-Iの脳内への移行と局所脳血流量の変化とが関連していることを示唆した.
さらに局所脳血流量の関与を調べるため,バレル皮質に微小透析プローブを埋め込み,ヒゲ刺激時のバレル皮質(細胞間質液)におけるヒト組換えIGF-I濃度の変化を調べた.結果はこれまでの結果と一致し,ヒゲ刺激によって透析液中のヒト組換えIGF-Iの濃度は有意に増加した.灌流液に神経活動を阻害するNa+チャネル阻害薬(テトロドトキシン)をくわえると,ヒゲ刺激によるヒト組換えIGF-I濃度の増加は完全に抑制された.同様に,血管拡張を阻害するためNO合成酵素阻害薬(L-NAME)をくわえてもヒト組換えIGF-I濃度の増加は抑制された.これらの結果は,血液中のIGF-Iの脳内への移行には,刺激に応じた神経活動の活性化と,それにともなう局所脳血流量の増加とが必要であることを示していた.
血液脳関門におけるIGF-Iの脳内への移行の機構を解明するためin vitro実験を行った.血液中のIGF-Iはまずエンドサイトーシスにより血管内皮細胞に取り込まれることが最初のステップとなるため,IGF-Iが実際に血管内皮細胞に取り込まれるかどうかを調べた.まず,血液脳関門を構成する主要な3種類の細胞,すなわち,血管内皮細胞,アストロサイト,ニューロンの培養液にビオチン標識ヒト組換えIGF-Iをくわえると,これはすべての細胞に取り込まれることが確認された.とくに,血管内皮細胞で取り込みが顕著だった.また,すべての細胞の細胞質にビオチン標識ヒト組換えIGF-Iの存在することが免疫組織化学的な観察によっても確認された.
つづいて,IGF-IのエンドサイトーシスがIGF-Iの結合タンパク質であるIGFBP-3により阻害されるかどうかを調べた.培養液にIGFBP-3をくわえると血管内皮細胞におけるIGF-Iのエンドサイトーシスは有意に抑制された.逆に,IGFBP-3分解酵素であるPSA(prostate-specific antigen,前立腺特異抗原)をくわえるとIGF-Iのエンドサイトーシスは劇的に促進された.ドミナントネガティブIGF-I受容体を遺伝子導入した血管内皮細胞において,あるいは,IGF-I受容体阻害薬を用いた場合,IGF-Iのエンドサイトーシスは抑制されたことから,このエンドサイトーシスはIGF-I受容体を介していることが示された.PSAは脳内で合成されるプロテアーゼではないため,IGFBP-3を分解する内因性のプロテアーゼの有力候補としてMMP-9(matrix metalloproteinase-9,マトリックスメタロプロテイナーゼ9)に着目した.MMP-9は脳内の血管内皮細胞にも存在し,さらに重要なことには,神経活動の活性化によってそのプロテアーゼ活性の高まることが知られている6).予想どおり,活性型MMP-9を培養液にくわえると血管内皮細胞へのIGF-Iのエンドサイトーシスは有意に促進された.
エンドサイトーシスによって血管内皮細胞に取り込まれたIGF-Iは脳実質側に放出されなくてはならない.そこで,IGF-Iが血液脳関門をトランスサイトーシスによって通過するのかどうかを検証しその調節機構の詳細にせまった.実験ではin vitro血液脳関門モデルを用いた.このモデルはダブルチャンバー構造からなり,両方のチャンバーのあいだにアストロサイトと血管内皮細胞とを培養することで人工的な血液脳関門を再現した.上部チャンバー(血液側)にビオチン標識ヒト組換えIGF-Iをくわえ,下部チャンバー(脳実質側)においてこれを定量することでIGF-Iのトランスサイトーシスを評価した.
この研究の中核となる仮説は,神経活動の活性化が血液中のIGF-Iの脳内への移行をひき起こすことである.そこで,グルタミン酸を下部チャンバーにくわえたところIGF-Iのトランスサイトーシスは有意に促進された.このIGF-IのトランスサイトーシスはMMP-9阻害薬によって阻害され,なおかつ,グルタミン酸によってMMP-9のプロテアーゼ活性の高まることも確認された.以上の結果より,神経活動の活性化によりIGF-Iは血液脳関門をトランスサイトーシスによって通過し,さらに,その調節には神経活動の活性化によるMMP-9のプロテアーゼ活性の増加が関与することが示された.
神経活動の活性化にともない,アストロサイトからはプロスタグランジンE2,エポキシエイコサトリエン酸,ATPなどの血管拡張物質が放出される7).興味深いことに,これら血管拡張物質によってもIGF-Iのトランスサイトーシスは促進され,その作用はMMP-9阻害薬によって阻害された.これら血管拡張物質は局所脳血流量を増加させるだけでなく,MMP-9のプロテアーゼ活性を高めることによって,神経活動に依存したIGF-Iの脳内への移行の調節に関与していることが明らかになった.
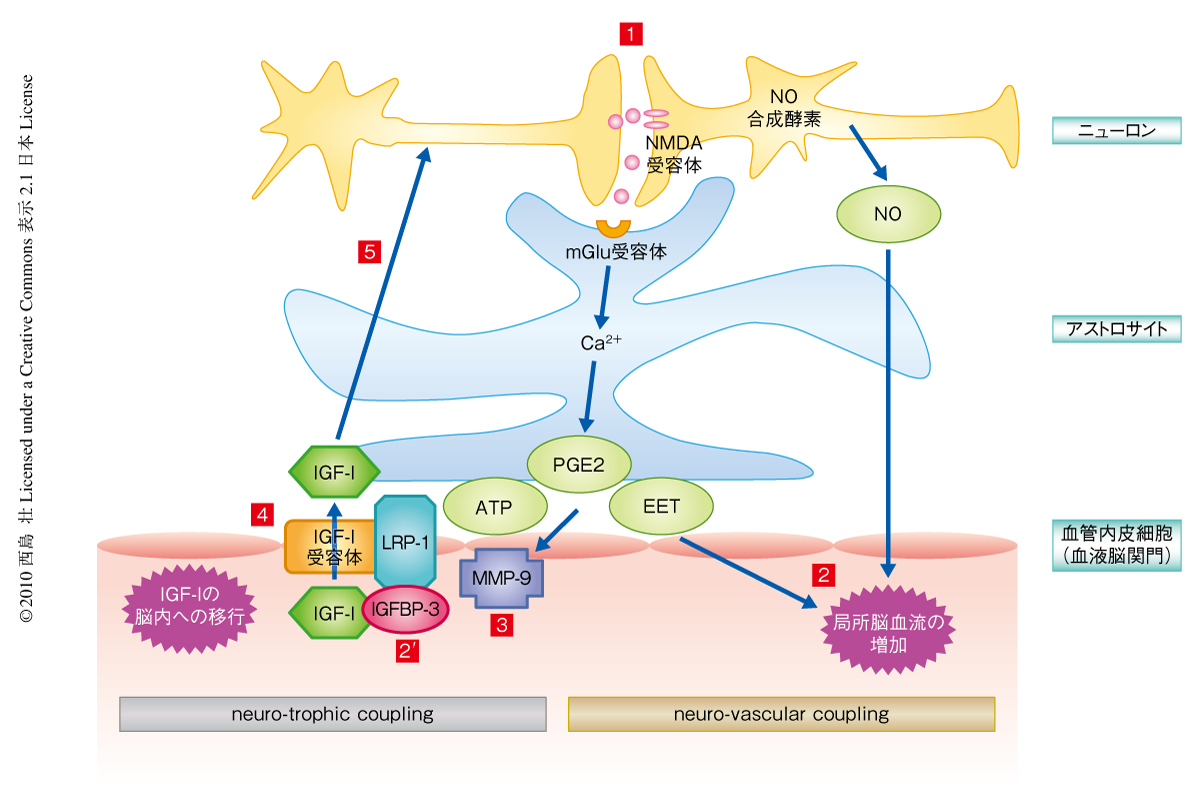
この研究によって明らかになった血液中のIGF-Iの脳内への移行の機構を示す(図2).神経活動に依存して血液中のIGF-Iを脳内に取り込む機構が,活性化した脳部位に酸素や栄養基質を供給するため局所脳血流量を増加させる機構(neuro-vascular coupling)ときわめて似ていたことから,筆者らは,神経活動に依存した血液中のIGF-Iの脳内への移行の機構をneuro-trophic coupling(神経-栄養効果カップリング)と名づけた.
神経活動の活性化はニューロンの抗酸化能力を高めるなど神経保護効果をもたらすが8),この研究ではさらに,神経活動の活性化は多様な神経保護効果をもつIGF-Iを血液中から脳内に取り込む引き金となることが明らかとなった.この事実は,神経活動に依存してひき起こされる生理的な反応が脳機能を維持するためきわめて重要であるというこれまでの知見を強く支持する.豊かな社会的活動や教育そして運動が脳機能に有益であることや9),てんかんや電気刺激療法に神経新生を促進する作用があり10),また,神経刺激にはリハビリテーション効果のあることも知られている11).これらの背景には共通して,この研究で明らかとなったneuro-trophic couplingによるIGF-Iの脳内への移行が関与しているのかもしれない.今後,脳損傷部位の神経刺激によるニューロリハビリテーションなどにもこの機構が応用される可能性がある.
略歴:2006年 筑波大学大学院人間総合科学研究科を修了,同 博士研究員,国際科学振興財団 専任研究員,スペインCajal Institute外国人若手研究員を経て,2009年より首都大学東京大学院人間健康科学研究科 助教.
研究テーマ:運動が脳機能に及ぼす効果の神経科学的解明.
抱負:ストレス社会を生き抜くための運動の重要性を明らかにすべく,運動が脳機能に及ぼす効果について神経科学的なアプローチにより研究している.趣味・特技はバドミントン.
© 2010 西島 壮 Licensed under CC 表示 2.1 日本
(首都大学東京大学院人間健康科学研究科 ヘルスプロモーションサイエンス学域行動生理学研究室)
email:西島 壮
DOI: 10.7875/first.author.2010.046
Neuronal activity drives localized blood-brain-barrier transport of serum insulin-like growth factor-I into the CNS.
Takeshi Nishijima*, Joaquin Piriz*, Sylvie Duflot*, Ana M. Fernandez, Gema Gaitan, Ulises Gomez-Pinedo, Jose M. Garcia Verdugo, Felix Leroy, Hideaki Soya, Angel Nuñez, Ignacio Torres-Aleman (* These authors contributed equally to this work.)
Neuron, 67, 834-846 (2010)
要 約
血液中のIGF-Iは中枢神経系に移行するとニューロンの成長や生存,細胞興奮性の調節といった多様な神経保護作用を発現する.しかしながら,IGF-Iの血液脳関門の通過をひき起こす機構は明らかにされていない.筆者らは,電気刺激,体性感覚刺激,そして,行動刺激により神経活動を活性化させると,活性化した脳部位へのIGF-Iの移行が促進されることを明らかにした.そして,この血液中のIGF-Iの脳内への移行は神経活動の活性化によって放出されるATP,アラキドン酸誘導体などの血管拡張物質により仲介されていた.これらの血管拡張性物質はMMP-9のプロテアーゼ活性を活性化し,IGF-Iの結合タンパク質であるIGFBP-3を分解する.そして,遊離したIGF-Iは血管内皮細胞におけるIGF-I受容体とLRP-1のはたらきによって血液脳関門を通過し脳内に移行する.この神経活動に依存したIGF-Iの脳内への移行は,てんかんの神経新生効果や,神経刺激のリハビリテーション効果,神経活動に対する脳血流の修飾作用といった現象を説明しうる.
はじめに
脳毛細血管床および脈絡叢にある血液脳関門のはたらきによって脳実質および脳脊髄液への血液中の物質の移動は厳密に制御されている(図1).かつては,血液中のタンパク質はほとんど血液脳関門を通過できないと考えられてきたが,近年,ホルモンや成長因子は血管内皮細胞に発現する受容体やトランスポーターのはたらきによって血液脳関門を通過することが明らかになってきた1).しかしながら,ホルモンや成長因子が血液脳関門を通過することの生理的な意義やその調節機構はいまだ解明されていない.
IGF-I(insulin-like growth factor-I,インスリン様成長因子-I)は血液から脳内に移行する成長ホルモンのひとつとして知られている.IGF-Iは多様な神経保護効果をもっており,血管新生,神経新生,ニューロンの興奮性,そして,認知機能などにかかわっている2).したがって,IGF-Iが血液脳関門を通過しなければならないことは明白であり,実際に,血管内皮細胞やアストロサイトのエンドフィートとよばれる血管周囲を取り囲む構造にはIGF-I受容体が発現していて3),IGF-Iが血液脳関門を通過しうる構造をしている.筆者らのグループは,以前,血液中のIGF-Iが脈絡叢から脳脊髄液に取り込まれることを明らかにしたが4),この研究ではさらに,神経活動に依存した積極的かつ局所的なIGF-Iの脳内への取り込み機構が存在するとの仮説をたてた.
この仮説を検証するにあたっては,以下のポイントに着目して研究を展開した.1)IGF-Iは神経活動の活性化により血液から脳内に移行するのか,複数の刺激方法(刺激部位)で実証する.2)神経活動が活性化した部位では局所脳血流量が増加する.したがって,神経活動に依存した血液中のIGF-Iの脳内への移行にはこの局所脳血流量の増加が関与しうる.さらに,局所脳血流量の増加の重要な仲介役であるアストロサイトが5),IGF-Iの脳内への移行の仲介役である可能性が高い.3)血液中ではIGF-IのほとんどがIGFBP-3(insulin-like growth factor-binding protein-3,インスリン様成長因子結合タンパク質3)と結合している.したがって,IGF-Iを脳内に移行させるためにはIGFBP-3を分解してIGF-Iを遊離させる過程が必要なはずである.4)血管内皮細胞にはIGF-I受容体とLRP-1(lipoprotein receptor related protein-1,リポタンパク質受容体関連タンパク質1)とが発現している.したがって,IGF-IはIGF-I受容体とLRP-1とを介したトランスサイトーシスにより血液脳関門を通過する可能性が高い.
1.神経活動の活性化は血液中のIGF-Iの脳内への取り込みを刺激する
まず,血液中のIGF-Iは神経活動が活性化した脳部位へ移行するのかどうかを検証した.ラット動脈にヒト組換えIGF-Iもしくはジゴキシゲニン標識IGF-Iを投与し,活性化させた脳部位でこれらが増加するのかどうか調べた.脳内で検出されたヒト組換えIGF-Iあるいはジゴキシゲニン標識IGF-Iはラットの内因性IGF-Iとは区別でき,血液に由来するものと断定できる.
まず,麻酔下でラット小脳脚を電気刺激したのち,小脳におけるヒト組換えIGF-I濃度を測定した.その結果,電気刺激によって小脳におけるヒト組換えIGF-Iが有意に増加することが認められた.さらに,ほかの脳部位においても同様に神経活動に応じて血液中のIGF-Iが取り込まれるのかどうか確認するため,体性感覚野(バレル皮質)を対側ヒゲ刺激によって刺激したところ,偽刺激群と比較してヒゲ刺激群においてヒト組換えIGF-Iの増加がみられた.なお,ヒゲ刺激はバレル皮質におけるIGF-IをコードするmRNAの発現を増加させなかったが,チロシン残基がリン酸化されたIGF-I受容体は増加しており,刺激部位においてIGF-Iの作用の亢進していることが確認された.さらに,生理的な条件下でも同様に刺激に依存した血液中のIGF-Iの取り込みが生じるかどうかを確認するため,マウスを豊かな環境で短期間(2時間)飼育した.その結果,海馬におけるIGF-I濃度は約60%も増加し,チロシン残基がリン酸化されたIGF-I受容体も増加していた.
以上,小脳脚の電気刺激,バレル皮質の体性感覚刺激,豊かな環境刺激,と刺激方法が異なっても,刺激をうけた脳部位ではIGF-Iが増加すること,さらに,そのIGF-Iは脳内で産生されたものではなく血液から供給されていることが示された.
2.神経活動に依存したIGF-Iの脳内への移行には局所脳血流量の増加が必要である
神経活動に依存して血液中のIGF-Iを脳内に取り込む機構のひとつとして,まず,神経活動が活性化した部位では局所脳血流量が増加することに着目した.ヒゲ刺激によってバレル皮質におけるIGF-Iの取り込みが増加することはさきに述べた.同じ条件下において,バレル皮質の局所脳血流量の変化をレーザードップラー血流計で測定したところ,予想どおり,局所脳血流量は増加していた.この結果は,血液中のIGF-Iの脳内への移行と局所脳血流量の変化とが関連していることを示唆した.
さらに局所脳血流量の関与を調べるため,バレル皮質に微小透析プローブを埋め込み,ヒゲ刺激時のバレル皮質(細胞間質液)におけるヒト組換えIGF-I濃度の変化を調べた.結果はこれまでの結果と一致し,ヒゲ刺激によって透析液中のヒト組換えIGF-Iの濃度は有意に増加した.灌流液に神経活動を阻害するNa+チャネル阻害薬(テトロドトキシン)をくわえると,ヒゲ刺激によるヒト組換えIGF-I濃度の増加は完全に抑制された.同様に,血管拡張を阻害するためNO合成酵素阻害薬(L-NAME)をくわえてもヒト組換えIGF-I濃度の増加は抑制された.これらの結果は,血液中のIGF-Iの脳内への移行には,刺激に応じた神経活動の活性化と,それにともなう局所脳血流量の増加とが必要であることを示していた.
3.IGF-Iはエンドサイトーシスによって血管内皮細胞に取り込まれる
血液脳関門におけるIGF-Iの脳内への移行の機構を解明するためin vitro実験を行った.血液中のIGF-Iはまずエンドサイトーシスにより血管内皮細胞に取り込まれることが最初のステップとなるため,IGF-Iが実際に血管内皮細胞に取り込まれるかどうかを調べた.まず,血液脳関門を構成する主要な3種類の細胞,すなわち,血管内皮細胞,アストロサイト,ニューロンの培養液にビオチン標識ヒト組換えIGF-Iをくわえると,これはすべての細胞に取り込まれることが確認された.とくに,血管内皮細胞で取り込みが顕著だった.また,すべての細胞の細胞質にビオチン標識ヒト組換えIGF-Iの存在することが免疫組織化学的な観察によっても確認された.
つづいて,IGF-IのエンドサイトーシスがIGF-Iの結合タンパク質であるIGFBP-3により阻害されるかどうかを調べた.培養液にIGFBP-3をくわえると血管内皮細胞におけるIGF-Iのエンドサイトーシスは有意に抑制された.逆に,IGFBP-3分解酵素であるPSA(prostate-specific antigen,前立腺特異抗原)をくわえるとIGF-Iのエンドサイトーシスは劇的に促進された.ドミナントネガティブIGF-I受容体を遺伝子導入した血管内皮細胞において,あるいは,IGF-I受容体阻害薬を用いた場合,IGF-Iのエンドサイトーシスは抑制されたことから,このエンドサイトーシスはIGF-I受容体を介していることが示された.PSAは脳内で合成されるプロテアーゼではないため,IGFBP-3を分解する内因性のプロテアーゼの有力候補としてMMP-9(matrix metalloproteinase-9,マトリックスメタロプロテイナーゼ9)に着目した.MMP-9は脳内の血管内皮細胞にも存在し,さらに重要なことには,神経活動の活性化によってそのプロテアーゼ活性の高まることが知られている6).予想どおり,活性型MMP-9を培養液にくわえると血管内皮細胞へのIGF-Iのエンドサイトーシスは有意に促進された.
4.血液脳関門におけるIGF-Iのトランスサイトーシス
エンドサイトーシスによって血管内皮細胞に取り込まれたIGF-Iは脳実質側に放出されなくてはならない.そこで,IGF-Iが血液脳関門をトランスサイトーシスによって通過するのかどうかを検証しその調節機構の詳細にせまった.実験ではin vitro血液脳関門モデルを用いた.このモデルはダブルチャンバー構造からなり,両方のチャンバーのあいだにアストロサイトと血管内皮細胞とを培養することで人工的な血液脳関門を再現した.上部チャンバー(血液側)にビオチン標識ヒト組換えIGF-Iをくわえ,下部チャンバー(脳実質側)においてこれを定量することでIGF-Iのトランスサイトーシスを評価した.
この研究の中核となる仮説は,神経活動の活性化が血液中のIGF-Iの脳内への移行をひき起こすことである.そこで,グルタミン酸を下部チャンバーにくわえたところIGF-Iのトランスサイトーシスは有意に促進された.このIGF-IのトランスサイトーシスはMMP-9阻害薬によって阻害され,なおかつ,グルタミン酸によってMMP-9のプロテアーゼ活性の高まることも確認された.以上の結果より,神経活動の活性化によりIGF-Iは血液脳関門をトランスサイトーシスによって通過し,さらに,その調節には神経活動の活性化によるMMP-9のプロテアーゼ活性の増加が関与することが示された.
神経活動の活性化にともない,アストロサイトからはプロスタグランジンE2,エポキシエイコサトリエン酸,ATPなどの血管拡張物質が放出される7).興味深いことに,これら血管拡張物質によってもIGF-Iのトランスサイトーシスは促進され,その作用はMMP-9阻害薬によって阻害された.これら血管拡張物質は局所脳血流量を増加させるだけでなく,MMP-9のプロテアーゼ活性を高めることによって,神経活動に依存したIGF-Iの脳内への移行の調節に関与していることが明らかになった.
5.血液中のIGF-Iの脳内への移行の機構
この研究によって明らかになった血液中のIGF-Iの脳内への移行の機構を示す(図2).神経活動に依存して血液中のIGF-Iを脳内に取り込む機構が,活性化した脳部位に酸素や栄養基質を供給するため局所脳血流量を増加させる機構(neuro-vascular coupling)ときわめて似ていたことから,筆者らは,神経活動に依存した血液中のIGF-Iの脳内への移行の機構をneuro-trophic coupling(神経-栄養効果カップリング)と名づけた.
おわりに
神経活動の活性化はニューロンの抗酸化能力を高めるなど神経保護効果をもたらすが8),この研究ではさらに,神経活動の活性化は多様な神経保護効果をもつIGF-Iを血液中から脳内に取り込む引き金となることが明らかとなった.この事実は,神経活動に依存してひき起こされる生理的な反応が脳機能を維持するためきわめて重要であるというこれまでの知見を強く支持する.豊かな社会的活動や教育そして運動が脳機能に有益であることや9),てんかんや電気刺激療法に神経新生を促進する作用があり10),また,神経刺激にはリハビリテーション効果のあることも知られている11).これらの背景には共通して,この研究で明らかとなったneuro-trophic couplingによるIGF-Iの脳内への移行が関与しているのかもしれない.今後,脳損傷部位の神経刺激によるニューロリハビリテーションなどにもこの機構が応用される可能性がある.
文 献
- Banks, W. A.: Denial versus dualism: the blood-brain barrier as an interface of the gut-brain axis. Endocrinology, 147, 2609-2610 (2006)[PubMed]
- Russo, V. C., Gluckman, P. D., Feldman, E. L. et al.: The insulin-like growth factor system and its pleiotropic functions in brain. Endocr. Rev., 26, 916-943 (2005)[PubMed]
- Garcia-Segura, L. M., Perez, J., Pons, S. et al.: Localization of insulin-like growth factor I (IGF-I)-like immunoreactivity in the developing and adult rat brain. Brain Res., 560, 167-174 (1991)[PubMed]
- Carro, E., Spuch, C., Trejo, J. L. et al.: Choroid plexus megalin is involved in neuroprotection by serum insulin-like growth factor I. J. Neurosci., 25, 10884-10893 (2005)[PubMed]
- Zonta, M., Angulo, M. C., Gobbo, S. et al.: Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation. Nat. Neurosci., 6, 43-50 (2003)[PubMed]
- Michaluk, P. & Kaczmarek, L.: Matrix metalloproteinase-9 in glutamate-dependent adult brain function and dysfunction. Cell Death Differ., 14, 1255-1258 (2007)[PubMed]
- Iadecola, C., Nedergaard, M.: Glial regulation of the cerebral microvasculature. Nat. Neurosci., 10, 1369-1376 (2007)[PubMed]
- Papadia, S., Soriano, F. X., Leveille, F. et al.: Synaptic NMDA receptor activity boosts intrinsic antioxidant defenses. Nat. Neurosci., 11, 476-487 (2008)[PubMed]
- Fratiglioni, L., Paillard-Borg, S. & Winblad, B.: An active and socially integrated lifestyle in late life might protect against dementia. Lancet Neurol., 3, 343-353 (2004)[PubMed]
- Kempermann, G.: Regulation of adult hippocampal neurogenesis - implications for novel theories of major depression. Bipolar Disord., 4, 17-33 (2002)[PubMed]
- Kaelin-Lang. A.: Enhancing rehabilitation of motor deficits with peripheral nerve stimulation. NeuroRehabilitation, 23, 89-93 (2008)[PubMed]
著者プロフィール
略歴:2006年 筑波大学大学院人間総合科学研究科を修了,同 博士研究員,国際科学振興財団 専任研究員,スペインCajal Institute外国人若手研究員を経て,2009年より首都大学東京大学院人間健康科学研究科 助教.
研究テーマ:運動が脳機能に及ぼす効果の神経科学的解明.
抱負:ストレス社会を生き抜くための運動の重要性を明らかにすべく,運動が脳機能に及ぼす効果について神経科学的なアプローチにより研究している.趣味・特技はバドミントン.
© 2010 西島 壮 Licensed under CC 表示 2.1 日本
