WinglessシグナルはユビキチンリガーゼGodzillaに依存的なトランスサイトーシスにより活性化される
山崎 泰男
(スウェーデンGothenburg大学Sahlgrenska Academy)
email:山崎泰男
DOI: 10.7875/first.author.2016.024
Godzilla-dependent transcytosis promotes Wingless signalling in Drosophila wing imaginal discs.
Yasuo Yamazaki, Lucy Palmer, Cyrille Alexandre, Satoshi Kakugawa, Karen Beckett, Isabelle Gaugue, Ruth H. Palmer, Jean-Paul Vincent
Nature Cell Biology, 18, 451-457 (2016)
Wntシグナルは個体の発生および組織の恒常性の制御に必須であり,ショウジョウバエから哺乳動物にいたるまで高度に保存されている.ショウジョウバエの主要なWntはWinglessとよばれ,もっともよく機能の解析されたWntのひとつである.Winglessは翅原基とよばれる上皮細胞において生合成され分泌される.筆者らは,翅原基におけるWinglessの輸送の過程をその生合成から分泌まで追跡した.その結果,Winglessは頂端膜の近傍において生合成されたのち,いちど頂端膜の外に提示されるものの,エンドサイトーシスによりふたたび細胞に取り込まれた.そののち,トランスサイトーシスによりシグナル受容体の局在する基底外側膜まで輸送され,そこで分泌された.この頂端-基底軸にそったトランスサイトーシスはエンドソームに局在するユビキチンリガーゼGodzillaにより制御されていた.Godzillaが存在しないとWinglessの頂端-基底軸にそったトランスサイトーシスは停止し,Winglessシグナルは活性化されなかった.
上皮組織は消化管,気管,肺胞などに存在するからだの内外の表面をおおうシート状の組織である.体外あるいは管腔に面する側の細胞膜を頂端膜,他方を基底外側膜とよぶ.上皮組織を構成する上皮細胞は,ひとつの細胞にもかかわらず,その頂端膜の側と基底外側膜の側とで分布する分子および機能が異なる.このような細胞の方向性を極性とよぶ.極性化した上皮細胞は密着結合により互いに強く結合しており,これによりリガンドなどの物質が細胞のあいだをすり抜けることをふせいでいる.すなわち,極性化した細胞の頂端膜と基底外側膜はシグナルの制御の見地から互いに“絶縁”されている1).筆者らは,タグスイッチング系を用いて,ショウジョウバエの翅原基におけるWinglessの細胞内輸送の過程を生合成から分泌まで追跡した.
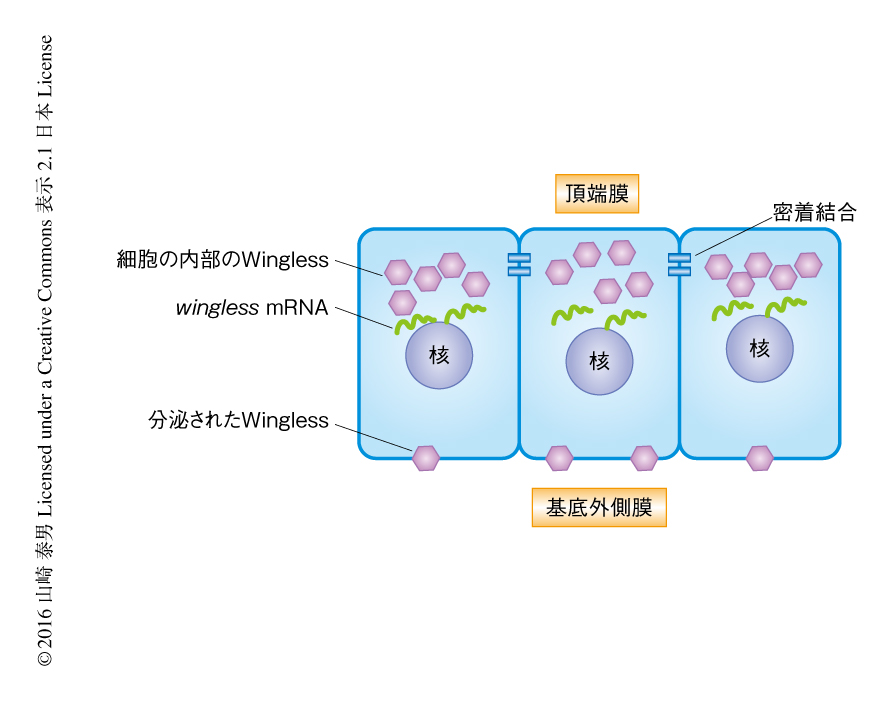
2種類の異なる抗体染色プロトコールによるWinglessの可視化により,翅原基においては異なる染色像が得られる2,3).界面活性剤により細胞の膜成分を透過処理したのちWinglessを染色すると,頂端膜の近傍に強い染色がみられた.Winglessの産生細胞においては多量のWinglessが生合成されていることから,この条件での染色はおもに細胞におけるWinglessの局在を反映すると考えられた(図1).実際に,in situハイブリダイゼーション法によりwingless mRNAを検出するとそのシグナルは頂端膜の近傍に観察されたことから,Winglessは頂端膜の近傍において生合成されていると考えられた(図1).一方,細胞の膜成分の透過処理をせずに細胞の外に分泌(あるいは,提示)されたWinglessを染色すると,頂端膜の側にはあまりWinglessは検出されず,基底外側膜の側に染色がみられるようになった(図1).以上の事実から,Winglessは頂端膜の近傍において生合成されたのち,基底外側膜の側へと輸送され,そこから分泌されるのではないかと考えた.
Winglessの細胞内輸送を追跡するため,内在性のwingless遺伝子座を異なる2種類のタグ化WinglessをコードするcDNAと置換したショウジョウバエ系統を作製した.この系統においては,一方のタグ化Winglessからもう一方のタグ化Winglessへと発現を任意の時期に切り替えることができる.この系においてはつねにいずれかのWinglessが内在性の遺伝子プロモーターの制御により発現しているので,組織の発生はまったく正常であり,Winglessの正常な細胞内輸送を追跡することができる.
タグを切り替えてから2時間のち,最初のWinglessのシグナルは頂端膜の近傍に検出された.このシグナルの多くはゴルジ体のマーカーと共局在したことから,予想どおり,Winglessは頂端膜の近傍において生合成されることが示された.タグを切り替えてから3時間のちには,シグナルとゴルジ体のマーカーとの共局在率は42%まで低下した.おそらくこれは,Winglessが分泌顆粒あるいはエンドソームへと移行したことによると考えられた.こののち,シグナルは基底外側膜の側へと移行しはじめ,タグを切り替えてから8時間のちには,シグナルはWinglessを産生する細胞の全体にみられるようになった.さらに,タグを切り替えてから16時間のちには,Winglessを産生していない近傍の細胞にもシグナルがみられるようになった.これは,Winglessが基底外側膜から分泌され,シグナル受容体に依存的して隣接した細胞に取り込まれたことを示すと考えられた.すなわち,Winglessは頂端膜の近傍において生合成されたのち,頂端-基底軸にそってトランスサイトーシスされ,基底外側膜から分泌されることが示された(図2).
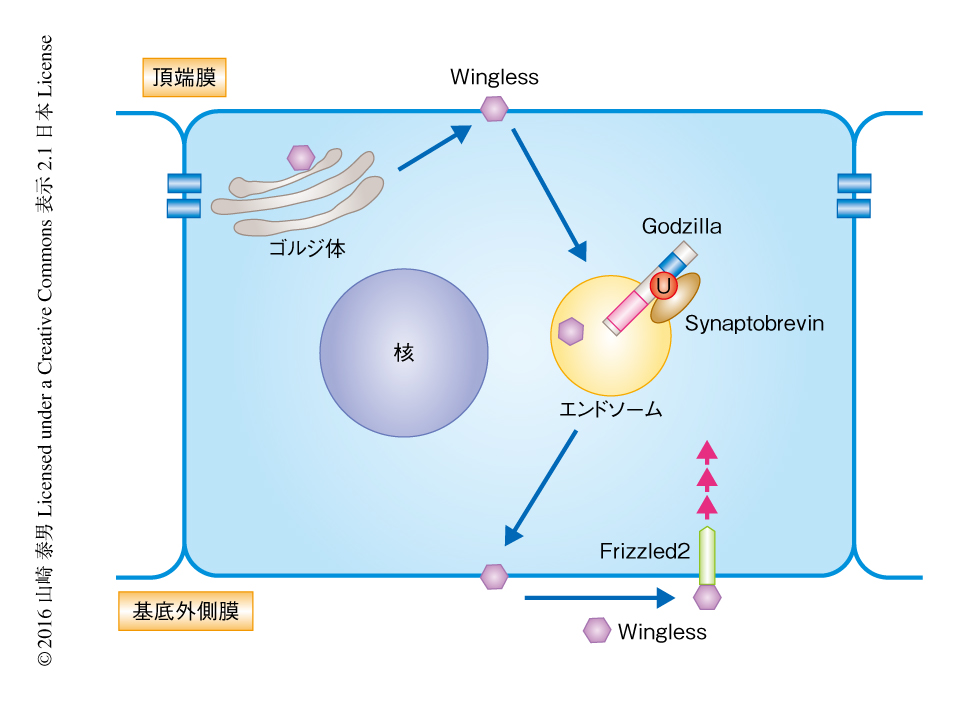
Winglessの頂端-基底軸にそったトランスサイトーシスはどのように制御されているのだろうか? Winglessは頂端膜を介したエンドサイトーシスをトランスサイトーシス経路へと移行する“乗り継ぎ点”として使うと仮定すると,エンドサイトーシスを阻害することによりWinglessの頂端-基底軸にそったトランスサイトーシスは観察されなくなるはずである.そこで,ショウジョウバエにおけるダイナミンのホモログであるShibireの温度依存性の変異体を用いて,Winglessのトランスサイトーシスに対するエンドサイトーシスの関与について調べた.使用した変異体は18℃では正常に機能するが34℃ではその機能を失う.細胞の外のWinglessを観察すると,18℃(すなわち,エンドサイトーシスが正常な状態)ではおもに基底外側膜にその染色がみられた.34℃にシフトすると(すなわち,エンドサイトーシスを停止すると)頂端膜に顕著な染色がみられるようになり,基底外側膜の染色は失われた.この輸送の障害は完全に可逆的であり,温度を18℃にもどすと頂端膜におけるWinglessの蓄積は消失し,ふたたび基底外側膜に染色が観察された.以上の結果から,Winglessの頂端-基底軸にそったトランスサイトーシスは頂端膜を介したエンドサイトーシスにより制御されることが明らかにされた(図2).
なにがWinglessのトランスサイトーシスを制御しているかについて明らかにするため,RNAi法を用いたスクリーニングをした.その結果,ユビキチンリガーゼGodzillaをノックダウンすると成虫の羽に“notching”とよばれるWinglessシグナルの阻害にともない生じる特徴的な表現型が生じた.GodzillaはPA-TM-RINGユビキチンリガーゼに分類されるエンドソームに局在する膜型のユビキチンリガーゼで,初期エンドソームからリサイクリングエンドソームへの発芽を制御することが報告されている4).詳細に調べると,Godzillaをノックダウンした細胞ではWinglessの標的であるSenselessの発現が阻害され,Winglessはその産生細胞に強く蓄積していた.Winglessの発現量に変化はなかったことから,GodzillaはWinglessの分泌を促進的に制御すると考えられた.同様のWinglessの蓄積は,Godzillaをノックアウトした細胞と野生型の細胞からなるモザイク組織においても確認された.以上の事実から,Godzillaはエンドソームからの発芽経路を介してWinglessの細胞内輸送(おそらくは,トランスサイトーシス)を制御するのではないかと考えた.
Godzillaによるエンドソームからの発芽制御は,エンドソームに存在するSNAREタンパク質Synaptobrevin(ヒトでは,VAMP3)のユビキチン化を介することが明らかにされている4).Synaptobrevinの機能を欠損した組織には,Godzillaの機能を欠損した組織と同様に,Winglessシグナルの不活性化および細胞におけるWinglessの蓄積が観察された.したがって,GodzillaによるWinglessの細胞内輸送の制御は,その活性に依存すると考えられた.Godzillaがエンドソームを介するWinglessの細胞内輸送を制御しているのであれば,WinglessはエンドソームにおいてGodzillaと共局在するはずである.恒常活性型のRab5を用いてエンドソームを巨大化しWinglessおよびGodzillaの局在を観察すると,両者はエンドソームにおいて明瞭に共局在していていた.これらの結果から,GodzillaとWinglessはエンドソームにおいて会合することにより,Winglessをトランスサイトーシス経路へと移行するのではないかと考えた.
GodzillaのWinglessのトランスサイトーシスへの関与について明らかにするため,Godzillaをノックダウンした細胞において細胞の外に分泌(および,提示)されたWinglessの局在を観察した.Godzillaをノックダウンした細胞において基底外側膜のWinglessは消失し,一方,頂端膜に強い蓄積がみられた.すなわち,エンドサイトーシスを阻害した場合と同様の表現型が観察された.さらに確認のため,ユビキチンリガーゼ活性を欠失したGodzilla変異体を作製し,トランスサイトーシスおよびWinglessシグナルに対する影響について調べた.Godzilla変異体を翅原基に異所性に発現させるとSenselessの発現は阻害され,基底外側膜のWinglessも消失した.以上の結果から,GodzillaはエンドソームにおいてWinglessの頂端-基底軸にそったトランスサイトーシスを制御していることが明らかにされた(図2).
なぜWinglessはトランスサイトーシスされるのだろうか? おそらく,基底外側膜にシグナル受容体が存在するからではないかと考えた.翅原基の頂端膜の側は上皮細胞によりおおわれているため,血リンパは基底外側膜の側のみからしか接触しない.そこで,血リンパにNotumを異所性に発現させWinglessシグナルに対する影響について観察した.Notumは細胞の外に分泌されたWntを脱アシル化することによりWingless/Wntシグナルを阻害する5).その結果,Winglessの標的であるSenselessの発現は血リンパに由来するNotumにより阻害された.すなわち,Winglessシグナルは基底外側膜から入力されることが示唆された.さらに直接的な方法として,Winglessの内因性のシグナル受容体Frizzled2の翅原基における局在を観察したところ,Frizzled2はおもに基底外側膜の側に局在していた.これらの結果から,Winglessによる,少なくともSenselessの発現を十分に誘導する量のシグナルは,基底外側膜から入力されると結論した.
ショウジョウバエの胚上皮組織において内因性のWinglessは頂端膜から分泌されることが示唆されていたことから6,7),翅原基においてもWinglessは頂端膜から分泌されると考えられていた.今回の研究により,翅原基においてWinglessはむしろ基底外側膜の側から分泌され,そのシグナルを入力していることが明らかにされた.この機構が,ショウジョウバエのほかの上皮組織,さらには,哺乳動物においても普遍的であるのかどうかについては,今後,検討されるべき課題であるが,3次元培養された哺乳動物に由来する培養細胞において,Wnt3aがトランスサイトーシスされることが報告されていることは興味深い8).しかし,依然として疑問は残る.なぜWinglessはトランスサイトーシスされる必要があるのだろうか? Wingless/Wntがエキソソームに含まれることが報告されていることを考えると9),トランスサイトーシスはエキソソームへのパッキングのためなのかもしれない.また,Wntの活性化には細胞において低いpH(たとえば,エンドソーム)に曝露される必要のあることを示唆する報告もある10).今後,いずれの可能性についても検討すべきであろう.
これまで,エンドソームに含まれるWingless/Wntは,シグナル受容体に結合したのち受容体に依存性のエンドサイトーシスにより細胞に取り込まれたもの,すなわち,“シグナル中あるいはシグナル後”のリガンドであると考えられてきた.今回の研究により,エンドソームには“シグナル前”のリガンド,すなわち,分泌されるまえのWingless/Wntも含まれることが明らかにされ,研究結果の解釈に新たな可能性をくわえなければならないと考えられる.
略歴:2003年 明治薬科大学大学院薬学研究科博士課程 修了,2004年 明治薬科大学薬学部 助手,2009年 スウェーデンUmea大学 博士研究員を経て,2014年よりスウェーデンGothenburg大学 博士研究員.
研究テーマ:細胞内輸送,とくに,極性的な輸送.
関心事:分子レベルの事象が,細胞,組織,個体にどのように伝達されるのかに興味がある.
© 2016 山崎 泰男 Licensed under CC 表示 2.1 日本
(スウェーデンGothenburg大学Sahlgrenska Academy)
email:山崎泰男
DOI: 10.7875/first.author.2016.024
Godzilla-dependent transcytosis promotes Wingless signalling in Drosophila wing imaginal discs.
Yasuo Yamazaki, Lucy Palmer, Cyrille Alexandre, Satoshi Kakugawa, Karen Beckett, Isabelle Gaugue, Ruth H. Palmer, Jean-Paul Vincent
Nature Cell Biology, 18, 451-457 (2016)
要 約
Wntシグナルは個体の発生および組織の恒常性の制御に必須であり,ショウジョウバエから哺乳動物にいたるまで高度に保存されている.ショウジョウバエの主要なWntはWinglessとよばれ,もっともよく機能の解析されたWntのひとつである.Winglessは翅原基とよばれる上皮細胞において生合成され分泌される.筆者らは,翅原基におけるWinglessの輸送の過程をその生合成から分泌まで追跡した.その結果,Winglessは頂端膜の近傍において生合成されたのち,いちど頂端膜の外に提示されるものの,エンドサイトーシスによりふたたび細胞に取り込まれた.そののち,トランスサイトーシスによりシグナル受容体の局在する基底外側膜まで輸送され,そこで分泌された.この頂端-基底軸にそったトランスサイトーシスはエンドソームに局在するユビキチンリガーゼGodzillaにより制御されていた.Godzillaが存在しないとWinglessの頂端-基底軸にそったトランスサイトーシスは停止し,Winglessシグナルは活性化されなかった.
はじめに
上皮組織は消化管,気管,肺胞などに存在するからだの内外の表面をおおうシート状の組織である.体外あるいは管腔に面する側の細胞膜を頂端膜,他方を基底外側膜とよぶ.上皮組織を構成する上皮細胞は,ひとつの細胞にもかかわらず,その頂端膜の側と基底外側膜の側とで分布する分子および機能が異なる.このような細胞の方向性を極性とよぶ.極性化した上皮細胞は密着結合により互いに強く結合しており,これによりリガンドなどの物質が細胞のあいだをすり抜けることをふせいでいる.すなわち,極性化した細胞の頂端膜と基底外側膜はシグナルの制御の見地から互いに“絶縁”されている1).筆者らは,タグスイッチング系を用いて,ショウジョウバエの翅原基におけるWinglessの細胞内輸送の過程を生合成から分泌まで追跡した.
1.翅原基におけるWinglessの局在
2種類の異なる抗体染色プロトコールによるWinglessの可視化により,翅原基においては異なる染色像が得られる2,3).界面活性剤により細胞の膜成分を透過処理したのちWinglessを染色すると,頂端膜の近傍に強い染色がみられた.Winglessの産生細胞においては多量のWinglessが生合成されていることから,この条件での染色はおもに細胞におけるWinglessの局在を反映すると考えられた(図1).実際に,in situハイブリダイゼーション法によりwingless mRNAを検出するとそのシグナルは頂端膜の近傍に観察されたことから,Winglessは頂端膜の近傍において生合成されていると考えられた(図1).一方,細胞の膜成分の透過処理をせずに細胞の外に分泌(あるいは,提示)されたWinglessを染色すると,頂端膜の側にはあまりWinglessは検出されず,基底外側膜の側に染色がみられるようになった(図1).以上の事実から,Winglessは頂端膜の近傍において生合成されたのち,基底外側膜の側へと輸送され,そこから分泌されるのではないかと考えた.
2.タグスイッチング系によるWinglessの追跡
Winglessの細胞内輸送を追跡するため,内在性のwingless遺伝子座を異なる2種類のタグ化WinglessをコードするcDNAと置換したショウジョウバエ系統を作製した.この系統においては,一方のタグ化Winglessからもう一方のタグ化Winglessへと発現を任意の時期に切り替えることができる.この系においてはつねにいずれかのWinglessが内在性の遺伝子プロモーターの制御により発現しているので,組織の発生はまったく正常であり,Winglessの正常な細胞内輸送を追跡することができる.
タグを切り替えてから2時間のち,最初のWinglessのシグナルは頂端膜の近傍に検出された.このシグナルの多くはゴルジ体のマーカーと共局在したことから,予想どおり,Winglessは頂端膜の近傍において生合成されることが示された.タグを切り替えてから3時間のちには,シグナルとゴルジ体のマーカーとの共局在率は42%まで低下した.おそらくこれは,Winglessが分泌顆粒あるいはエンドソームへと移行したことによると考えられた.こののち,シグナルは基底外側膜の側へと移行しはじめ,タグを切り替えてから8時間のちには,シグナルはWinglessを産生する細胞の全体にみられるようになった.さらに,タグを切り替えてから16時間のちには,Winglessを産生していない近傍の細胞にもシグナルがみられるようになった.これは,Winglessが基底外側膜から分泌され,シグナル受容体に依存的して隣接した細胞に取り込まれたことを示すと考えられた.すなわち,Winglessは頂端膜の近傍において生合成されたのち,頂端-基底軸にそってトランスサイトーシスされ,基底外側膜から分泌されることが示された(図2).
3.Winglessの頂端-基底軸にそったトランスサイトーシスはエンドサイトーシスにより制御される
Winglessの頂端-基底軸にそったトランスサイトーシスはどのように制御されているのだろうか? Winglessは頂端膜を介したエンドサイトーシスをトランスサイトーシス経路へと移行する“乗り継ぎ点”として使うと仮定すると,エンドサイトーシスを阻害することによりWinglessの頂端-基底軸にそったトランスサイトーシスは観察されなくなるはずである.そこで,ショウジョウバエにおけるダイナミンのホモログであるShibireの温度依存性の変異体を用いて,Winglessのトランスサイトーシスに対するエンドサイトーシスの関与について調べた.使用した変異体は18℃では正常に機能するが34℃ではその機能を失う.細胞の外のWinglessを観察すると,18℃(すなわち,エンドサイトーシスが正常な状態)ではおもに基底外側膜にその染色がみられた.34℃にシフトすると(すなわち,エンドサイトーシスを停止すると)頂端膜に顕著な染色がみられるようになり,基底外側膜の染色は失われた.この輸送の障害は完全に可逆的であり,温度を18℃にもどすと頂端膜におけるWinglessの蓄積は消失し,ふたたび基底外側膜に染色が観察された.以上の結果から,Winglessの頂端-基底軸にそったトランスサイトーシスは頂端膜を介したエンドサイトーシスにより制御されることが明らかにされた(図2).
4.WinglessはユビキチンリガーゼGodzillaに依存的にトランスサイトーシスされる
なにがWinglessのトランスサイトーシスを制御しているかについて明らかにするため,RNAi法を用いたスクリーニングをした.その結果,ユビキチンリガーゼGodzillaをノックダウンすると成虫の羽に“notching”とよばれるWinglessシグナルの阻害にともない生じる特徴的な表現型が生じた.GodzillaはPA-TM-RINGユビキチンリガーゼに分類されるエンドソームに局在する膜型のユビキチンリガーゼで,初期エンドソームからリサイクリングエンドソームへの発芽を制御することが報告されている4).詳細に調べると,Godzillaをノックダウンした細胞ではWinglessの標的であるSenselessの発現が阻害され,Winglessはその産生細胞に強く蓄積していた.Winglessの発現量に変化はなかったことから,GodzillaはWinglessの分泌を促進的に制御すると考えられた.同様のWinglessの蓄積は,Godzillaをノックアウトした細胞と野生型の細胞からなるモザイク組織においても確認された.以上の事実から,Godzillaはエンドソームからの発芽経路を介してWinglessの細胞内輸送(おそらくは,トランスサイトーシス)を制御するのではないかと考えた.
Godzillaによるエンドソームからの発芽制御は,エンドソームに存在するSNAREタンパク質Synaptobrevin(ヒトでは,VAMP3)のユビキチン化を介することが明らかにされている4).Synaptobrevinの機能を欠損した組織には,Godzillaの機能を欠損した組織と同様に,Winglessシグナルの不活性化および細胞におけるWinglessの蓄積が観察された.したがって,GodzillaによるWinglessの細胞内輸送の制御は,その活性に依存すると考えられた.Godzillaがエンドソームを介するWinglessの細胞内輸送を制御しているのであれば,WinglessはエンドソームにおいてGodzillaと共局在するはずである.恒常活性型のRab5を用いてエンドソームを巨大化しWinglessおよびGodzillaの局在を観察すると,両者はエンドソームにおいて明瞭に共局在していていた.これらの結果から,GodzillaとWinglessはエンドソームにおいて会合することにより,Winglessをトランスサイトーシス経路へと移行するのではないかと考えた.
GodzillaのWinglessのトランスサイトーシスへの関与について明らかにするため,Godzillaをノックダウンした細胞において細胞の外に分泌(および,提示)されたWinglessの局在を観察した.Godzillaをノックダウンした細胞において基底外側膜のWinglessは消失し,一方,頂端膜に強い蓄積がみられた.すなわち,エンドサイトーシスを阻害した場合と同様の表現型が観察された.さらに確認のため,ユビキチンリガーゼ活性を欠失したGodzilla変異体を作製し,トランスサイトーシスおよびWinglessシグナルに対する影響について調べた.Godzilla変異体を翅原基に異所性に発現させるとSenselessの発現は阻害され,基底外側膜のWinglessも消失した.以上の結果から,GodzillaはエンドソームにおいてWinglessの頂端-基底軸にそったトランスサイトーシスを制御していることが明らかにされた(図2).
5.Winglessのシグナル受容体は翅原基の基底外側膜に局在する
なぜWinglessはトランスサイトーシスされるのだろうか? おそらく,基底外側膜にシグナル受容体が存在するからではないかと考えた.翅原基の頂端膜の側は上皮細胞によりおおわれているため,血リンパは基底外側膜の側のみからしか接触しない.そこで,血リンパにNotumを異所性に発現させWinglessシグナルに対する影響について観察した.Notumは細胞の外に分泌されたWntを脱アシル化することによりWingless/Wntシグナルを阻害する5).その結果,Winglessの標的であるSenselessの発現は血リンパに由来するNotumにより阻害された.すなわち,Winglessシグナルは基底外側膜から入力されることが示唆された.さらに直接的な方法として,Winglessの内因性のシグナル受容体Frizzled2の翅原基における局在を観察したところ,Frizzled2はおもに基底外側膜の側に局在していた.これらの結果から,Winglessによる,少なくともSenselessの発現を十分に誘導する量のシグナルは,基底外側膜から入力されると結論した.
おわりに
ショウジョウバエの胚上皮組織において内因性のWinglessは頂端膜から分泌されることが示唆されていたことから6,7),翅原基においてもWinglessは頂端膜から分泌されると考えられていた.今回の研究により,翅原基においてWinglessはむしろ基底外側膜の側から分泌され,そのシグナルを入力していることが明らかにされた.この機構が,ショウジョウバエのほかの上皮組織,さらには,哺乳動物においても普遍的であるのかどうかについては,今後,検討されるべき課題であるが,3次元培養された哺乳動物に由来する培養細胞において,Wnt3aがトランスサイトーシスされることが報告されていることは興味深い8).しかし,依然として疑問は残る.なぜWinglessはトランスサイトーシスされる必要があるのだろうか? Wingless/Wntがエキソソームに含まれることが報告されていることを考えると9),トランスサイトーシスはエキソソームへのパッキングのためなのかもしれない.また,Wntの活性化には細胞において低いpH(たとえば,エンドソーム)に曝露される必要のあることを示唆する報告もある10).今後,いずれの可能性についても検討すべきであろう.
これまで,エンドソームに含まれるWingless/Wntは,シグナル受容体に結合したのち受容体に依存性のエンドサイトーシスにより細胞に取り込まれたもの,すなわち,“シグナル中あるいはシグナル後”のリガンドであると考えられてきた.今回の研究により,エンドソームには“シグナル前”のリガンド,すなわち,分泌されるまえのWingless/Wntも含まれることが明らかにされ,研究結果の解釈に新たな可能性をくわえなければならないと考えられる.
文 献
- Tuma, P. & Hubbard, A. L.: Transcytosis: crossing cellular barriers. Physiol. Rev., 83, 871-932 (2003)[PubMed]
- Strigini, M. & Cohen, S. M.: Wingless gradient formation in the Drosophila wing. Curr. Biol., 10, 293-300 (2000)[PubMed]
- Wu, J., Klein, T. J. & Mlodzik, M.: Subcellular localization of frizzled receptors, mediated by their cytoplasmic tails, regulates signaling pathway specificity. PLoS Biol., 2, E158 (2004)[PubMed]
- Yamazaki, Y., Schonherr, C., Varshney, G. K. et al.: Goliath family E3 ligases regulate the recycling endosome pathway via VAMP3 ubiquitylation. EMBO J., 32, 524-537 (2013)[PubMed]
- Kakugawa, S., Langton, P. F., Zebisch, M. et al.: Notum deacylates Wnt proteins to suppress signalling activity. Nature, 519, 187-192 (2015)[PubMed]
- Simmonds, A. J., dosSantos, G., Livne-Bar, I. et al.: Apical localization of wingless transcripts is required for wingless signaling. Cell, 105, 197-207 (2001)[PubMed]
- Wilkie, G. S. & Davis, I.: Drosophila wingless and pair-rule transcripts localize apically by dynein-mediated transport of RNA particles. Cell, 105, 209-219 (2001)[PubMed]
- Yamamoto, H., Awada, C., Hanaki, H. et al.: The apical and basolateral secretion of Wnt11 and Wnt3a in polarized epithelial cells is regulated by different mechanisms. J. Cell Sci., 126, 2931-2943 (2013)[PubMed]
- Gross, J. C., Chaudhary, V., Bartscherer, K. et al.: Active Wnt proteins are secreted on exosomes. Nat. Cell Biol., 14, 1036-1045 (2012)[PubMed]
- Coombs, G. S., Yu, J., Canning, C. A. et al.: WLS-dependent secretion of WNT3A requires Ser209 acylation and vacuolar acidification. J. Cell Sci., 123, 3357-3367 (2010)[PubMed]
活用したデータベースにかかわるキーワードと統合TVへのリンク
著者プロフィール
略歴:2003年 明治薬科大学大学院薬学研究科博士課程 修了,2004年 明治薬科大学薬学部 助手,2009年 スウェーデンUmea大学 博士研究員を経て,2014年よりスウェーデンGothenburg大学 博士研究員.
研究テーマ:細胞内輸送,とくに,極性的な輸送.
関心事:分子レベルの事象が,細胞,組織,個体にどのように伝達されるのかに興味がある.
© 2016 山崎 泰男 Licensed under CC 表示 2.1 日本
