器官の正確な形状は細胞の成長の時空間的な平均化により達成される
津川 暁1・Arezki Boudaoud 2・Adrienne H. K. Roeder 3
(1北海道大学電子科学研究所 附属社会創造数学研究センターデータ数理研究分野,2フランスLyon大学Laboratory of Plant Reproduction and Development,3米国Cornell大学Weill Institute for Cell and Molecular Biology)
email:津川 暁
DOI: 10.7875/first.author.2016.079
Variable cell growth yields reproducible organ development through spatiotemporal averaging.
Lilan Hong, Mathilde Dumond, Satoru Tsugawa, Aleksandra Sapala, Anne-Lise Routier-Kierzkowska, Yong Zhou, Catherine Chen, Annamaria Kiss, Mingyuan Zhu, Olivier Hamant, Richard S. Smith, Tamiki Komatsuzaki, Chun-Biu Li, Arezki Boudaoud, Adrienne H. K. Roeder
Developmental Cell, 38, 15-32 (2016)
発生生物学においてもっとも重要な未解決の問題のひとつは,生命システムが正確な大きさや形状をどのように獲得するのかである.この問題の難易度が非常に高いのは,器官における細胞のランダムな挙動のためである.たとえば,植物の器官は最終的な形状は全体としてほとんど同じなのに,隣接した細胞の成長や分裂の割合は異なる.筆者らは,植物の器官は細胞の不均一性にもかかわらずどのように同じ形状を獲得するのかという問題に注目し,モデル植物であるシロイヌナズナのがく片を詳細に調べた.遺伝子工学的な手法および数理モデルにより,正確な形状に到達するためにはむしろ細胞の成長のランダム性が必要とされているという,いっけん直感に反する結果が得られた.また,成長の過程において活性酸素種ががく片の先端部から基部へと移行して細胞の成長を段階的に制御することにより,器官の大きさが決定される可能性のあることが明らかにされた.
なぜゾウは大きくネズミは小さいのか.なぜヒトの2つの手の大きさは1%しか違わないのか.発生生物学はいっけん単純そうなこれらの疑問にいまだ答えることができない.発生生物学においてもっとも重要な未解決の問題のひとつは,生命システムがいかに正確な大きさや形状を獲得し,かつ,いかに成長を停止するタイミングを知るのかである1).植物はほとんど同形の花をつける性質があり,顕微鏡による観察が比較的容易であるため,この問題に対する格好の研究材料であるといえる.これまでの研究により,すべての花を大きく(あるいは,小さく)するような遺伝子や,すべての細胞を大きく(あるいは,小さく)するような遺伝子は同定されていたが2,3),植物がどのように成長を停止しほぼ同じ大きさや形状を獲得するのかはよくわかっていなかった.この研究においては,ヒトの2つの手でいえば指紋や皮下脂肪がそれぞれ異なるように,植物の器官において細胞の大きさや形状が個々の花において異なるというミクロな観察が手がかりになった3,4).
シロイヌナズナはひとつの花弁に同じ大きさの4つのがく片をもち,ひとつの個体は100個以上の花を咲かせる.すなわち,単一の個体において多様性を調べることができる.これは一般に,動物においては実現できない.がく片は花の中心の器官をとりかこみ保護する役割をもつため,がく片の大きさが一様であることは機能的に決定的に重要である.くわえて,花弁の大きさは環境の変化に対して相対的に不変であるため,器官の大きさを支配する固有のしくみを調べることができる.さらに,がく片は最外の器官であるため,成長の過程をイメージングすることが可能である.変異体の遺伝学的なスクリーニングにより,がく片の大きさおよび花弁のほかの器官の大きさが多様に変化する変異体をみつけ,花芽が異常な開き方をすることを見い出した.それらの変異体のうち,ftsh4変異体について詳細に調べた.
環境の変化が損傷や機能な損失を最小限にとどめて処理されるとき,そのシステムあるいは生物は頑健性をもつという.形態形成においては,擾乱に対し表現型が変化せず同じ形状を形成する性質を頑健性とよぶ.先行研究における遺伝学的なスクリーニングにおいては,すべての器官の大きさを制御することはできていたが,器官がどのように正確な大きさや形状をとるのかをみとおすことはできていなかった.そこで,器官の頑健性に直接に影響をおよぼすような変異体をスクリーニングした.器官の大きさを制御するしくみを知るためには,器官の大きさが平均値を中心にランダムに分布するような変異体において,何が制御因子であるかを同定する必要があるからである.シロイヌナズナにおいてAAA-ATPase活性およびメタロプロテアーゼ活性をもつ膜タンパク質性のプロテアーゼのひとつであるFtsH4の変異体においては,器官の大きさおよび形状が変化した.これまで,出芽酵母において頑健性に欠陥をもつ変異体が報告されていたが5,6),多細胞性の真核生物についての報告はなかった.
なぜftsh4変異体において器官の大きさの頑健性が異常をきたすのかを理解するため,数理モデルを構築した.先行研究より7,8),成長の局所における制御と器官の形状をつなぐ成長をモデルとした.がく片の先端部から基部にかけて成長が遅くなるという観察にもとづき,先端部からある閾値までの領域において成長を遅くする仮想的なシグナルをくわえた.シグナルは下方に伝達され最基部に到達すると停止するとした.このいわゆる“成長前線”モデルにおいて,シミュレーションによるがく片の大きさは実際と同様に一定になった.
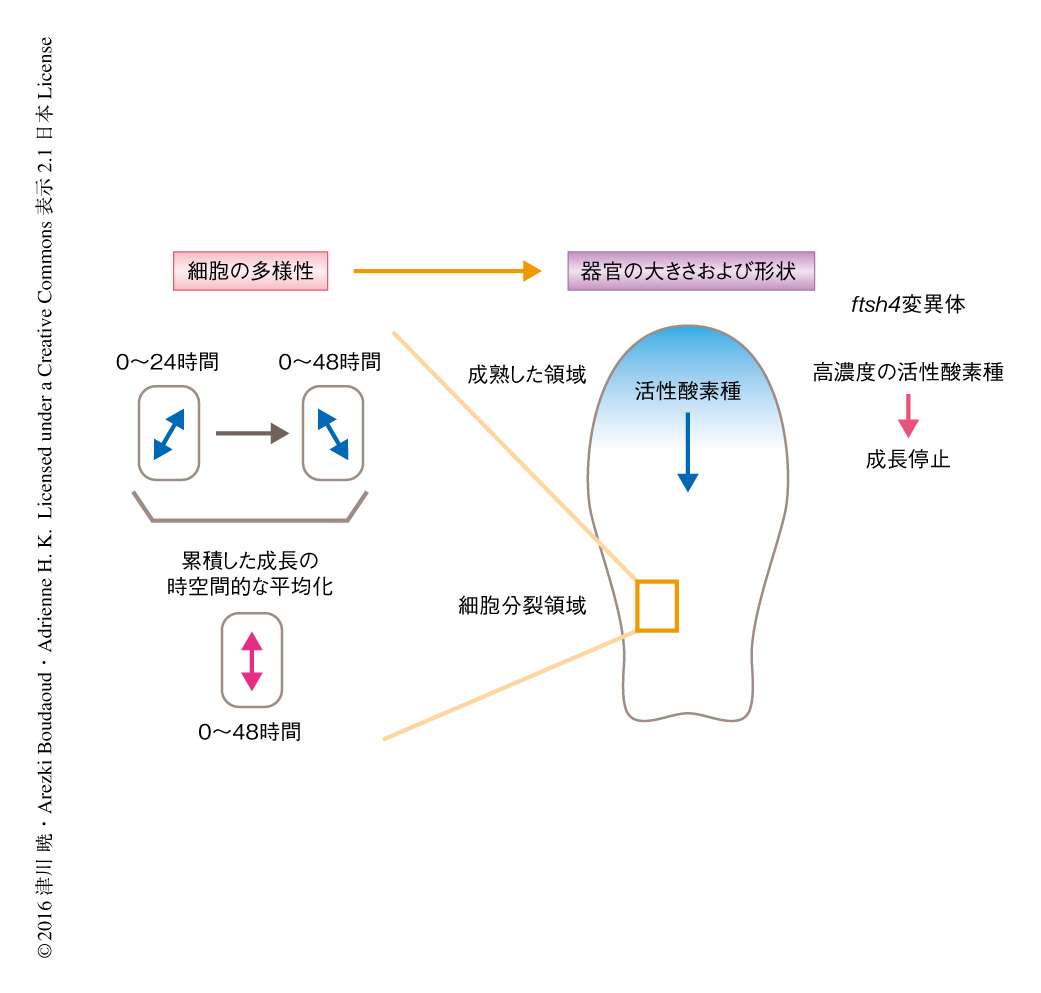
しかしながら,この成長前線モデルは成長したがく片において観察される細胞の多様性を考慮に入れていなかった.そこで,最初の疑問である細胞の多様性からどのようにして器官が制御されるのかという問題にたちもどり,がく片を“風船”にみたて,がく片の場所に依存した異なる弾性率をランダムにあたえた.いい換えると,弾性の異なる細胞をランダムに配置した.細胞を平衡に到達するまでふくらませ,がく片がどういう形状に到達するかを評価した.初期の時刻においてランダムに弾性率をあたえたモデルにおいては,いびつな“変異体もどき”の初期の弾性率の空間分布に応じた多様な形状をとった.しかしながら,成長の経過とともに時間的にランダムに弾性率をふりなおしたモデルにおいては,がく片は均一な形状をとった.このことから,成長の過程における細胞の多様性の時空間的な平均化が正常な器官を形成するうえで重要であることが示唆された.
野生型のがく片において細胞の成長を観察したところ,12時間の間隔でみたときには時間的にも空間的にも多様であったが,細胞およびその娘細胞の成長の方向を48時間の間隔で累積したところほぼそろい,がく片の成長の方向と一致した(図1a).このことから,野生型のがく片は成長の方向を時空間的に平均化するしくみをもつことが示された.一方,ftsh4変異体のがく片の細胞は平均化の過程に欠陥が生じていた.ftsh4変異体の成長の方向は野生型よりも小さな時空間的な変動を示したが,48時間の間隔で累積したところ非常に多様なままであり,がく片の成長の方向とは一致しなかった(図1b).数理モデルおよび細胞壁の弾性の測定から,細胞を弾性体と考え,硬い細胞は成長が遅く柔らかい細胞は成長が速いと想定すると,野生型においては細胞の成長の多様性が高く,ftsh4変異体においては細胞の成長の多様性が低い可能性が示唆された.そこで,隣接した細胞の成長の空間的な多様性を定量化する手法を開発したところ,実際に,野生型のほうがftsh4変異体よりも細胞の成長の空間的な多様性の高いことが明らかにされた.また,野生型とftsh4変異体の器官の形状を比較するため形状の多様性を定量的に解析したところ,ftsh4変異体は野生型と比べがく片の大きさおよび形状の変動性がいちじるしく高かった.

以上の結果から,野生型のがく片においてはより多様な細胞の成長があり,その多様性が時空間的に平均化されることが明らかにされた.時空間的な平均化の考え方は,ショウジョウバエの胚においてhunchback遺伝子の確率的な転写と転写産物の均一な分布を説明する際に提案されているが9),形態形成において議論されたことはない.このような時空間的な平均化によりノイズに対応するしくみは,ほかの多くの生物学的な過程において共通するものなのかもしれない.
すでに,FtsH4変異体はミトコンドリアに障害を起こし,その結果として活性酸素種を増加させ葉の形態異常をひき起こすことが知られている10).そこで,がく片の成長の段階において野生型およびftsh4変異体における活性酸素種の分布を評価した.その結果,野生型においては,がく片の成長とともに活性酸素種の分布ががく片の先端部から基部へと移行することがわかった(図2).一方で,ftsh4変異体においてはそのような活性酸素種の分布の段階的な移行は顕著には認められず,活性酸素種は成長の段階の早期において高濃度に達し,がく片の全体に不均一に分布していた.また,ftsh4変異体において活性酸素種を減少させることによりがく片の大きさおよび形状は均一化され,一方,野生型において活性酸素種を減少させると通常より大きく形状の均一ながく片が形成された.
この研究は,器官の発生は細胞動態の巧みな組織化の連鎖により起こるとされてきた従来の発生生物学の基本的な問題に挑戦し,生命の正常な機能のひとつとして無秩序から秩序を創出するしくみを提案した.これまで,観察の結果から除くべきと考えられてきた細胞のノイズやゆらぎを,今後は,むしろ積極的に調べていくことにより形態形成にひそむしくみが明らかにされると考えている.また,この考え方はほかの生物にも適用できることから,研究の進んでいるショウジョウバエの羽や大腸菌の形態への応用にくわえ,ヒトを含むより高度な多細胞生物の大きさや形状を決める基本原理となっていることが期待される.
略歴:2013年 早稲田大学理工学術院先進理工学研究科博士課程 修了,2014年 北海道大学電子科学研究所 博士研究員を経て,2015年より同 特任助教.
研究テーマ:植物の器官の形態形成にひそむ多様性および不均一性の定量化.
抱負:形態形成,地震,乱流などの複雑な自然現象にひそむ普遍的な法則を明らかにしたい.
Arezki Boudaoud
フランスLyon大学Professor.
Adrienne H. K. Roeder
米国Cornell大学Assistant Professor.
© 2016 津川 暁・Arezki Boudaoud・Adrienne H. K. Roeder Licensed under CC 表示 2.1 日本
(1北海道大学電子科学研究所 附属社会創造数学研究センターデータ数理研究分野,2フランスLyon大学Laboratory of Plant Reproduction and Development,3米国Cornell大学Weill Institute for Cell and Molecular Biology)
email:津川 暁
DOI: 10.7875/first.author.2016.079
Variable cell growth yields reproducible organ development through spatiotemporal averaging.
Lilan Hong, Mathilde Dumond, Satoru Tsugawa, Aleksandra Sapala, Anne-Lise Routier-Kierzkowska, Yong Zhou, Catherine Chen, Annamaria Kiss, Mingyuan Zhu, Olivier Hamant, Richard S. Smith, Tamiki Komatsuzaki, Chun-Biu Li, Arezki Boudaoud, Adrienne H. K. Roeder
Developmental Cell, 38, 15-32 (2016)
要 約
発生生物学においてもっとも重要な未解決の問題のひとつは,生命システムが正確な大きさや形状をどのように獲得するのかである.この問題の難易度が非常に高いのは,器官における細胞のランダムな挙動のためである.たとえば,植物の器官は最終的な形状は全体としてほとんど同じなのに,隣接した細胞の成長や分裂の割合は異なる.筆者らは,植物の器官は細胞の不均一性にもかかわらずどのように同じ形状を獲得するのかという問題に注目し,モデル植物であるシロイヌナズナのがく片を詳細に調べた.遺伝子工学的な手法および数理モデルにより,正確な形状に到達するためにはむしろ細胞の成長のランダム性が必要とされているという,いっけん直感に反する結果が得られた.また,成長の過程において活性酸素種ががく片の先端部から基部へと移行して細胞の成長を段階的に制御することにより,器官の大きさが決定される可能性のあることが明らかにされた.
はじめに
なぜゾウは大きくネズミは小さいのか.なぜヒトの2つの手の大きさは1%しか違わないのか.発生生物学はいっけん単純そうなこれらの疑問にいまだ答えることができない.発生生物学においてもっとも重要な未解決の問題のひとつは,生命システムがいかに正確な大きさや形状を獲得し,かつ,いかに成長を停止するタイミングを知るのかである1).植物はほとんど同形の花をつける性質があり,顕微鏡による観察が比較的容易であるため,この問題に対する格好の研究材料であるといえる.これまでの研究により,すべての花を大きく(あるいは,小さく)するような遺伝子や,すべての細胞を大きく(あるいは,小さく)するような遺伝子は同定されていたが2,3),植物がどのように成長を停止しほぼ同じ大きさや形状を獲得するのかはよくわかっていなかった.この研究においては,ヒトの2つの手でいえば指紋や皮下脂肪がそれぞれ異なるように,植物の器官において細胞の大きさや形状が個々の花において異なるというミクロな観察が手がかりになった3,4).
1.器官の大きさおよび形状が多様になるftsh4変異体の同定
シロイヌナズナはひとつの花弁に同じ大きさの4つのがく片をもち,ひとつの個体は100個以上の花を咲かせる.すなわち,単一の個体において多様性を調べることができる.これは一般に,動物においては実現できない.がく片は花の中心の器官をとりかこみ保護する役割をもつため,がく片の大きさが一様であることは機能的に決定的に重要である.くわえて,花弁の大きさは環境の変化に対して相対的に不変であるため,器官の大きさを支配する固有のしくみを調べることができる.さらに,がく片は最外の器官であるため,成長の過程をイメージングすることが可能である.変異体の遺伝学的なスクリーニングにより,がく片の大きさおよび花弁のほかの器官の大きさが多様に変化する変異体をみつけ,花芽が異常な開き方をすることを見い出した.それらの変異体のうち,ftsh4変異体について詳細に調べた.
環境の変化が損傷や機能な損失を最小限にとどめて処理されるとき,そのシステムあるいは生物は頑健性をもつという.形態形成においては,擾乱に対し表現型が変化せず同じ形状を形成する性質を頑健性とよぶ.先行研究における遺伝学的なスクリーニングにおいては,すべての器官の大きさを制御することはできていたが,器官がどのように正確な大きさや形状をとるのかをみとおすことはできていなかった.そこで,器官の頑健性に直接に影響をおよぼすような変異体をスクリーニングした.器官の大きさを制御するしくみを知るためには,器官の大きさが平均値を中心にランダムに分布するような変異体において,何が制御因子であるかを同定する必要があるからである.シロイヌナズナにおいてAAA-ATPase活性およびメタロプロテアーゼ活性をもつ膜タンパク質性のプロテアーゼのひとつであるFtsH4の変異体においては,器官の大きさおよび形状が変化した.これまで,出芽酵母において頑健性に欠陥をもつ変異体が報告されていたが5,6),多細胞性の真核生物についての報告はなかった.
2.一様な器官の形成には細胞の多様性が必要とされる
なぜftsh4変異体において器官の大きさの頑健性が異常をきたすのかを理解するため,数理モデルを構築した.先行研究より7,8),成長の局所における制御と器官の形状をつなぐ成長をモデルとした.がく片の先端部から基部にかけて成長が遅くなるという観察にもとづき,先端部からある閾値までの領域において成長を遅くする仮想的なシグナルをくわえた.シグナルは下方に伝達され最基部に到達すると停止するとした.このいわゆる“成長前線”モデルにおいて,シミュレーションによるがく片の大きさは実際と同様に一定になった.
しかしながら,この成長前線モデルは成長したがく片において観察される細胞の多様性を考慮に入れていなかった.そこで,最初の疑問である細胞の多様性からどのようにして器官が制御されるのかという問題にたちもどり,がく片を“風船”にみたて,がく片の場所に依存した異なる弾性率をランダムにあたえた.いい換えると,弾性の異なる細胞をランダムに配置した.細胞を平衡に到達するまでふくらませ,がく片がどういう形状に到達するかを評価した.初期の時刻においてランダムに弾性率をあたえたモデルにおいては,いびつな“変異体もどき”の初期の弾性率の空間分布に応じた多様な形状をとった.しかしながら,成長の経過とともに時間的にランダムに弾性率をふりなおしたモデルにおいては,がく片は均一な形状をとった.このことから,成長の過程における細胞の多様性の時空間的な平均化が正常な器官を形成するうえで重要であることが示唆された.
3.野生型のがく片は空間的によりランダムに成長する
野生型のがく片において細胞の成長を観察したところ,12時間の間隔でみたときには時間的にも空間的にも多様であったが,細胞およびその娘細胞の成長の方向を48時間の間隔で累積したところほぼそろい,がく片の成長の方向と一致した(図1a).このことから,野生型のがく片は成長の方向を時空間的に平均化するしくみをもつことが示された.一方,ftsh4変異体のがく片の細胞は平均化の過程に欠陥が生じていた.ftsh4変異体の成長の方向は野生型よりも小さな時空間的な変動を示したが,48時間の間隔で累積したところ非常に多様なままであり,がく片の成長の方向とは一致しなかった(図1b).数理モデルおよび細胞壁の弾性の測定から,細胞を弾性体と考え,硬い細胞は成長が遅く柔らかい細胞は成長が速いと想定すると,野生型においては細胞の成長の多様性が高く,ftsh4変異体においては細胞の成長の多様性が低い可能性が示唆された.そこで,隣接した細胞の成長の空間的な多様性を定量化する手法を開発したところ,実際に,野生型のほうがftsh4変異体よりも細胞の成長の空間的な多様性の高いことが明らかにされた.また,野生型とftsh4変異体の器官の形状を比較するため形状の多様性を定量的に解析したところ,ftsh4変異体は野生型と比べがく片の大きさおよび形状の変動性がいちじるしく高かった.
以上の結果から,野生型のがく片においてはより多様な細胞の成長があり,その多様性が時空間的に平均化されることが明らかにされた.時空間的な平均化の考え方は,ショウジョウバエの胚においてhunchback遺伝子の確率的な転写と転写産物の均一な分布を説明する際に提案されているが9),形態形成において議論されたことはない.このような時空間的な平均化によりノイズに対応するしくみは,ほかの多くの生物学的な過程において共通するものなのかもしれない.
4.ftsh4変異体において活性酸素種の増加により大きさの欠陥がひき起こされる
すでに,FtsH4変異体はミトコンドリアに障害を起こし,その結果として活性酸素種を増加させ葉の形態異常をひき起こすことが知られている10).そこで,がく片の成長の段階において野生型およびftsh4変異体における活性酸素種の分布を評価した.その結果,野生型においては,がく片の成長とともに活性酸素種の分布ががく片の先端部から基部へと移行することがわかった(図2).一方で,ftsh4変異体においてはそのような活性酸素種の分布の段階的な移行は顕著には認められず,活性酸素種は成長の段階の早期において高濃度に達し,がく片の全体に不均一に分布していた.また,ftsh4変異体において活性酸素種を減少させることによりがく片の大きさおよび形状は均一化され,一方,野生型において活性酸素種を減少させると通常より大きく形状の均一ながく片が形成された.
おわりに
この研究は,器官の発生は細胞動態の巧みな組織化の連鎖により起こるとされてきた従来の発生生物学の基本的な問題に挑戦し,生命の正常な機能のひとつとして無秩序から秩序を創出するしくみを提案した.これまで,観察の結果から除くべきと考えられてきた細胞のノイズやゆらぎを,今後は,むしろ積極的に調べていくことにより形態形成にひそむしくみが明らかにされると考えている.また,この考え方はほかの生物にも適用できることから,研究の進んでいるショウジョウバエの羽や大腸菌の形態への応用にくわえ,ヒトを含むより高度な多細胞生物の大きさや形状を決める基本原理となっていることが期待される.
文 献
- Vogel, G.: Mysteries of development. How do organs know when they have reached the right size? Science, 340, 1156-1157 (2013)[PubMed]
- Anastasiou, E., Kenz, S., Gerstung, M. et al.: Control of plant organ size by KLUH/CYP78A5-dependent intercellular signaling. Dev. Cell, 13, 843-56 (2007)[PubMed]
- Roeder, A. H., Chickarmane, V., Cunha, A. et al.: Variability in the control of cell division underlies sepal epidermal patterning in Arabidopsis thaliana. PLoS Biol., 8, e1000367 (2010)[PubMed]
- Tauriello, G., Meyer, H. M., Smith, R. S. et al.: Variability and constancy in cellular growth of Arabidopsis sepals. Plant Phys., 169, 2342-2358 (2015)[PubMed]
- Levy, S. F. & Siegal, M. L.: Network hubs buffer environmental variation in Saccharomyces cerevisiae. PLoS Biol., 6, e264 (2008)[PubMed]
- Bauer, C. R., Li, S. & Siegal, M. L.: Essential gene disruptions reveal complex relationships between phenotypic robustness, pleiotropy, and fitness. Mol. Syst. Biol., 11, 773 (2015)[PubMed]
- Coen, E., Rolland-Lagan, A. G., Matthews, M. et al.: The genetics of geometry. Proc. Natl. Acad. Sci. USA, 101, 4728-4735 (2004)[PubMed]
- Sauret-Gueto, S., Schiessl, K., Bangham, A. et al.: JAGGED controls Arabidopsis petal growth and shape by interacting with a divergent polarity field. PLoS Biol., 11, e1001550 (2013)[PubMed]
- Little, S. C., Tikhonov, M. & Gregor, T.: Precise developmental gene expression arises from globally stochastic transcriptional activity. Cell, 154, 789-800 (2013)[PubMed]
- Kato, Y., Miura, E., Ido, K. et al.: The variegated mutants lacking chloroplastic FtsHs are defective in D1 degradation and accumulate reactive oxygen species. Plant Phys., 151, 1790-1801 (2009)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2013年 早稲田大学理工学術院先進理工学研究科博士課程 修了,2014年 北海道大学電子科学研究所 博士研究員を経て,2015年より同 特任助教.
研究テーマ:植物の器官の形態形成にひそむ多様性および不均一性の定量化.
抱負:形態形成,地震,乱流などの複雑な自然現象にひそむ普遍的な法則を明らかにしたい.
Arezki Boudaoud
フランスLyon大学Professor.
Adrienne H. K. Roeder
米国Cornell大学Assistant Professor.
© 2016 津川 暁・Arezki Boudaoud・Adrienne H. K. Roeder Licensed under CC 表示 2.1 日本
